
Abstract
Hepatocellular carcinoma (HCC) is often diagnosed at an advanced stage, and over the past several decades, many researchers have worked to develop novel effective therapies for HCC patients. The functional contributions of mesenchymal stem cells to human malignancies, including HCC growth and progression, are controversial, and the potential mechanisms underlying these effects are not clear. The aim of this study was to investigate the effect of adipose-derived mesenchymal stem cells (ADSCs) on the growth of HCC cells. In this study, a conditioned medium from ADSCs (ADSC-CM) efficiently inhibited HCC cell proliferation and division, and induced HCC cell death through the downregulation of Akt signaling. These findings indicated that the ADSC-CM could inhibit HCC growth. Thus, the ADSC-CM is a good candidate for the treatment of HCC patients for whom no effective therapy is available.
Introduction
Hepatocellular carcinoma (HCC) is the fifth most common cancer in men and the seventh in women. 1 HCC is a highly aggressive tumor, with a high rate of intrahepatic metastases, and it frequently spreads beyond the liver before it is first detected. 2 Therefore, HCC is associated with high mortality as a result of recurrence and advanced stage at detection. Early cancers can be treated by surgical removal (resection). However, the current surveillance techniques are not very good at detecting early liver cancers. Thus, clinicians are always searching for an efficient and feasible approach for unresectable patients.
Mesenchymal stem cells (MSC) can be isolated from many human tissues, such as bone marrow 3 and adipose tissue. 4,5 In spite of their similar characteristics, 6 adipose-derived mesenchymal stem cells (ADSCs) have attracted more extensive attention due to its advantages over bone mesenchymal stem cells (BMSC). Moreover, ADSCs were reported to be a source of therapeutic cells 7 that can be used for tissue engineering and regeneration. 8,9 Now, researchers are also focusing on the use of ADSCs in debilitating diseases such as myocardial infarction, 10 diabetes mellitus, 11 and neurodegenerative diseases. 12,13 ADSCs can also differentiate into hepatocytes under certain conditions, and these cells have protective potential in liver injury. 14 Furthermore, ADSCs have gained interest due to their relationship with carcinoma, though the functional effects of ADSCs on tumor growth and progression are still not clear 15 –17 : ADSCs can be recruited by prostate cancer cells and promote tumor growth through increasing tumor vascularity. 18 Furthermore, ADSCs support breast tumor growth and progression, 19,20 but can block the G1–S phase transition, and subsequently inhibit proliferation of pancreatic cancer cells in vitro and in vivo. 21 It is interesting that the reported functions of MSC in the progression of HCC are also controversial: a conditioned medium from MSC enhanced tumor growth, but significantly inhibited the invasiveness and metastasis of HCC 22 ; Hanno Niess et al. found that systemically injected MSC promoted orthotopic HCC growth 23 ; however, MSC exhibit intrinsic inhibition of HepG2. 24 Unfortunately, there are few reports about the effect of ADSCs on HCC.
The purpose of the present investigation was to study the effects of the conditioned medium from ADSCs (ADSC-CM) on the growth of HCC cells and explore the underlying mechanisms. If the ADSC-CM exerts an inhibitory effect on HCC cells, it could potentially be used to treat currently incurable HCC patients.
Material and Methods
ADSC isolation and culture
Human adipose tissue was collected after liposuction surgery from the Cosmetic and Plastic Surgery Centre of the affiliated Zhongshan Hospital of the Xiamen University. The study was approved by Human Research Ethics Committee of the hospital. The raw lipoaspirates were washed, digested with 0.01% collagenase I (Sigma), and passed through a 40-μm mesh. Then, erythrocytes were eliminated using Red Blood Cell lysis buffer (Beyotime), and the remaining cells were cultured in plastic plates with DMEM supplemented with 10% fetal bovine serum (FBS; Gibco). Cells at passages 3–6 were used for experiments.
Conditioned medium from ADSCs
The conditioned medium was harvested from cultured ADSCs after incubation in a serum-free DMEM for 24 hours. At the end of the incubation period, the medium was filtered and used immediately, or stored at −80°C.
HCC cell lines and culture
The human hepatic cancer cell lines HepG2, Huh7, SMMC7721, and Bel7402 were purchased from Shanghai Cell Bank, Chinese Academy of Sciences. HepG2, Huh7, SMMC7721, and the multidrug resistant cell line SMMC7721/ADM were cultured in plastic plates with the DMEM supplemented with 10% FBS. Bel7402 cells were cultured with RPMI-1640 (Hyclone) supplemented with 10% FBS.
Flow cytometry
ADSCs were trypsinized, and 1×106 ADSCs were incubated with PE-CD90, PE-CD105, PE-CD45, and PE-CD34 (all from eBioscience). The fluorescently labeled cells were analyzed on a flow cytometer (Beckman) using CELLQuest Pro software. The appropriate isotype control antibodies were also used.
In vitro adipogenic and osteogenic differentiation
Adipogenic and osteogenic differentiation assays were performed using the Human Mesenchymal Stem Adipogenic or Osteogenic Differentiation Medium Kits (Cyagen Biosciences) according to the manufacturer's instructions. Adipogenic differentiation was confirmed by the formation of neutral lipid vacuoles stainable with Oil Red O (Sigma), and osteogenesis was assessed by Von Kossa staining. Noninduced cells were used as controls.
Cell viability assay
HCC cells were cultured in triplicate at 4×103 cells/well in 96-well plates and allowed to adhere overnight. The culture medium was then replaced with the ADSC-CM containing 10% FBS for 3 days. Cell viability was evaluated by using the BrdU Cell Proliferation Kit (Chemicon) as described previously. 25 Absorbance was measured at 450 nm using a microtiter plate reader (Bio-Rad). The absorbance values directly correlated with the amount of DNA synthesis, and thus with the number of proliferating cells in the culture.
TUNEL assay
Cells were harvested after incubation with the ADSC-CM for 24 hours, and then DNA fragmentation from the apoptotic cells was evaluated with a terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nicked-end labeling (TUNEL) assay using the FragEL DNA Fragmentation Detection Kit (Calbiochem). The apoptosis index was calculated by counting the number of TUNEL-positive cells in five randomly selected high-power fields (×400 magnification).
EdU incorporation assay
Cell proliferation was assessed using the Cell-Light 5-ethynyl-20-deoxyuridine (EdU) DNA Cell Proliferation Kit (RiboBio) according to the manufacturer's instructions. Images were captured and analyzed using a Leica DM2500 fluorescence light microscope. The number of EdU-positive cells was calculated as [(EdU add-in cells/Hoechst-stained cells)×100%] in five arbitrarily selected fields from each group at×200 magnification.
Western blot analysis
Cells were harvested and lyzed in an RIPA buffer and centrifuged to remove debris. Cell extracts were resolved on 10% SDS–polyacrylamide gels and transferred to polyvinylidene difluoride membranes (Millipore). The membranes were blocked and then incubated with primary antibodies overnight at 4°C. Anti-Akt, antiphosphorylated Akt, anti-Bcl-2, anti-Bax, anticaspase-3, and anti-Bcl-xl were purchased from Cell Signaling Technology. HRP-conjugated secondary antibodies were added, and the blots were incubated for 1 hour at room temperature. The immunoreactive products were then detected using an Enhanced Chemiluminescence kit (Pierce Thermo Scientific) and quantified by densitometry. β-actin (Sigma) was used as a loading control. Densitometric analyses of the bands were performed in Gel Pro Analyzer 4.0.
CFSE proliferation assay
The stable incorporation of the intracellular fluorescent dye 5-(and -6)-carboxyfluorescein diacetate succinimidyl ester (CFSE) into cells provides a powerful tool for monitoring cell migration and quantifying cell division due to the sequential decrease in fluorescent labeling in daughter cells. HCC cells were stained with CFSE at a final working concentration of 10 μM, followed by incubation at 37°C for 15 minutes. The staining was quenched, and stained cells were cultured with or without the ADSC-CM for 48 hours. The cells were then harvested and analyzed by flow cytometry. FACS data were analyzed using CellQuest and ModFit software.
Cell apoptosis assay
Cells were harvested after incubation with the ADSC-CM for 48 hours and then stained with AnnexinV-FITC and propidium iodide (PI) according to the instructions provided with the AnnexinV/PI Apoptosis Detection Kit (Keygen). Then, cells were analyzed by flow cytometry using CELL Quest Pro software.
Statistical analysis
All data were analyzed using SPSS software (version 13.0). The results were expressed as the mean±SD. Statistical analyses were performed using one-way ANOVA and Student's t-test. The significance level was set at 0.05.
Results
Characteristics of cells derived from adipose tissue
The fibroblastoid cells derived from adipose tissue formed a monolayer, 3 days after the initial plating. After depletion of the nonadherent cells, a primary culture was established, and the fibroblastoid cells were termed ADSCs. ADSCs proliferated slowly, but continuously. After repeated differential trypsinization and passaging, cells exhibited homogeneous shapes (Fig. 1A). To determine the immunophenotype of the isolated cells, a panel of surface markers was analyzed by flow cytometry. As reported, ADSCs strongly expressed the markers of MSC CD105 and CD90, and were negative for the hematopoietic surface markers CD34 and CD45 6,7 (Fig. 1B). Furthermore, ADSCs were injected subcutaneously into nude mice (5×106 cells per mouse; n=10) to evaluate their security. Mice injected with ADSCs did not develop any tumors over 3 months of observation.

Characterization of adipose-derived mesenchymal stem cells (ADSCs).
Multilineage differentiation capacity of ADSCs
Next, we evaluated the adipogenic and osteogenic potentials of fibroblastoid cells derived from adipose tissue. To induce adipocyte differentiation, cells were cultured in an adipogenic medium, and adipogenic differentiation was confirmed by Oil Red O staining, which marks intracellular lipid droplets (Fig. 2, upper panel). Osteogenic differentiation was suggested by the observation of morphological changes and the appearance of calcium crystals in the culture. Osteogenic differentiation was further confirmed by Von Kossa staining of mineralization of the matrix (Fig. 2, lower panel).
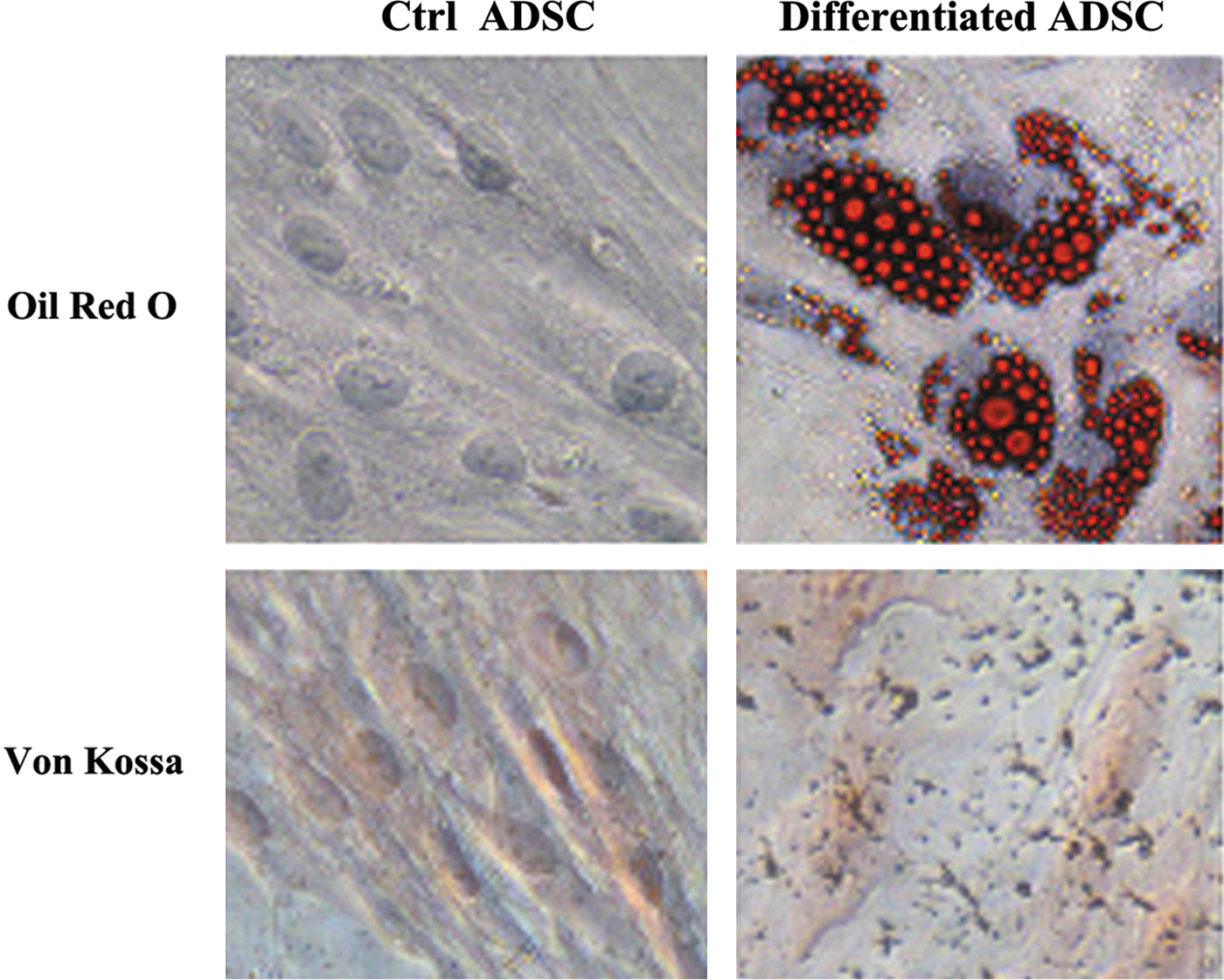
The multilineage differentiation potential of ADSCs was investigated. Adipogenesis was demonstrated by Oil Red O staining, and osteogenesis was demonstrated by Von Kossa staining. Cells were cultured in either the control (left panel) or differentiation medium (right panel). Ctrl, control. Color images available online at
ADSC-CM inhibits the HCC cell proliferation and division
To analyze the effects of the ADSC-CM on HCC cell proliferation, we performed a cell viability assay. Several HCC cell lines, including HepG2, Huh7, SMMC7721, Bel7402, and the multidrug-resistant cell line SMMC7721/ADM were incubated with or without the ADSC-CM. As shown in Figure 3A, the relative cell viability was decreased when HCC cells were cultured with the ADSC-CM. Moreover, SMMC7721 cell viability was dramatically suppressed (Fig. 3A, p<0.001). We further conducted an EdU incorporation assay to evaluate the effect of the ADSC-CM on the proliferation of SMMC7721 cells. As shown in Figure 3B, the proportion of EdU-positive cells was decreased after the ADSC-CM treatment compared with the control medium (DMEM) treatment. Furthermore, the proportion of cells incorporating EdU was decreased by 50.5%±2.9% (Fig. 3C, p<0.05).

The effect of the conditioned medium from ADSCs (ADSC-CM) on the proliferation of human hepatic cancer cell lines.
Meanwhile, to further confirm the inhibitory effect of the ADSC-CM, we also assessed its effect on HCC cell division. Hepatoma SMMC7721 cells were labeled with CFSE and then cultured with the ADSC-CM for 48 hours. A halving of CFSE fluorescence indicates a single round of cell division. As was shown in Figure 3D, in the cells fixed with 4% paraformaldehyde after labeling with CFSE, which were regarded as parent cells (P0), no cells underwent division, and the proliferative index (PIX) was 1.00 (blue). Cells cultured with the ADSC-CM resulted in a significant decrease in PIX relative to controls cultured with the DMEM only (Ctrl, PIX=12.81 vs. ADSC-CM, PIX=5.58). In the control group, 89.62% of cells had completed five divisions (Cambridge blue). However, in the ADSC-CM treatment group, most cells had undergone only three or four divisions: 74.12% of cells were in the fourth division (violet), and 18.22% of cells were in the third division (grass green). Taken together, these data suggested that the ADSC-CM could inhibit HCC cell proliferation.
ADSC-CM induces hepatoma SMMC7721 cell apoptosis
Next, we tried to investigate whether SMMC7721 cells cultured with the ADSC-CM undergo apoptosis. Using Annexin V/PI double-staining-based flow cytometry analysis, we found that the ADSC-CM strongly increased the percentage of apoptotic cells (Annexin V-positive), from 14.4%±2.7% in the ADSC-CM-treated group to 5.8%±2.5% in the control group. The levels of cell apoptosis were elevated by ∼2.5-fold in cells cultured with the ADSC-CM versus the control medium (p<0.05, Fig. 4A). We also assessed SMMC7721 cell apoptosis by the TUNEL assay. The number of TUNEL-positive cells was upregulated in the ADSC-CM group compared with the control group. (p<0.01, Fig. 4B). Further experiments revealed that the ADSC-CM elevated the expression of cleaved-caspase-3 (p<0.05, Fig. 4C), a well-known member of the apoptosis pathway. Taken together, our data indicated that the ADSC-CM suppressed HCC cell growth through inhibiting cell proliferation and enhancing cell apoptosis.
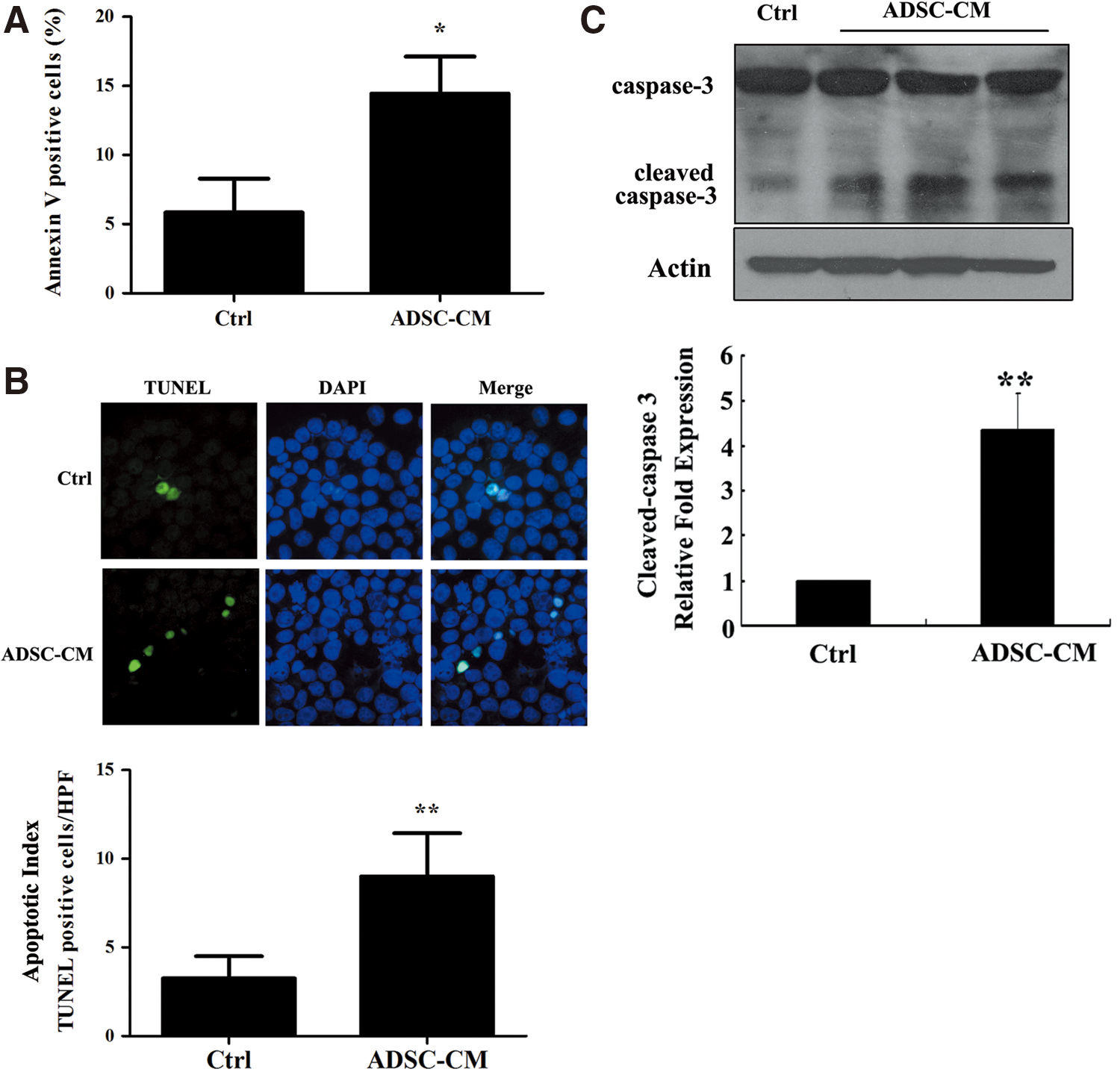
ADSC-CM initiates apoptosis of SMMC7721 cells. Cells were cultured with the ADSC-CM or the control medium for 24 hours and then collected.
HCC cell apoptosis induced by the ADSC-CM is mediated by the downregulation of Akt signaling
We next investigated the mechanisms involved in the effects of the ADSC-CM on SMMC-7721 hepatoma cells. The activation of Akt and ERK1/2 signaling is involved in the growth of many types of cancer cells, including HCC cells. Western blot analysis revealed that stimulation with the ADSC-CM induced a strong inhibition of Akt phosphorylation relative to the control (Fig. 5A, p<0.05), but did not obviously affect the phosphorylation of ERK1/2 (data not shown). Meanwhile, the expression of the proapoptosis protein Bax was upregulated, whereas the prosurvival proteins Bcl-2 and Bcl-xl were downregulated by ADSC-CM treatment (Fig. 5B).
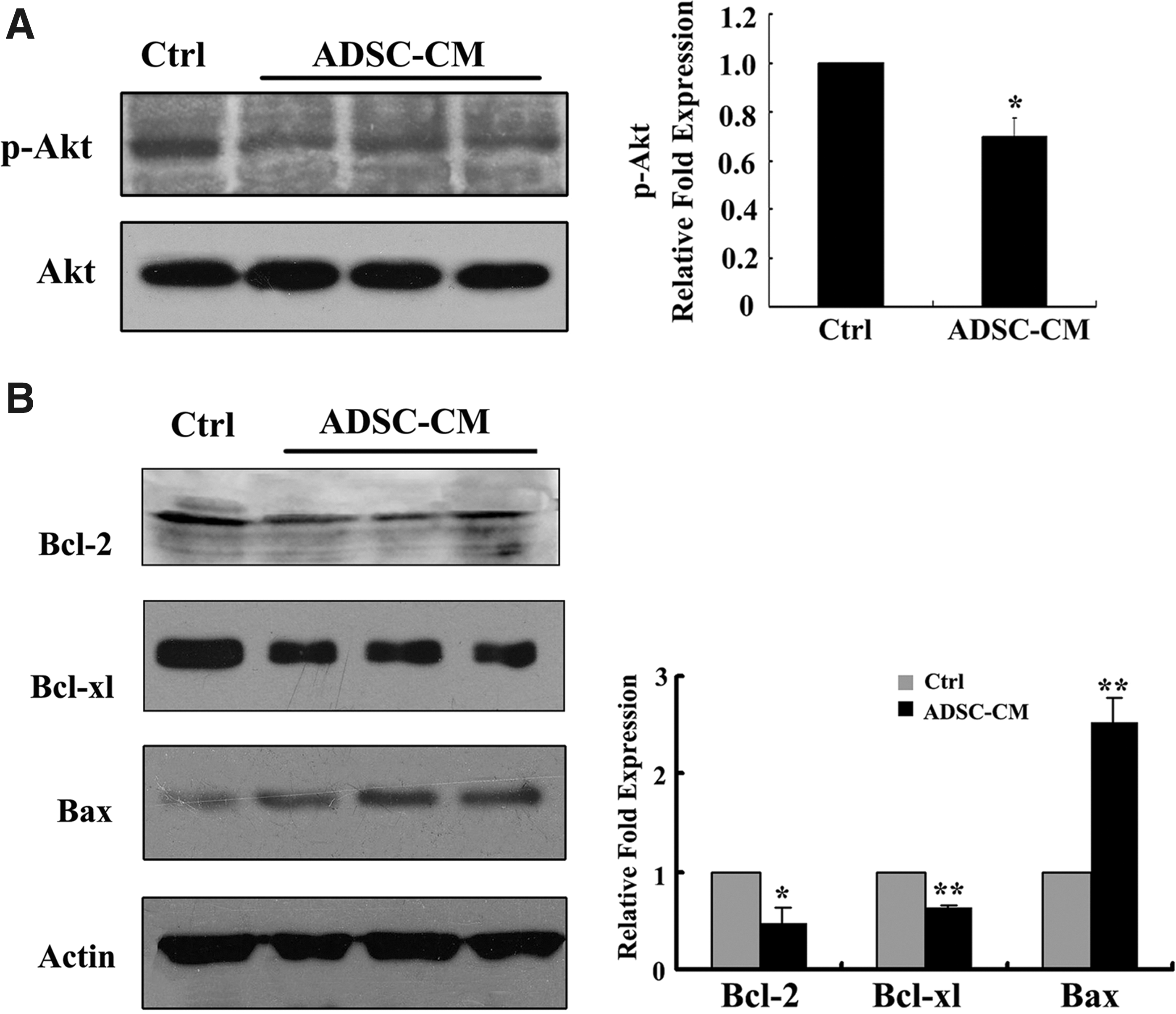
The mechanism by which the ADSC-CM induced hepatoma SMMC7721 cell apoptosis. SMMC7721 cells were harvested after incubation with the ADSC-CM from three different donors or the control medium for 48 hours. Cell lysates were prepared and analyzed by western blot to detect phospho-Akt
Discussion
Our findings demonstrated that the ADSC-CM efficiently inhibited HCC cell proliferation and promoted HCC cell death in vitro. Our data also suggest a candidate molecular pathway for the suppression of in vitro HCC progression, namely the downregulation of Akt signaling. Therefore, ADSCs may provide a cell-based therapeutic alternative for patients suffering from HCC.
ADSCs remain difficult to define due to the lack of definitive cellular markers. Although ADSCs and BMSC share many characteristics, 6 they are fundamentally different cell types and are differentially committed cells. 26 We attempt to clearly discriminate between these cells and fibroblasts using four markers: CD34- (endothelial cells, CD34+), CD45− (hematopoietic cells, CD45+), CD105+ (vascular smooth muscle cells or pericytes, CD105−; MSC, CD105+), and CD90+ (MSC, CD90+). 6,26 –29
ADSCs have been reported to exhibit contradictory properties in cancer biology. 18 –21,30 –32 ADSCs possess several features that make them be recruited to the tumors 33,34 and then sustain cancer cells in their ability to survive the multifaceted host response against cancer. Interestingly, the effects of BMSC on tumor proliferation also vary with the tumor type: BMSC facilitate melanoma cancer cell proliferation 35 and breast cancer metastasis, 36 but suppress the growth of Kaposi's sarcoma cells. 37
According to our findings, the ADSC-CM inhibited proliferation and induced apoptosis of HCC cells in vitro. Akt is involved in the apoptosis mechanism, which is an important cell survival factor in various cell types, particularly cancer cells. The activation of PI3K/Akt signaling has been established as critical for tumor cell growth and survival in numerous cancers. 38 HCC proliferation is associated with multiple signaling pathways, 39 including Akt and ERK1/2. Akt phosphorylation at Ser473 has been implicated in early HCC recurrence and poor prognosis. 40,41 In our study, Akt phosphorylation on Ser473 was downregulated by ADSC-CM treatment, suggesting that the ADSC-CM inhibited HCC cell growth by blocking Akt signaling. However, ERK1/2 activity was not obviously changed (data not show). The precise molecular mechanisms by which ADSCs inhibit Akt require further research.
Our research was based on the use of a conditioned medium, suggesting that the paracrine activity of ADSCs may exert pro- or antitumor progression effects. At present, there are no conclusive clinical data showing that adipocyte-derived factors promote carcinogenesis. 42 Adipose lineage cells secrete a broad spectrum of bioactive cytokines, such as HGF, IGF-1, IGFBPs, TGF-β, TNF-α, MCP-1, RANTES, TIMP-1, TIMP-2, interleukin-6, interleukin-8, and hormones (e.g., leptin), 33,43,44 which may affect tumor angiogenesis and host immune responses and modulate the tumor microenvironment in cancer. Certain cytokines may play an important role in our experiments, which do not involve any cell–cell contact.
Akt activation can stimulate cell proliferation through multiple downstream targets that impinge on cell cycle regulation and the activation of mTOR (the mammalian target of rapamycin). 39 Akt-dependent phosphorylation of GSK3 is likely to drive cell proliferation through the synthesis of proteins involved in the cell cycle entry. Activated Akt prevents p27 from entering the nucleus and losing the ability to inhibit CyclinD1 and CDK2. 45 The levels of phosphorylated mTOR have been shown to be elevated in 15% of HCC cases, and the levels of total p70 S6 kinase (the direct substrate of phosphorylated mTOR) have been shown to be increased in 45% of cases. 46 Furthermore, aberrant mTOR signaling is associated with the IGF cascade, which regulates several cellular processes, including proliferation, motility, and the inhibition of apoptosis. Additionally, HGF, an ADSC paracrine growth factor, binds to the high-affinity tyrosine kinase receptor c-MET and activates it, regulating cell proliferation, migration, survival, branching morphogenesis, and angiogenesis. 39 Therefore, further research targeting IGF-activated AKT/mTOR and HGF/c-MET signaling or other possible mechanisms for the growth of HCC cells will be explored in the next step of our work.
Our results indicated that the ADSC-CM inactivated the PI3K-Akt pathway, regulating the Bcl-2 family of inhibitory molecules, eclipsing stimulatory counterparts, and contributing to Akt-dependent caspase activation. Akt promotes cell survival by inhibiting apoptosis by phosphorylating and inactivating several targets, including Bad, caspase-9, 47 and caspase-3, 48,49 and simultaneously upregulates prosurvival proteins like Bcl-2 and Bcl-xl. 50 The proapoptotic protein Bax forms oligomers and translocates from the cytosol to the mitochondrial membrane and increases the membrane permeability, which leads to cytochrome c release from the mitochondria, the activation of caspase-9, and the initiation of the caspase cascades. 47 ADSC-CM-induced liver cancer cell apoptosis was confirmed by our immunofluorescence and TUNEL assays.
ADSCs have been used as cellular vectors to deliver proapoptotic molecules or ligands to a variety of tumor cells. 51,52 Our work represents an exploration of the use of nonengineered cells to treat HCC. Furthermore, ADSCs are easily obtained and amplified; therefore, large quantities of low-passage-number ADSCs and the conditioned medium are easily accessible. ADSCs may provide a promising and practicable reagent for HCC therapy. However, further in vivo studies of ADSCs or the ADSC-CM may provide a more comprehensive insight into its efficacy.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (No. 81171967), the National Key Sci-Tech Special Project of China (No. 2012ZX10002011-005), Projects of Xiamen Science and Technology Program (No. 3502Z20114010), and Public Health Department of Fujian Foundation for Young Scientists (No. 2009-2-65).
Disclosure Statement
There are no existing financial conflicts.