
Abstract
To develop a therapeutic vaccine against hepatitis B virus surface antigen (HBsAg)-expressing liver cancer, we tried to prepare a vaccine by formulating recombinant HBsAg with BW006, a B type CpG oligodeoxynucleotide (ODN) with Th1-biasing activity, and examined its potency of inducing therapeutic immunity against HBsAg-expressing liver cancer cells in mice. When applied therapeutically, BW006 could assist HBsAg to induce vigorous immune responses capable of inhibiting the growth of HBsAg-expressing liver cancer cells and prolonging the survival of mice bearing HBsAg-expressing liver cancer cells. In vivo and in vitro experiments showed that the BW006-adjuvanted HBsAg enhanced the production of IgG2a antibodies, interferon-γ, and interleukin-12 and facilitated the generation of specific cytotoxic T lymphocyte that killed the HBsAg-expressing liver cancer cells. These results suggest that the BW006-adjuvanted HBsAg might be developed into a candidate tumor vaccine for the treatment of HBsAg-expressing liver cancer.
Introduction
It is well established that hepatocellular carcinoma (HCC) is associated with the persistent infection of hepatitis B virus (HBV). 1 Approximately, one-third of HCC patients are identified with HBsAg expressed on their hepatocytes and HCC cells. The patients with chronic HBV infection has been recognized as a high risk factor for the development of HCC, 2 implying that HBsAg could serve as a tumor-associated antigen (TAA) in HBsAg-expressing liver cancer. Presumably, HBsAg, included in commercialized vaccines for preventing HBV infection, could be formulated into a vaccine for the treatment of HBsAg-expressing liver cancer.
Currently, in commercialized vaccines for HBV infection prevention, recombinant HBsAg produced in yeast or mammalian cells 3 is formulated in aluminum salt (Alum). Although safe and well tolerated, the alum is a relatively weak adjuvant for the vaccines. About 10% of those people who received three doses of the vaccine failed to produce a protective level of anti-HBsAg antibody (HBsAb). 4 Moreover, the Alum-adjuvanted HBsAg is unable to induce Th1-biased immune responses with elevation of interferon-γ (IFN-γ), tumor necrosis factor (TNF)-α, interleukin (IL)-12, and IgG2a antibodies and instead induce a Th2-biased immune response characterized by enhancing production of IL-4, IL-10, and IgG1 antibodies. 5,6 As reported, Th1-biased cytokines are significantly correlated with the clearance of tumor cells by providing a favorable milieu for a TAA to induce specific cytotoxic T lymphocytes (CTLs). 7,8 Ideally, the adjuvant formulated with HBsAg should be capable of promoting a Th1-biased immune response with a vigorous generation of specific CTLs against HBsAg-expressing liver cancer cells. Apparently, it is required to have a potent Th1-biasing adjuvant to develop therapeutic vaccines for the treatment of HBsAg-expressing liver cancer cells.
In recent years, multiple studies have shown that the synthetic unmethylated CpG-containing oligodeoxynucleotide (CpG ODN) could be used as a Th1-biasing adjuvant. 9,10 After recognizing toll-like receptor 9 (TLR9), CpG ODN acts as a immune stimulator to induce Th1-biasing cytokines in favor of the generation of specific CTLs. 11,12 The properties make the CpG ODNs promising adjuvants for tumor vaccines. 13 –17 In mouse models, coadministration of tumor vaccine with CpG ODN was capable of inducing rejection of glioblastoma, 14 B16 melanoma, 18 prostate cancer, 19 and gastric cancer. 13 In clinical trials, CpG ODN-adjuvanted cancer vaccines showed therapeutic potential in patients with melanoma, breast cancer, sarcoma, ovarian cancer, and glioblastoma. 20 –22 Although there are no approving researches on CpG ODN-adjuvanted HBsAg as therapeutic cancer vaccines against HBsAg-expressing liver cancer, the CpG ODN has been shown to be a promising adjuvant for developing vaccines capable of inducing HBsAg-specific cellular immune responses in mice, baboons, and humans. 23 –28 When used in combination with Alum-adjuvanted HBsAg, CpG ODN facilitated the vaccine to induce a Th1-biased immune response to HBsAg. These data imply that CpG ODN could be of value in assisting HBsAg to induce immune responses against HBsAg-expressing liver cancer.
In our previous study, BW006, 29 a B-type CpG ODN, was found to be potent in stimulating the proliferation of human peripheral blood mononuclear cells (PBMCs) and mouse spleen cells. In current study, we investigated whether BW006 could assist HBsAg to induce therapeutic immunity against HBsAg-expressing liver cancer in mice. We found that therapeutic application of BW006-adjuvanted HBsAg induced Th1-biased immune responses that inhibited the growth of HBsAg-expressing liver cancer cells and prolonged the survival of mice bearing HBsAg-expressing liver cancer cells.
Materials and Methods
Oligodeoxynucleotides
Purified single-stranded ODNs were synthesized in Takara Company. The following CpG ODNs were used: 2216 (5′-GGgggACGATCGTCgggggG-3′), 1018 (5′-TGACTGTGAACGTTCGAGATGA-3′), 1585 (5′-ggGGTCAACGTTGAGggggg-3′), 1826 (5′-tc catgacgttcctgacgtt-3′), C274 (5′-tcgtcgaacgttcgagatgat-3′), BW006 (5′-TCGACGTTCGTCGTTCGTC GTTC-3′), and BW007 (5′-TGCAGCTTGCTGCTTGCTGCTTC-3′), the same as BW006 except that the CG sequence is reversed, was used as a control for BW006. Lowercase and capital letters represent phosphorothioate and phosphodiester linkage, respectively. All CpG ODNs were diluted in phosphate-buffered saline (PBS) and had no detectable endotoxin (Limulus amebocyte lysate assay; Associates of Cape Cod, Inc.). All reagents used were pyrogen-free.
Construction of the HBsAg expression vector
The gene encoding HBsAg was amplified from HBV genomic DNA by polymerase chain reaction (PCR) using forward (5′-AAGCTTGCCACCATGGAGAACATCACATC-3′) and reverse (5′-CCCCGAATTCTCAAATGTA TACCCAAAGAC-3′) primers containing EcoRI and HindIII sites, respectively. The PCR product (703 bp) was cloned into a pcDNA3 plasmid vector (Novagen) to yield pcDNA3-HBsAg. The insert in the pcDNA3-HBsAg was verified by DNA sequencing.
Cell and cell lines
Human PBMCs from freshly drawn healthy volunteer blood (The Blood Center of Jilin Province) were isolated by Ficoll–Hypaque density gradient centrifugation (Pharmacia). Splenocytes were isolated from euthanized BALB/c mice by lyzing erythrocytes with lysis buffer (10 mM KHCO3, 150 mM NH4Cl, and 10 mM EDTA, pH 7.4). Murine H22 hepatoma cells of BALB/c origin were transfected with the pcDNA3-HBsAg plasmid encoding the human HBsAg, resulting in HBsAg-expressing H22 cell line cells, designated HBsAg+ H22 cells. The HBsAg+ H22 cells were cultured in the selection medium with 800 μg/mL G418 (Sigma-Aldrich) and verified by reverse transcription polymerase chain reaction (RT-PCR) and Western blot analysis.
Mice
Female BALB/c mice (6–8 weeks old) were purchased from Animal Center of Norman Bethune Medical College, Jilin University; maintained in laminar flow rooms; and used for experiments in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals, and with the approval of the Scientific Investigation Board of Science and Technology of Jilin Province, China.
Proliferation assays
Proliferation of human PBMCs or mouse splenocytes was determined by [3H] thymidine incorporation assay. Briefly, human PBMCs or mouse splenocytes (6×105/well) were plated in 96-well U-bottomed plates (Costar) and cultured with or without CpG ODN (6 μg/mL) for 36 hours, followed by pulsing with [3H] thymidine (New England Nuclear) for 16 hours. The cells were harvested on glass fiber filters and detected in a scintillation counter.
Flow cytometry
PBMCs or mouse splenocytes were incubated with or without CpG ODN (6 μg/mL) for 12 hours, washed twice with FACS staining buffer (PBS supplemented with 0.5% bovine serum albumin, 0.01% NaN3, and 100 mM EDTA), and then analyzed for the expression of various cell surface molecules by dual-color staining with FITC-conjugated anti-CD69 mAb (BD Biosciences) and PE-conjugated anti-CD19 (BD Biosciences) followed by analysis on a FACS calibur.
Determination of cytokines
Female BALB/c mice (6–8 weeks old; n=5) were immunized subcutaneously (s.c.) at inguinal lymph node areas in both sides with HBsAg alone or HBsAg absorbed in Alum (HBsAg+Alum) or HBsAg formulated with BW006 (HBsAg+BW006) or BW006 plus HBsAg+Alum (HBsAg+Alum+BW006) and PBS, respectively, for four times at a 7-day interval. On day 7 after the last immunization, splenocytes were isolated and stimulated in vitro with HBsAg (50 μg/mL) for 24 hours and the supernatants were harvested to measure IFN-γ and IL-12 using ELISA kits (R&D System), according to the manufacturer's protocols.
Determination of HBsAg-specific antibodies
Mice were immunized as described above and bled on day 7 after the last immunization. The sera were isolated and HBsAg-specific IgG, IgG2a, or IgG1 were detected by ELISA. Briefly, all wells in 96-well plates were coated with HBsAg (1 μg/well) in PBS (pH 7.3) and incubated overnight at 4°C and blocked using 300 μL of 5% (v/v) fetal bovine serum in PBS at 37°C for 1 hour after washing. The test and control sera were subjected to a serial of 100-fold dilutions with PBS-Tween 20 (0.05% Tween 20, v/v; PBST) and the plates were incubated at 37°C for 1 hour. After washing three times with PBST, 100 μL/well of appropriately diluted HRP-conjugated anti-mouse IgG1 or IgG2a (Santa Cruz Biotechnology, Inc.) were added to the plates and incubated at 37°C for 1 hour. After removal of unbound conjugates, positive binding was detected by adding substrate (OPD/H2O2) and the plates were incubated for 15 minutes at 37°C in dark. The reaction was stopped by sulfuric acid and the absorbance was read at 492 nm.
Tumor inoculation and immunization
Mice (n=8) were inoculated with HBsAg+ H22 cells (2×106/mouse) in 0.2 mL serum-free medium s.c. at the right back near hind leg on day −1 and then s.c. injected with PBS or HBsAg (1 μg/mouse) or HBsAg+Alum (HBsAg 1 μg/mouse in aluminum hydroxide) or HBsAg+BW006 (HBsAg 1 μg/mouse plus BW006 12.5 μg/mouse) or HBsAg+Alum+BW006 (HBsAg 1 μg/mouse and BW006 12.5 μg/mouse in aluminum hydroxide) at inguinal lymph node areas in both sides on days 0, 7, 14, 21, respectively. The mice were monitored for measuring tumor volumes (length×width2×0.5, in cubic millimeters) every other day and recoding the survivals for 100 days post-tumor inoculation.
Specific CTL killing assay
Mice were immunized as described above. On day 7 after the last immunization, splenocytes were isolated and cultured with HBsAg for 2 days in vitro. Mouse IL-2 (20 U/mL) was added to the culture 24 hours later. The splenocytes were centrifuged, counted, and used as effector cells. HBsAg+ H22 cells were used as target cells. Briefly, 5×105 splenocytes were serially diluted and cultured with HBsAg+ H22 cells per well in a 96-well plate at effector/target ratios of 12.5:1, 25:1, and 50:1 at 37°C. After 4 hours, the cells were collected, stained with trypan blue, and counted. The amount of the remaining live HBsAg+ H22 cells was calculated using the following formula: cell number/mL=total cell number in four big squares/4×dilute multiple×104. The cell number in four big squares must be between 200 and 500, and concentration of cells must be greater than 104/mL. The cytotoxicity of the spleen cells was calculated based on the formula: [1−experiment cell number/cell number in medium control] ×100%.
Statistical analysis
Tumor growth curves were plotted based on tumor size measured when the first tumor transplanted mouse died. Differences in tumor size among the various groups were determined by the analysis of variance repeated-measures test. Statistical significance for survival of mice was assessed using the Kaplan–Meier analysis. A p-value of <0.05 was considered statistically significant. Statistical analyses were performed using SPSS software.
Results
Immunostimulatory properties of BW006
To find whether CpG ODNs could be used as an adjuvant for HBsAg to induce immune response against HBsAg-expressing liver cancer cells, we have synthesized a series of CpG ODNs and tested their immunostimulatory capacities to stimulate the proliferation of human PBMCs and the activation of immune cells in the PBMCs. In the process, a CpG ODN, designated as BW006, stood out. In human PBMC proliferation assay, BW006 could stimulate human PBMCs to proliferate vigorously. The efficacy was stronger than that induced by 2216 (p=0.001), a prototype of type-A CpG ODN for human, and similar to those induced by 1018, a prototype of type-B CpG ODN for human, and C274 (p>0.05 vs.BW006), a prototype of type-C CpG ODN (Fig. 1A). Meanwhile, BW007, a GC-reversed control for BW006, failed to stimulate the proliferation of the human PBMCs (p>0.05 vs. medium). To determine the cells activated by BW006, the human PBMCs cultured with BW006 were analyzed for their upregulation of CD69 molecule. The flow cytometry analysis showed that BW006 upregulated expression of CD69 molecule in CD19+, CD56+, or CD8+ cells (Fig. 1B), indicating that BW006 could activate human B cells, NK cells, and T cells. Furthermore, the human PBMCs activated by BW006 were analyzed for their upregulation of human leukocyte antigen class I (HLA-A2) molecules. The result demonstrated that BW006 could upregulate HLA-A2 in human PBMC (Fig.1C) in a similar capacity as 1018. Taken together, BW006 could stimulate the proliferation of human PBMCs; activate human B cells, NK cells, and T cells; and upregulate MHC I molecules, showing the potential as a promising adjuvant to assist HBsAg to induce immunity against HBsAg-expressing liver cancer cells.

Immunostimulatory activities of BW006 on human PBMCs. Human PBMCs were treated with or without CpG ODNs (6 μg/mL) for 36 hours followed by [3H] TdR incorporation for 16 hours and the cell proliferation was measured by 3H-thymidine incorporation and expressed as mean cpm±SD by columns. For detecting the expression levels of CD69 and HLA-A2, the PBMC cultured with or without CpG ODNs for 18 hours were stained with FITC-conjugated anti-CD69, HLA-A2, and PE-conjugated anti-CD19, CD56, or CD8 mAb and then analyzed on a FACS.
Because of the requirement for evaluating the adjuvant activity of BW006 for HBsAg in mouse models, we next tested whether BW006 could also stimulate the proliferation of murine splenocytes, activate murine immune cells, and upregulation of MHC class I molecule of the cells. In proliferation assay, splenocytes of BALB/c mice were stimulated by BW006 and control CpG ODN, including 1585 (a mouse A-type CpG ODN), 1826 (a mouse B-type CpG ODN), and C274 and then tested for their incorporation of [3H] TdR. The result showed that BW006 could stimulate the proliferation of the splenocytes to a level similar to that induced by C274 and a level much higher than that induced by 1826 (p<0.05) and 1585 (p<0.001) (Fig. 2A). To test the ability of BW006 for activating mouse immune cells, mouse splenocytes were stained by fluorescence-conjugated mAb after CpG ODN stimulation and analyzed by flow cytometry. The result demonstrated that BW006 could upregulate CD69 expression in CD19+, CD56+, or CD8+ cells in a similar capacity as that induced by 1826 (p>0.05) (Fig. 2B), and stimulate the upregulation of IAb1 molecules in mouse splenocytes. The results reveal that adjuvanticity of BW006 could be evaluated in mice.
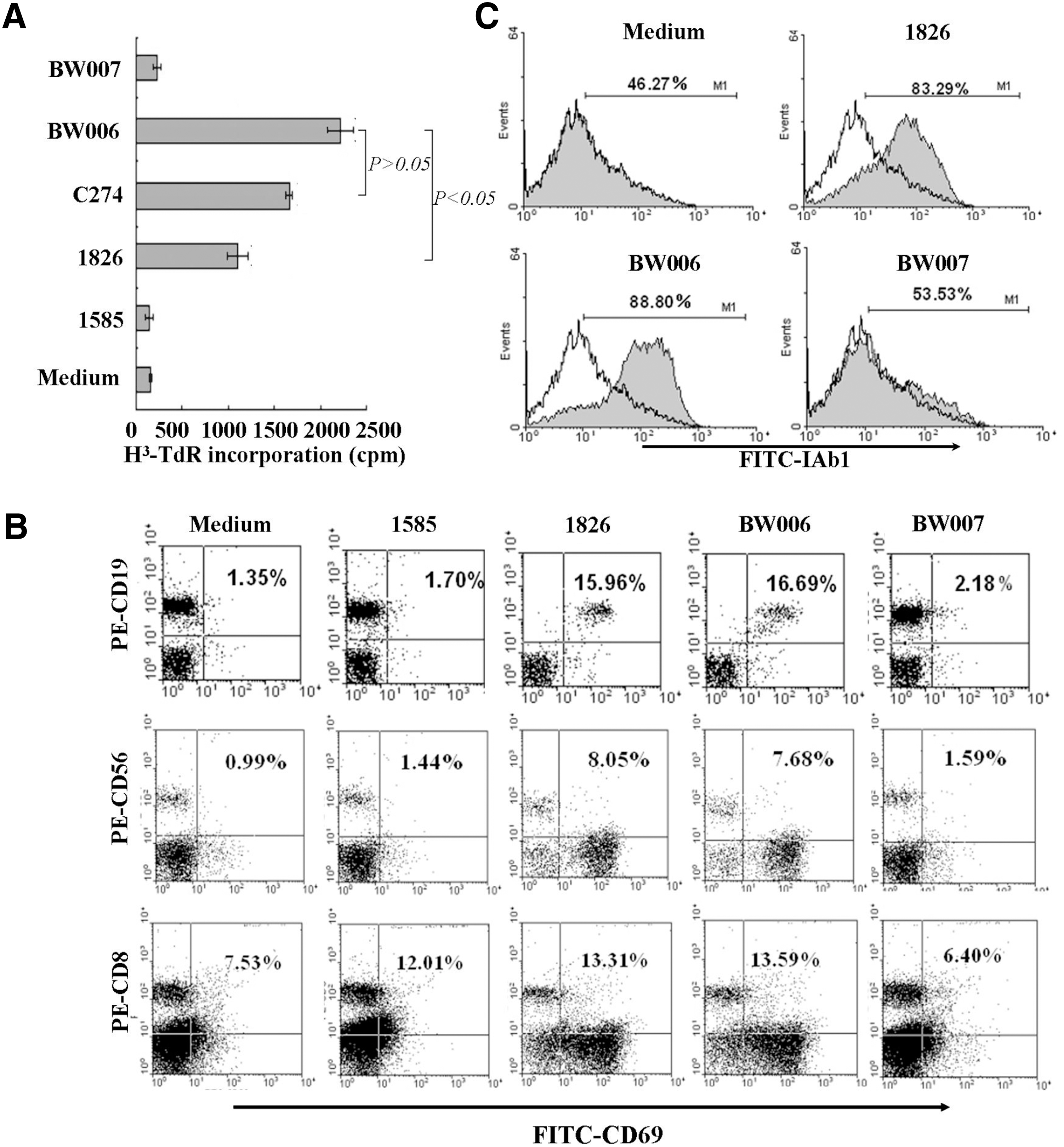
Immunostimulatory activities of BW006 on mouse splenocytes. Mouse splenocytes were cultured with or without CpG ODNs (6 μg/mL) for 36 hours followed by [3H] TdR incorporation for 16 hours and the cell proliferation was measured by 3H-thymidine incorporation and expressed as mean cpm±SD by columns. For detecting the expression of CD69 and IAb-1, splenocytes cultured with or without CpG ODNs for 18 hours were stained with FITC-conjugated anti-CD69, IAb-1, and PE-conjugated anti-CD19, CD56, or CD8 mAb and then analyzed on a FACS.
Effect of BW006 on assisting HBsAg to induce Th1-biased immune responses in mice
To investigate whether BW006 could facilitate HBsAg to induce a Th1-polarized immune response, mice were injected s.c. at inguinal lymph node areas in both sides with HBsAg alone or HBsAg+Alum or HBsAg+BW006 or HBsAg+Alum+BW006 and PBS, respectively, for four times at a 7-day interval. On day 7 after the last immunization, the mice were bled and HBsAg-specific IgG or IgG2a, a subtype Th1-biased antibody, or IgG1, a subtype Th2-biased antibody, in the sera were detected by ELISA. As shown in Figure 3A, the levels of IgG induced by HBsAg+BW006+Alum were the highest than those induced by the HBsAg+BW006 (p<0.05), HBsAg+Alum (p<0.001), and HBsAg alone (p<0.001), respectively. Whereas the IgG2a/IgG1 ratio induced by HBsAg+BW006 (1.02) was significantly higher than those induced by HBsAg+BW006+Alum (0.68) (p<0.05), HBsAg+Alum (p<0.001), and HBsAg alone (p<0.001), respectively. The data indicated that the addition of BW006 to HBsAg resulted in a shift toward a Th1-biased immune response. Next, we measured IFN-γ and IL-12 production induced by BW006-adjuvanted HBsAg. Mice were immunized as described above. On day 7 after the last immunization, splenocytes were isolated and cultured with HBsAg (50 μg/mL) for 24 hours. The levels of IFN-γ and IL-12 in the supernatants were determined by ELISA. As shown in Figure 3B, levels of IFN-γ and IL-12 in spleen cell cultures from the mice immunized with HBsAg+BW006 were not significantly different from those induced by HBsAg+Alum+BW006 (p>0.05) but much higher than those induced by HBsAg+Alum, HBsAg, or PBS (p<0.001). Thus, these data suggest that BW006 could induce a milieu in favor of a Th1-polarized immune response to HBsAg.
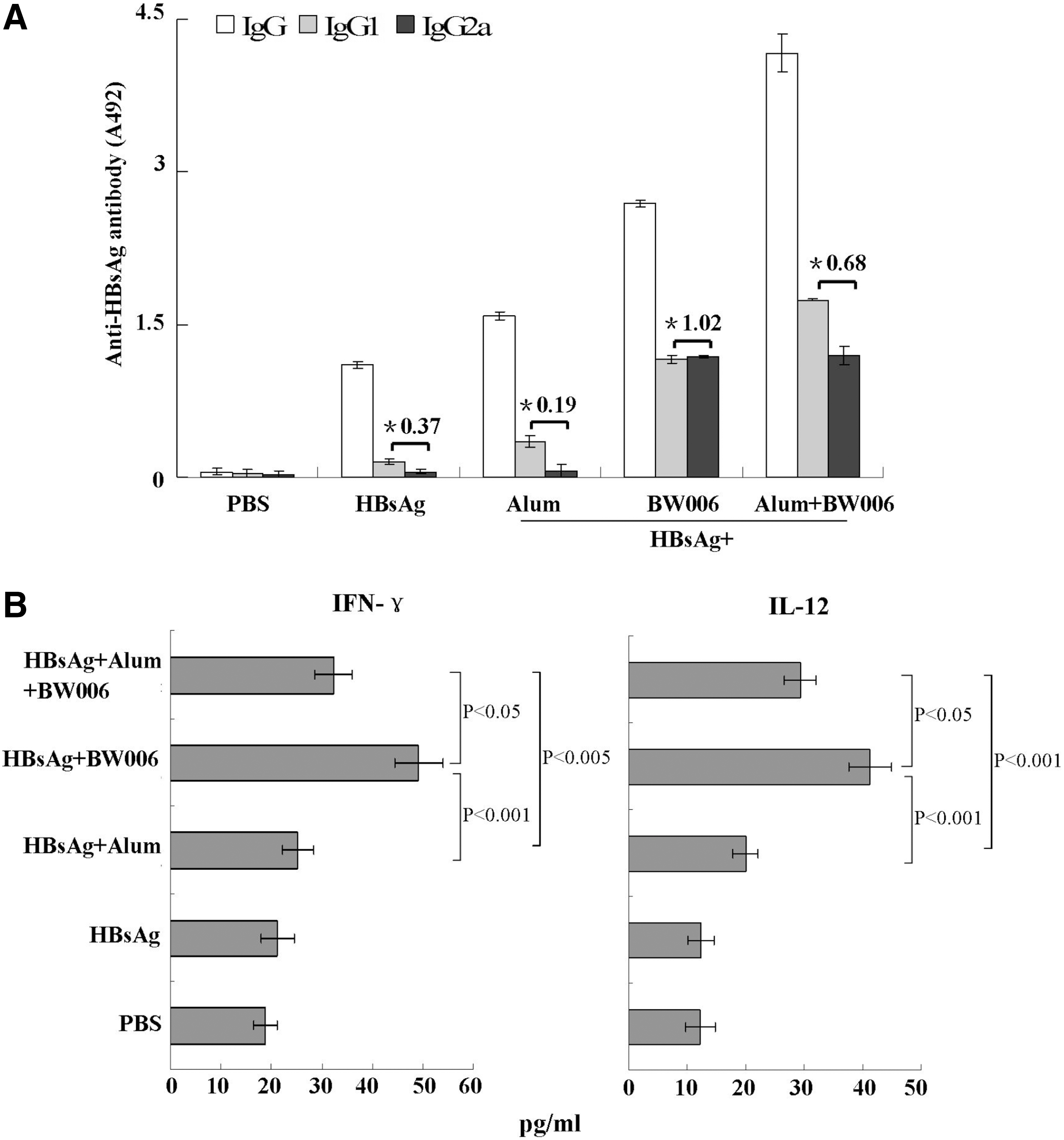
Th1 immune response induced by HBsAg+BW006 in mice. Mice (n=5) were immunized s.c. with HBsAg, HBsAg+Alum, HBsAg+BW006, or HBsAg+Alum+BW006, respectively, for four times at a 7-day interval. The mice injected by PBS as control. On day 7 after the last immunization, the mice were bled for detecting HBsAg-specific antibodies and splenocytes were isolated and stimulated with HBsAg (50 μg/mL) for 24 hours for testing the secretion of IFN-γ and IL-12 by ELISA.
Establishment of HBsAg-expressing liver cancer cells
To establish a HBsAg-expressing liver cancer cell line for assessing the efficacy of BW006-adjuvanted HBsAg inducing an immune response against HBsAg-expressing liver cancer, pcDNA3 plasmids carrying HBsAg-encoding gene were stably transfected into murine H22 liver cancer cells (H22 cells). The transfected cells, selected in a medium containing G418 (800 μg/mL) and through cloning by limited dilution, were identified by RT-PCR and Western blotting. As shown in Figure 4, a DNA fragment of 703 bp, representing HBsAg-specific mRNA, was amplified from the transfected cells (Fig. 4A), and proteins in a 28-kDa band, representing the expressed HBsAg, were recognized by anti-HBsAg mAb in the cell lysate of the transfected cells (Fig. 4B). The identified cells, designated as HBsAg+ H22 cells, were used for tumor inoculation and in vitro assays.

Identification of HBsAg+ H22 cell line.
Therapeutic effect of HBsAg+BW006 against HBsAg-expressing liver cancer cells in mice
To investigate whether BW006 could promote HBsAg to induce therapeutic immune response against HBsAg-expressing liver cancer cells, mice (n=8) were inoculated with HBsAg+ H22 cells (2×106/mouse) on day −1 and then s.c injected with PBS or HBsAg or HBsAg+Alum or HBsAg+BW006 or HBsAg+Alum+BW006 at inguinal lymph node areas in both sides on days 0, 7, 14, and 21, respectively. The mice were monitored for measuring tumor volumes every other day and recoding the survival. The result showed that HBsAg+BW006 significantly inhibited the growth of HBsAg+ H22 cells compared with PBS (p=0.028); however, HBsAg+Alum or HBsAg+Alum+BW006 failed to induce tumor growth inhibition (p>0.05) (Fig. 5A). Interestingly, the established tumors were completely rejected in 2 out of 8 mice treated with HBsAg+BW006 (Fig. 5A). By day 90 post-tumor challenge, all of mice received with PBS or HBsAg+Alum or HBsAg+Alum+BW006 had succumbed to the implanted HBsAg+ H22 cells and the average survival of mice of all groups was 61–64 days. In contrast, by day 64 and 90 post-tumor implantation, 87.5% and 37.5% of mice in HBsAg+BW006 group were still alive (Fig. 5B). These results indicated that therapeutic administration of HBsAg+BW006 induced effective immune responses against HBsAg-expressing liver cancer cells in mice.
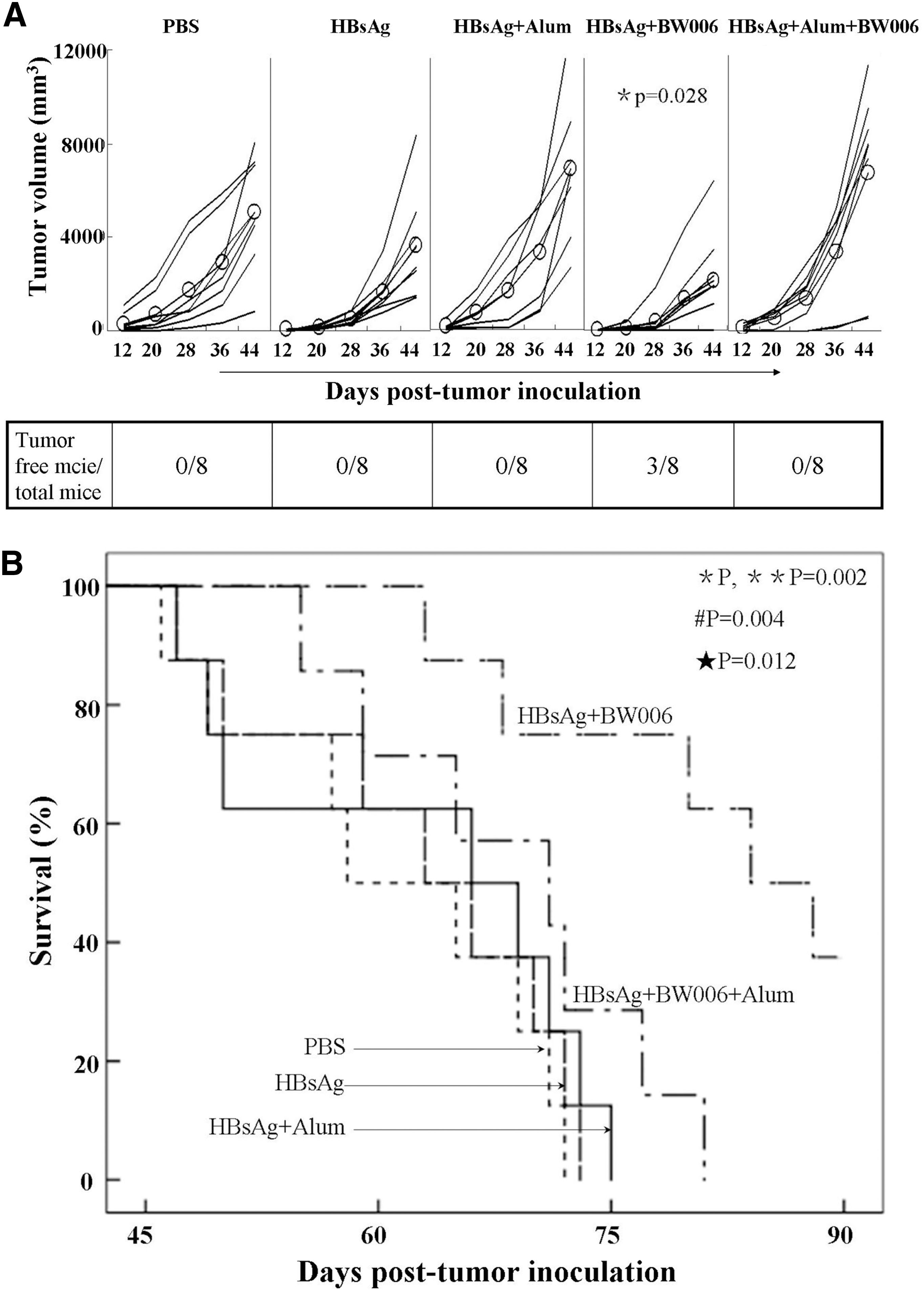
Therapeutic effect of HBsAg+BW006 on anti-HBsAg-expressing liver cancer cells in mice. Female BALB/c mice (n=8) were inoculated with 2×106 HBsAg+ H22 cells on day −1 and s.c. injected with HBsAg, HBsAg+Alum, HBsAg+BW006, HBsAg+Alum+BW006, or PBS on days 0, 7, 14, and 21, respectively. Tumor volume was measured every other day and survival of mice was recorded. *, versus PBS; **, versus HBsAg; #, versus HBsAg+Alum; ★, versus HBsAg+BW006+Alum.
CTL induced by HBsAg+BW006 in mice
To test whether HBsAg+BW006 induced therapeutic effect on HBsAg-expressing liver cancer was associated with generation of HBsAg-specific CTL response, mice (n=5) were immunized as described above. The immunized mice were sacrificed on day 7 after the last immunization and their spleen cells were isolated. The cells were incubated with HBsAg for 2 days, and then cocultured with HBsAg+ H22 cells at effector/target ratio of 12.5:1, 25:1, and 50:1, respectively, for 12 hours. The living HBsAg+ H22 cells were stained by trypan blue and counted. It was found that at the E:T ratios 25:1 and 50:1, the CTLs from mice immunized by HBsAg+BW006 lysed HBsAg+ H22 and failed to lyse wild-type H22 cells (Fig. 6). The result showed that HBsAg+BW006 could induce the generation of CTL specific to HBsAg+ H22 cells.

HBsAg-specific cytotoxic T lymphocyte response induced by HBsAg+BW006. Mice (n=5) were injected s.c. with HBsAg or HBsAg+Alum or HBsAg+BW006 or HBsAg+Alum+BW006 or PBS, respectively, for four times at a 7-day interval. On day 7 after the last immunization, splenocytes isolated from the immunized mice were cultured with HBsAg (50 μg/mL) for 2 days to generate effector cells. HBsAg+ H22 cells were used as target cells. Percentages of specific lysis are shown at different E:T ratios. The mean±SD of triplicate determinations is shown. *, versus PBS; **, versus HBsAg; #, versus HBsAg+Alum; ★, versus HBsAg+BW006+Alum.
Discussion
In this study, we found that HBsAg+BW006, not HBsAg+BW006+Alum, induced therapeutic immunity against HBsAg-expressing liver cancer cells in mice, indicating that BW006 itself could be applied as an adjuvant to enhance the immunogenicity of HBsAg as a tumor associate antigen.
The therapeutic effect of HBsAg+BW006 might be correlated to that BW006, as a B-type CpG ODN, could assist HBsAg to generate specific cellular immune responses to eliminate HBsAg-expressing tumor cells via the following ways. First, BW006 could promote HBsAg to be processed and cross-presented through MHC class I pathway, therefore facilitating the generation of HBsAg-specific CTLs. The assumption was supported by our in vitro evidence that BW006 enhanced the expression level of MHC I molecules of human PBMC (Fig. 1C) and mouse spleen cells (Fig. 2C), and also by the results from other group that CpG DSP, a B type CpG ODN, upregulated expression of MHC I molecules in malignant B cells from patients with chronic lymphocytic leukemia, enabling the malignant B cells to present their tumor antigens to CTLs and to be promptly killed by the CTLs. 30 Second, BW006 could provide a Th1-favoring milieu for HBsAg by stimulating the production of IL-12 and IFN-γ. It is established that IL-12 produced by TLR9-expressing B cells or plasma dendritic cells 10,31 can promote CD4+ T cells differentiation into Th1 CD4+ T cells, 32,33 which may allow the CTLs to persist and expand at the tumor site to eliminate antigen-specific tumor cells. 34 In addition, IL-12 induced by BW006 could stimulate immune cells to produce IFN-γ as shown in Figure 3B. Probably, the elevated IFN-γ in mice treated with the HBsAg+BW006 could upregulate the expression of MHC class I molecules in HBsAg-expressing liver cancer cells. Theses alterations, in turn, could render the HBsAg-expressing liver cancer cells more immunogenic and susceptible to CTL lysis. Similar observation was reported that the coadministration of CpG ODN (1826) and IL-12 gene-modified tumor vaccine induced potent anti-tumor Th1 immune response through the generation of IFN-γ. 12 Moreover, the IFN-γ can also enhance the anti-tumor effect of tumor vaccine by promoting the generation of Th1 CD4+ T cells, as confirmed by that CpG ODN (1826) enhanced the effect of RMA peptide on inducing immunity against RMA tumor in mice by inducing an IFN-γ-dependent CD4 cell immunity. 35 Furthermore, the local release of IFN-γ has been shown to interfere with tumor growth by inhibiting the vascularization of the tumor. 36,37 Third, BW006 could activate NK cells to kill the HBsAg-expressing liver cancer cell debris that could be taken up by DCs and then presented to CTLs, resulting in HBsAg-specific CTL responses. In addition, once activated by BW006, NK cells might upregulate their Fcγ receptors to mediate an enhanced antibody-dependent cellular cytotoxicity against tumor cells. The high level of IgG2a induced by HBsAg+BW006 could coat the tumor cells and make the tumor cells more susceptible to NK cell-mediated killing. The CpG ODN enhanced NK cell-mediated killing was also reported by the results showing that CpG ODN (2216) enhanced antibody-dependent cell cytotoxity (ADDCC) mediated by MUC1-specific monoclonal antibody against MUC1-positive breast cancer cells. 38
Unexpectedly, we found that the therapeutic effect of the BW006-adjuvanted HBsAg on HBsAg-expressing liver cancer was somehow masked by Alum, manifested by that HBsAg+BW006 could significantly inhibit the growth of HBsAg-expressing liver cancer cells and prolong the survival of mice bearing HBsAg-expressing liver cancer but HBsAg+BW006+Alum failed to. Presumably, the counteraction could be correlated to the enhanced production of Th2 cytokines induced by Alum. In this study, we found that HBsAg+BW006 induced vigorous Th1-biased immune response (IgG2a/IgG1=1.02) with the elevated production of IL-12 and IFN-γ. However, the Th1-biased immune response was shifted to Th2 immune response (IgG2a/IgG1=0.68) when Alum was coadministered with BW006-adjuvanted HBsAg. Generally, the Th2 cytokines, including IL-10 and IL-4, can create unfavorable surroundings for the generation of CTL responses by inhibiting the production of Th1 cytokines such as IL-12 and downregulating MHC I and MHC II molecules, which are crucial for tumor antigen presentation and recognition. 39
Taken together, BW006 is optimal for assisting HBsAg to induce immune response against HBsAg-expressing liver cancer cells in mice. The data may be of value in developing a therapeutic vaccine against HBsAg-expressing liver cancer cells for human use.
Footnotes
Acknowledgments
The authors gratefully acknowledge the financial support from Hi-Tech Research & Development Program of China (2006AA02Z487), Enterprise Promoting Program by Universities, Jilin, China (2009GJB10015), and Graduate Innovation Fund of Jilin University (20091031).
Disclosure Statement
The authors confirm that all the listed authors have no competing financial interests.