
Abstract
hTERTC27 is a newly constructed polypeptide that can induce telomere dysfunction. To study the synergistic antitumor effects of the hTERTC27 polypeptide driven by the early growth response protein-1 (Egr-1) promoter and chemotherapeutic 5-flurorouracil (5-FU) on nasopharyngeal carcinoma, a series of in vitro and in vivo experiments were performed. The results showed that hTERTC27 expression was significantly increased up to 7.21-folds by the 5-FU-activated Egr-1 promoter in C666-1 cells. Overexpressed hTERTC27 made the cells more sensitive to 5-FU, and additionally, inhibited cell proliferation about 20.41%. Combinational therapy of overexpressed hTERTC27 driven by the 5-FU-activated Egr-1 promoter and 5-FU synergistically inhibited cell proliferation and promoted apoptosis of C666-1 cells for about 4.75-fold and 1.76-fold in comparison with a sole therapy of hTERTC27 or 5-FU in vitro. In vivo experiments showed that overexpressed hTERTC27 driven by 5-FU-activated Egr-1 promoter and 5-FU synergistically reduced tumor volume, tumor weight, and local infiltration, which may be relative to tumor cell apoptosis. These results suggest that combinational therapy of overexpressed hTERTC27, which is driven by the 5-FU-activated Egr-1 promoter, and 5-FU may provide a novel approach to treat nasopharyngeal cancer.
Introduction
Nasopharyngeal carcinoma (NPC) is one of the most malignant neoplasms in the nasopharynx, especially in Southeastern Asia. 5-fluorouracil (5-FU), the most widely used anticancer agent, has been applied widely to the treatment of NPC. 1,2 However, 5-FU treatment has only limited benefit to the NPC patients because of severe side effects caused by high dosage and failing preventing recurrence and metastasis due to limited sensitivity. 3 Therefore, several strategies of gene therapy have been intensively explored to treat NPC. 4,5
Telomerase is an attractive target for cancer gene therapy, because it is highly active in most cancer cells, but it is inactive in most somatic cells. 6 The primary function of telomerase is to maintain the telomere length and to prolong the life span of cells. 7 The dysfunction of telomerase will prevent the long-term survival of tumor cells and lead to cell death or senescence. 8,9 Telomerase reverse transcriptase (TERT), the catalytic protein subunit of telomerase, is critical for the activity of telomerase in tumor cells. 10 Previously, we have reported that a 27-kDa C-terminal polypeptide of hTERT (hTERTC27) is capable of inducing telomere dysfunctions, and overexpression of hTERTC27 in human cervical carcinoma HeLa cells slows down the cell proliferation and inhibits the tumorigenicity by significantly increasing anaphase chromosome end-to-end fusion. 11 Ectopic overexpression of this polypeptide also makes the HeLa cells more sensitive to H2O2-induced oxidative injury. 12 Furthermore, hTERTC27 delivered by recombinant adeno-associated virus or recombinant adeno-associated virus and adenovirus cocktail system suppresses tumor growth in vivo in a human U87-MG glioblastoma xenograft model. 13,14
The early growth response protein-1 (Egr-1) promoter, which is sensitive to reactive oxygen intermediates, 15 is commonly used as a bridge in gene therapy, because it can be activated by oxygen free radicals generated by radiotherapy or chemotherapy and promotes the high expression of tumor therapeutic genes. 16,17 Therefore, we hypothesize that the radical agent 5-FU may activate the Egr-1 promoter, which can drive the overexpression of hTERTC27 that makes tumor cells sensitive to the chemotherapy of 5-FU, and have the chemotherapeutic effect on NPC.
In this study, hTERTC27 cDNA was constructed to the downstream of the Egr-1 promoter, which was transfected into the C666-1 cells. The synergistic antitumor effects of the 5-FU and hTERTC27 polypeptide driven by the 5-FU-activated Egr-1 promoter were assayed for the first time. Overexpressed hTERTC27 driven by the Egr-1 promoter made the C666-1 cells more sensitive to 5-FU and synergistically inhibited C666-1 cell proliferation and induced apoptosis with 5-FU treatment. These results suggest that the synergistic effect of 5-FU and Egr-1 promoter-driven hTERTC27 might be a potential clinical strategy for nasopharyngeal cancer therapy.
Materials and Methods
Cell culture
Human NPC C666-1 cells were maintained in our lab and cultured in the Dulbecco's modified Eagle's medium (DMEM; Hyclone), supplemented with 10% fetal bovine serum (FBS; Hyclone), 100 μg/mL penicillin (Ameresco), and 100 μg/mL streptomycin (Ameresco). Cells were cultured at 37°C in a humidified atmosphere with 5% CO2.
Plasmid construction and transfection
The pcDNA3.1-Egr-1 plasmid (pEgr) was a gift from Dr. Jianxiang Liu and Mei Tian. In this plasmid, the cytomegalovirus promoter was substituted with the Egr-1 promoter. cDNAs of a 27-kDa hTERT C-terminal polypeptide (hTERTC27, amino acid residues 882–1132) and enhanced green fluorescent protein (EGFP) were obtained by polymerase chain reaction using the pcDNA3.1-hTERTC27 and pEGFP-N2 (BD Biosciences) as templates, respectively. The hTERTC27 cDNA was subcloned into the pEgr plasmid at Kpn I and EcoR I sites to generate the pEgr-1-hTERTC27 plasmid (pEgr-C27). EGFP cDNA was then subcloned into the pEgr-C27 plasmid at EcoR I and Xba I sites to generate a pEgr-C27-GFP plasmid coexpressing hTERTC27 and GFP fusion protein. C666-1 cells were transfected with the pEgr, pEgr-C27, or pEgr-C27-GFP vectors using a Lipofectamine 2000 transfection reagent (Invitrogen), following the manufacturer's instructions, and selected using 600 μg/mL G418 (Merck) for 2 weeks. The cells stably transfected with the pEgr, pEgr-C27, or pEgr-C27-GFP vectors were called as C666-Con cells, C666-C27 cells, or C666-C27-GFP cells, respectively, and the unstransfected C666-1 cells were called as C666-Mock cells.
Detection of EGFP
About 2×106 C666-C27-GFP cells were seeded in 10-cm dishes. After 12 hours of incubation, the cells were treated with different concentrations (in μg/mL) of 5-FU (Sigma) of 0, 3.12, 6.25, 12.5, 25, respectively, for 48 hours. The cells were imaged with a fluorescence microscope (Zeiss). Then, cells were washed with phosphate-buffered saline (PBS) twice, harvested by trypsinization, and fixed in 70% ethanol. GFP expression was further determined by a flow cytometry analysis (Becton Dickinson).
Immunoblotting analysis
Cells were lyzed in an NP-40 lysis buffer containing 50 mM Tris (pH 7.4), 150 mM NaCl, 1% NP-40, 20 μg/mL PMSF, 1 μg/mL pepstatin A, and 1 μg/mL leupeptin (pH 7.4). Tumor tissue was homogenized in 250 mM sucrose containing 1 mM ethylenediaminetetraacetic acid, 20 μg/mL PMSF, 1 μg/mL pepstatin A, and 1 μg/mL leupeptin (pH 7.4) with a glass Dounce homogenizer. Total protein was assayed using a DC protein assay kit (BioRad). Proteins (30 μg) were electrophoresed through 12% (w/v) polyacrylamide and 1% (w/v) sodium dodecyl sulfate gels. Proteins were transferred from the gels by blotting onto a nitrocellulose membrane (Millipore). The membranes were blocked with 0.02 mol/L Tris–HCl (pH 7.0), 0.9% (w/v) NaCl, and 0.1% (v/v) Tween-20 containing 5% (w/v) nonfat milk for 1 hour at room temperature. Antibodies against the hTERT COOH-terminal fragment (1:250; Santa Cruz Biotechnology), cleaved caspase-3 (1:1000; Cell Signaling Technology), cleaved caspase-9 (1:1000; Cell Signaling Technology), cleaved PARP (1:1000; Cell Signaling Technology), and Bcl-2 (1:500; Santa Cruz Biotechnology) were added and incubated overnight at 4°C. Bound antibodies were located by a further incubation with horseradish peroxidase-conjugated goat anti-rabbit IgG (1:5000; Santa Cruz Biotechnology). β-actin (1:500; Santa Cruz Biotechnology) was used as a loading control. Blots were developed with an ECL Plus kit (Pierce) and exposed to Kodak films.
Determination of the 50% inhibitory concentration
About 2×103 C666-Mock, C666-Con, or C666-C27 cells were seeded in a 96-well plate. Twelve hours later, the cells were treated with different concentrations (in μg/mL) of 5-FU of 0, 0.78, 1.56, 3.12, 6.25, 12.5, 25, 50, 100, 200, or 400, respectively. An MTT assay was performed 48 hours later. Briefly, 20 μL of 5 mg/mL MTT (Sigma) in PBS was added, and the cells were then incubated for 4 hours at 37°C with 5% CO2. Cells were washed twice with PBS, and the pellet was then solubilized in 150 μL of 100% dimethyl sulfoxide (Sigma) by shaking for 5 minutes. Absorbance was measured on a microplate reader (BioRad) at a wavelength of 570 nm. The inhibitory rate (IR) and 50% inhibitory concentration (IC50) were calculated using Origin 7.5 software package.
Cell growth and proliferation assay
C666-Con or C666-C27 cells were seeded in six-well plates (2.5×104/well) and a 96-well plate (1.0×104/well). Twelve hours later, the cells were treated with PBS or 5-FU (6.25 or 12.5 μg/mL), respectively. Forty-eight hours later, cells in six-well plate were imaged with a light microscope (Zeiss) and then washed with PBS twice, harvested by trypsinization, and counted using a hemocytometer (Shanghai Jinghong Corporation). Cell proliferation in a 96-well plate was measured by the MTT assay as described above.
Colony formation assay
C666-Con or C666-C27 cells (5×102/well) were seeded in six-well plates. Twelve hours later, cells were exposed to PBS, 6.25, or 12.5 μg/mL 5-FU for 48 hours and allowed to grow for 14 days, fixed with methanol for 10 minutes, and stained with Giemsa for 15 minutes. The plates were washed and dried, and the colonies were counted. All experiments were done in triplicate.
Flow cytometry analysis of apoptosis
For quantitative analysis of apoptosis, cells were harvested at 48 hour after treating with 5-FU, washed once with ice-cold PBS, incubated with annexin V-fluorescein/PI (Boehringer Mannheim) in a calcium-containing HEPES (Sigma) buffer, and then immediately analyzed with a flow cytometry machine (Becton Dickinson).
TUNEL assay
The presence of apoptotic cells in the C666-1 cells was detected by the terminal deoxynucleotidyl transferase-mediated dUTP nick-end-labeling (TUNEL) assay using the in situ cell death detection kit (POD; Roche), according to the procedures recommended by the manufacturer. The apoptotic cells were analyzed under a fluorescence microscope.
Tumor xenograft formation and histological analysis
BALB/C nude mice at 6 weeks of age were purchased from the Medical Laboratory Animal Center of Guangdong Province, China. Mice were injected with 100 μL (5×107 cells/mL in PBS) of C666-Con cell suspension (12 mice) or C666-C27 cell suspension (12 mice) subcutaneously into the mid-back region. One week after injection, the mice inoculated with C666-Con or C666-C27 cells were randomized into two groups, respectively (Con, Con+5-FU and C27, C27+5-FU). Mice were administrated (i.p.) either saline or 5-FU (40 mg/kg daily, three times a day) for 5 days. Tumor xenograft formation was measured on a daily base to make the growth curves. At the end of the treatment, mice were sacrificed. Tumor xenografts were surgically dissected, weighed, measured and then fixed with 4% formaldehyde in PBS, and embedded in paraffin. Sections were stained with hematoxylin and eosin (HE staining). The tumor volume was calculated according to the equation L×W 2 /2 with the length (mm) being the longest axis of the tumor. This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the Medical Laboratory Animal Center of Guangdong Province, China (the permit number is SCXK2008-0002).
Statistical analysis
All data were presented as mean±SD. The results were compared by an analysis of variance. All statistical calculations were performed with SPSS 11.0 software package. A p-value<0.05 was regarded as statistically significant difference.
Results
5-FU increased hTERTC27-GFP expression driven by Egr-1 promoter in C666-1 cells
We firstly constructed the pEgr-C27-GFP plasmid, in which a cDNA-encoding hTERTC27-GFP fusion protein was put at downstream of Egr-1 promoter. A fluorescence microscopic assay showed the hTERTC27-GFP fusion protein was expressed in pEgr-C27-GFP stably transfected C666 cells (Fig. 1A). About 3.12 μg/mL 5-FU obviously increased the expression of hTERTC27-GFP fusion protein assayed by flow cytometry (p<0.01, Fig. 1B). With the increase of 5-FU dosage, hTERTC27-GFP expression was increased up to 7.21-fold (p<0.01, Fig. 1C), which was confirmed by the Western blot analysis (Fig. 1D). The results indicate that 5-FU can increase hTERTC27 expression by activating the Egr-1 promoter.
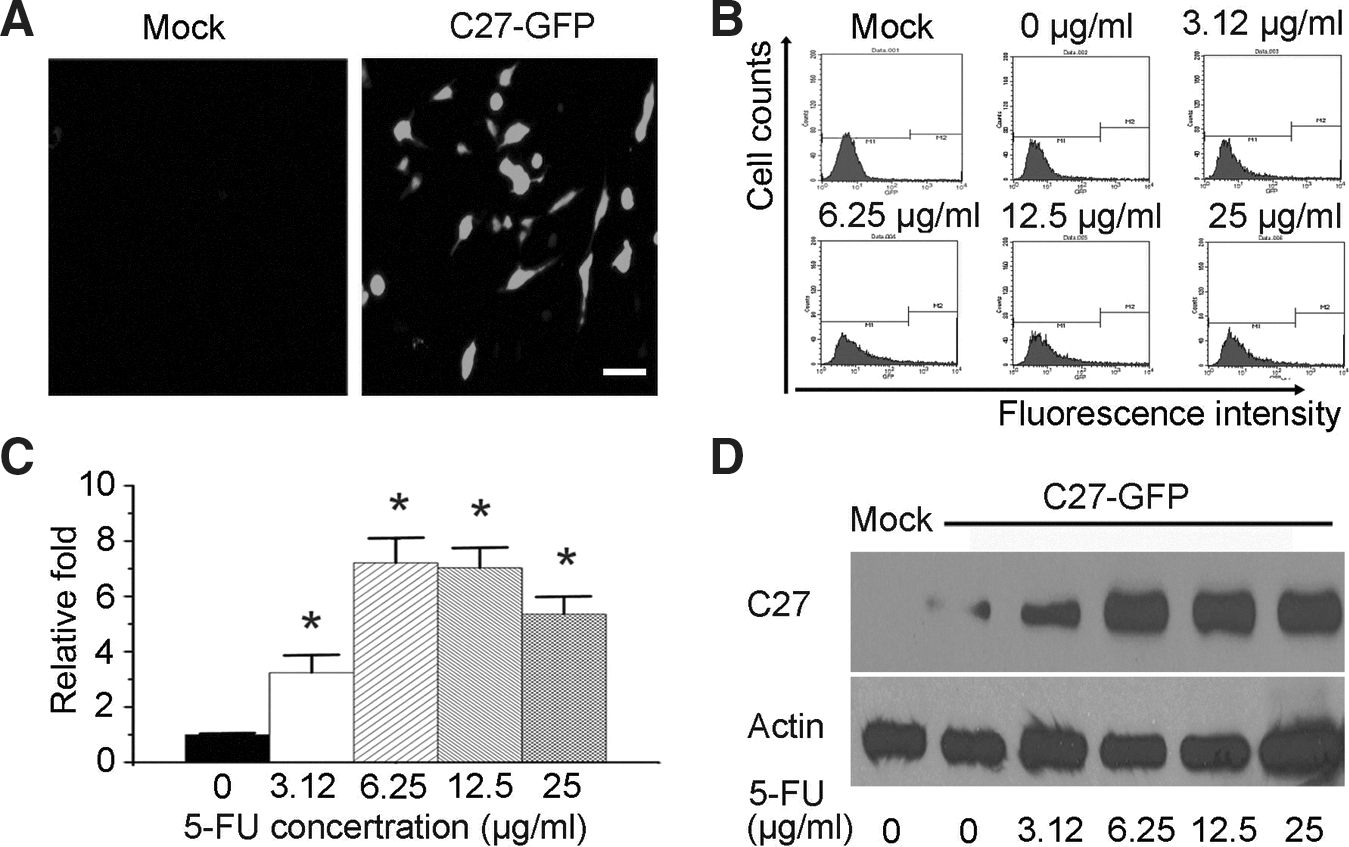
5-FU activated Egr-1 promoter to increase hTERTC27-GFP expression in C666-1 cells.
hTERTC27 increased sensitivity of C666-1 cells to 5-FU
To examine the effect of overexpressed hTERTC27 on the survival of 5-FU-treated C666-1 cells, growth inhibitions of C666-Mock, C666-Con, or C666-C27 cells were analyzed with the MTT assay after treatment with various concentrations of 5-FU for 48 hours. We found that C666-C27 cells showed a lower proliferation ability than C666-Con cells and C666-Mock cells. The IC50 of 5-FU in C666-C27 cells (15.11±0.92 μg/mL) was significantly lower than those in C666-Mock cells (22.88±2.45 μg/mL) and C666-Con cells (21.31±1.91 μg/mL) (p<0.05). IRs of cells were not significantly different between C666-Mock cells and C666-Con cells (Fig. 2). These results indicate that overexpressed hTERTC27 makes the C666-1 cells more sensitive to 5-FU treatment.
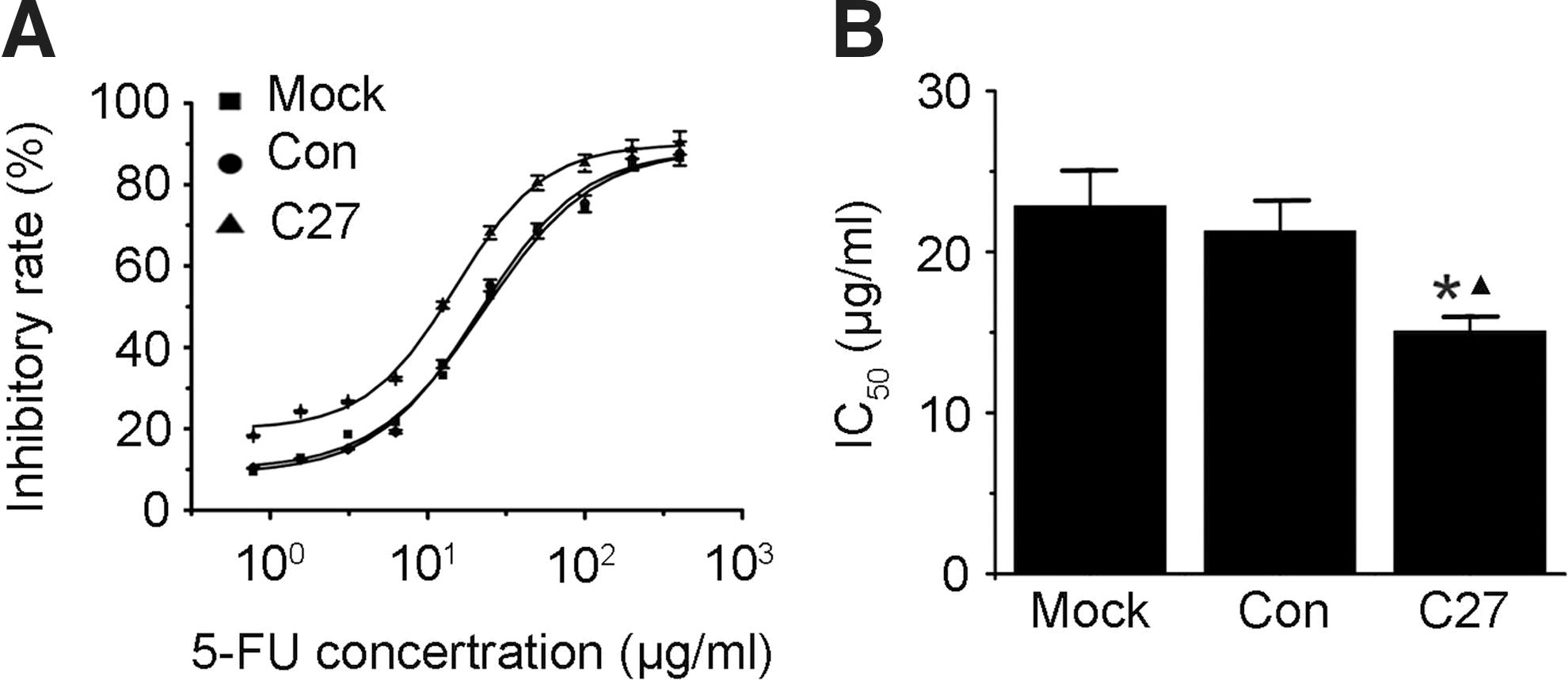
hTERTC27 increased sensitivity of C666-1 cells to 5-FU.
Overexpressed hTERTC27 and 5-FU synergistically inhibited C666-1 cell growth
The synergistic effect of the overexpressed hTERTC27 polypeptide and 5-FU on C666-1 cell proliferation was determined by the MTT assay. As shown in Figure 3A and B, cell proliferation was significantly inhibited in group C27 (p<0.05) and groups 5-FU (p<0.01) when compared with that in group control. Interestingly, groups C27+5-FU demonstrated lower cell proliferation rates than group C27 (p<0.01) and groups 5-FU (p<0.01). Consistently, colony formation assay showed that the colony formation rate in group C27 (71.67±6.50%) was significantly lower than that in group control (95.00%±7.23%) as showed in Figure 3C and D (p<0.05). Moreover, the combined therapeutic group of C27+6.25 μg/mL 5-FU (0.33%±0.10%) demonstrated a further decrease in the colony formation rate when compared with groups C27 (71.67%±6.50%, p<0.01) or 6.25 μg/mL 5-FU (8.33%±2.95%, p<0.01) alone. There was not any colony found in group 12.5 μg/mL 5-FU only and group 12.5 μg/mL 5-FU+C27 (Fig. 3C, D). These results indicate that overexpressed hTERTC27 and 5-FU synergistically inhibited the proliferation of C666-1 cells.
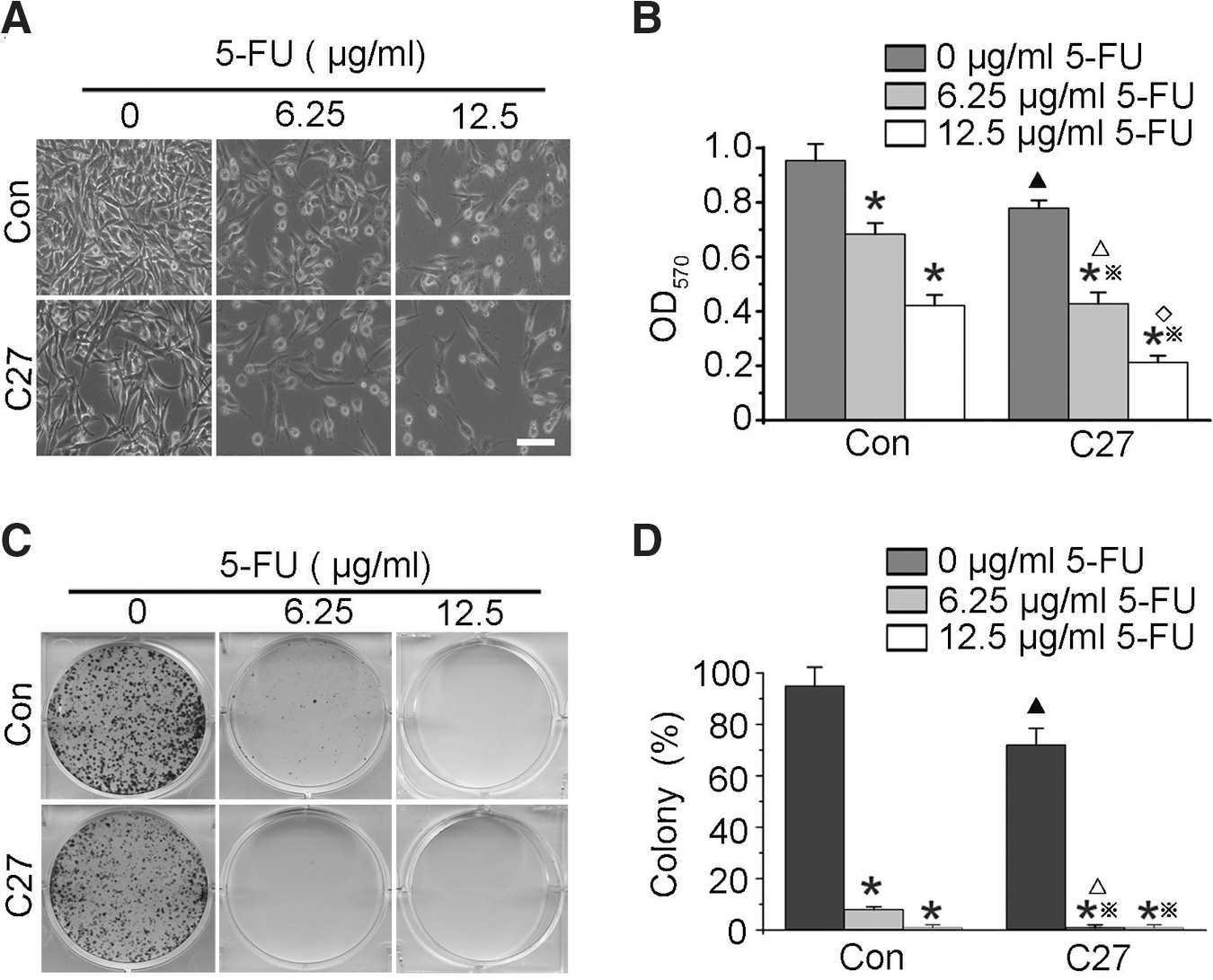
hTERTC27 and 5-FU synergistically inhibited C666-1 cell growth.
Overexpressed hTERTC27 and 5-FU synergistically induce apoptosis in C666-1 cells
To determine if overexpressed hTERTC27 and 5-FU cause apoptosis of C666-1 cells, the early apoptotic death (AnneV+/PI−) was assessed by flow cytometry. As shown in Figure 4A and B, the apoptotic death in group C27 (7.41%±1.81%) was significantly increased compared to that in group control (2.43%±0.37%, p<0.01). Further, group C27+6.25 μg/mL 5-FU (15.77%±3.98%) demonstrated a significant increase in the apoptotic death compared to group C27 (p<0.01) and group 6.25 μg/mL 5-FU (10.34%±2.07%, p<0.05), and this trend of difference was also found between group C27+12.5 μg/mL 5-FU (35.15%±5.66%) and group 12.5 μg/mL 5-FU (26.32%±4.12%). To confirm this result, the TUNEL assay was performed (Fig. 4C, D). It was found that ∼9.11% C666-1 cells were TUNEL positive in group C27, which is significantly higher than control (about 3.27%, p<0.01). However, about 36.73% and 61.65% C666-1 cells were TUNEL positive in groups C27+6.25 μg/mL 5-FU and C27+12.5 μg/mL 5-FU, respectively, whereas only about 24.34% and 44.14% C666-1 cells were under apoptosis in groups 6.25 μg/mL 5-FU and 12.5 μg/mL 5-FU. The apoptotic rates of C666-1 cells in combined therapeutic groups were remarkably higher than groups C27 (p<0.01) and 5-FU (p<0.05). These data suggest that overexpressed hTERTC27 driven by Egr-1 promoter and 5-FU synergistically promotes the C666-1 cell apoptosis.
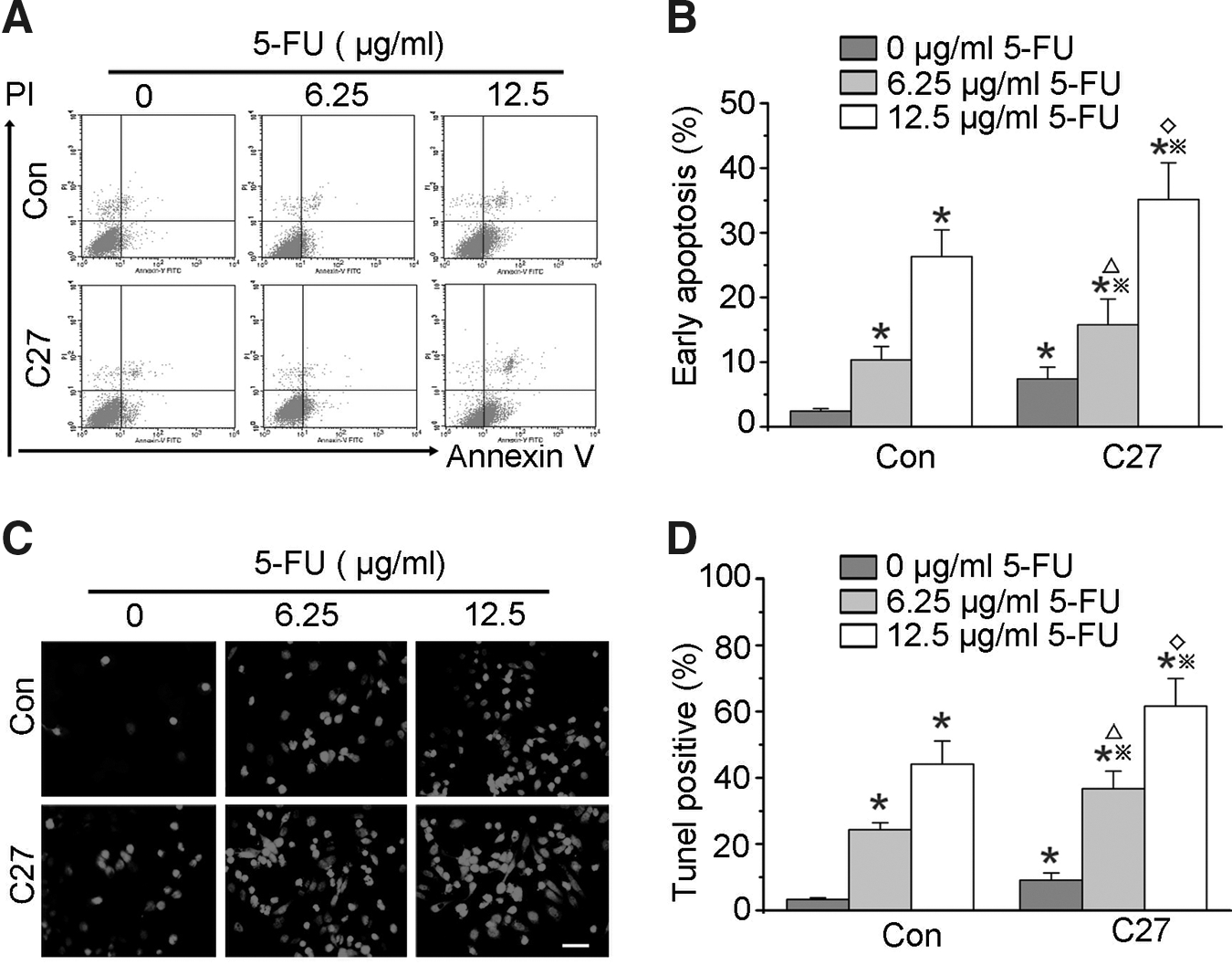
Synergistic effects of hTERTC27 and 5-FU on apoptosis in C666-1 cells.
5-FU-induced hTERTC27 polypeptide and 5-FU synergistically inhibited the growth of NPC tumor in vivo
BALB/C mice were injected subcutaneously with C666-Con cells or C666-C27 cells, respectively. A week after injection, animals were treated with 40 mg/kg 5-FU daily, and tumor growth was monitored for 5 days. As shown in Figure 5A, tumor growth was significantly inhibited by the hTERTC27 polypeptide and 5-FU. The tumor volume and weight in groups C27 and 5-FU were significantly diminished compared with those in group control (Fig. 5A–C, p<0.05). Furthermore, the volume and weight of tumor in group C27+5-FU were significantly lower than in groups C27 and 5-FU (Fig. 5B, C, p<0.05). In addition, local muscle infiltration was found in all mice of group control, whereas local muscle infiltration was found in only 67.00% of group C27 mice, 50.00% of group 5-FU mice, and 16.70% of group C27+5-FU mice, respectively (Fig. 5D).
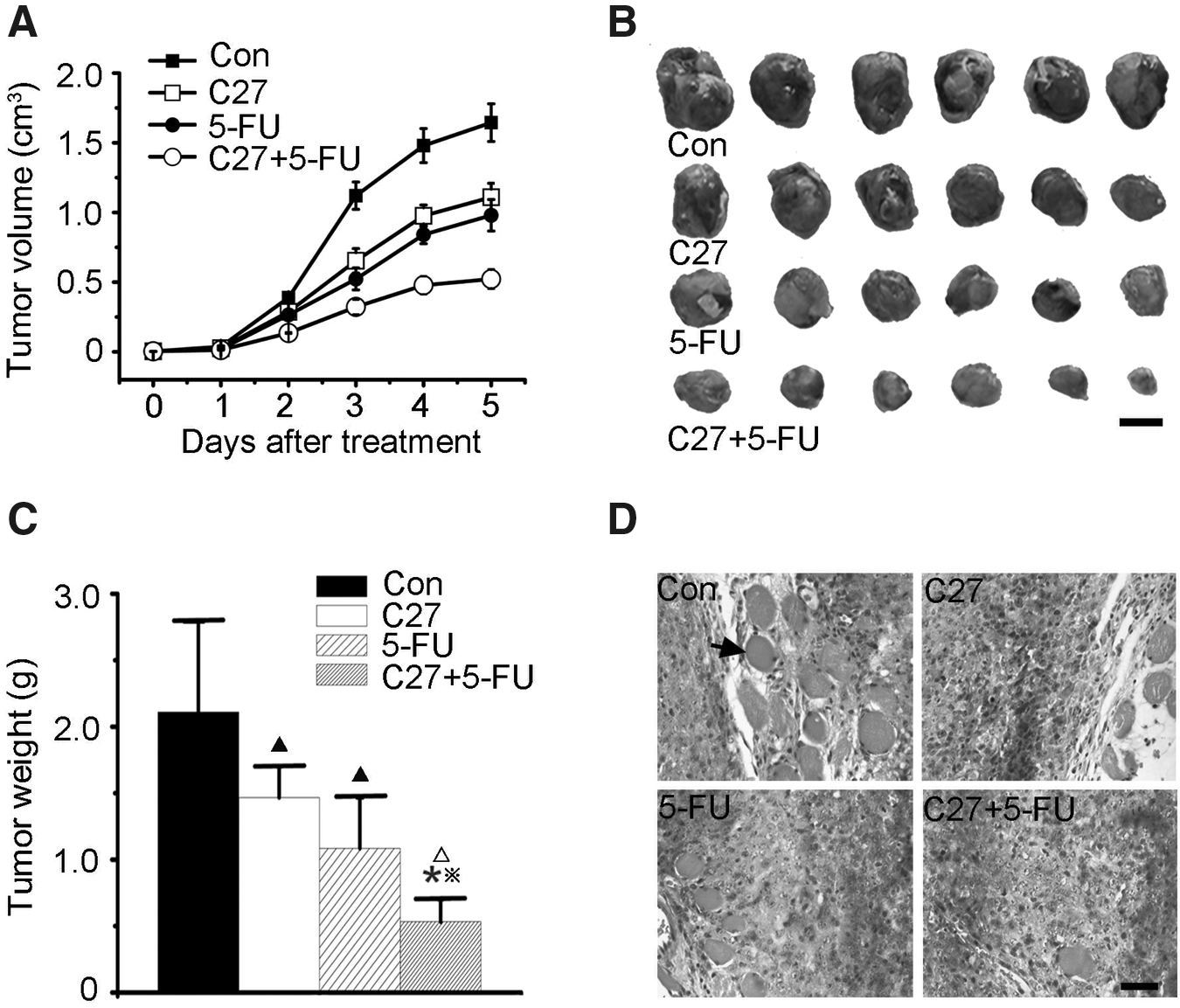
In vivo synergistic antitumor effect of hTERTC27 and 5-FU on nasopharyngeal carcinoma tumor.
5-FU-induced hTERTC27 polypeptide and 5-FU promote the cleavage of poly(ADP-ribose) polymerase, the activation of caspases, and down regulation of Bcl-2
It is well known that apoptosis is characterized by the cleavage of poly(ADP-ribose) polymerase (PARP) and activation of caspases. Thus, we examined the effect of hTERTC27 and 5-FU on the cleavage of PARP and the activation of caspases. As shown in Figure 6A and B, the amount of PARP cleavage was markedly enhanced in group C27+5-FU when compared to that in group 5-FU. Similar results were also observed in the activation of caspases (Fig. 6A, B). These results consistently showed that the combination of hTERTC27 and 5-FU synergistically induced apoptotic death in NPC tumors. Then, we evaluated changes in the signaling proteins of the mitochondrion-dependent apoptotic pathway. Bcl-2 is a very important protein of the antiapoptotic family. As shown in Figure 6C and D, the Bcl-2 expression in groups C27 and 5-FU remarkably decreased in comparison with that in group control. Further, the Bcl-2 expression in group C27+5-FU diminished obviously when compared with that in groups C27 or 5-FU.
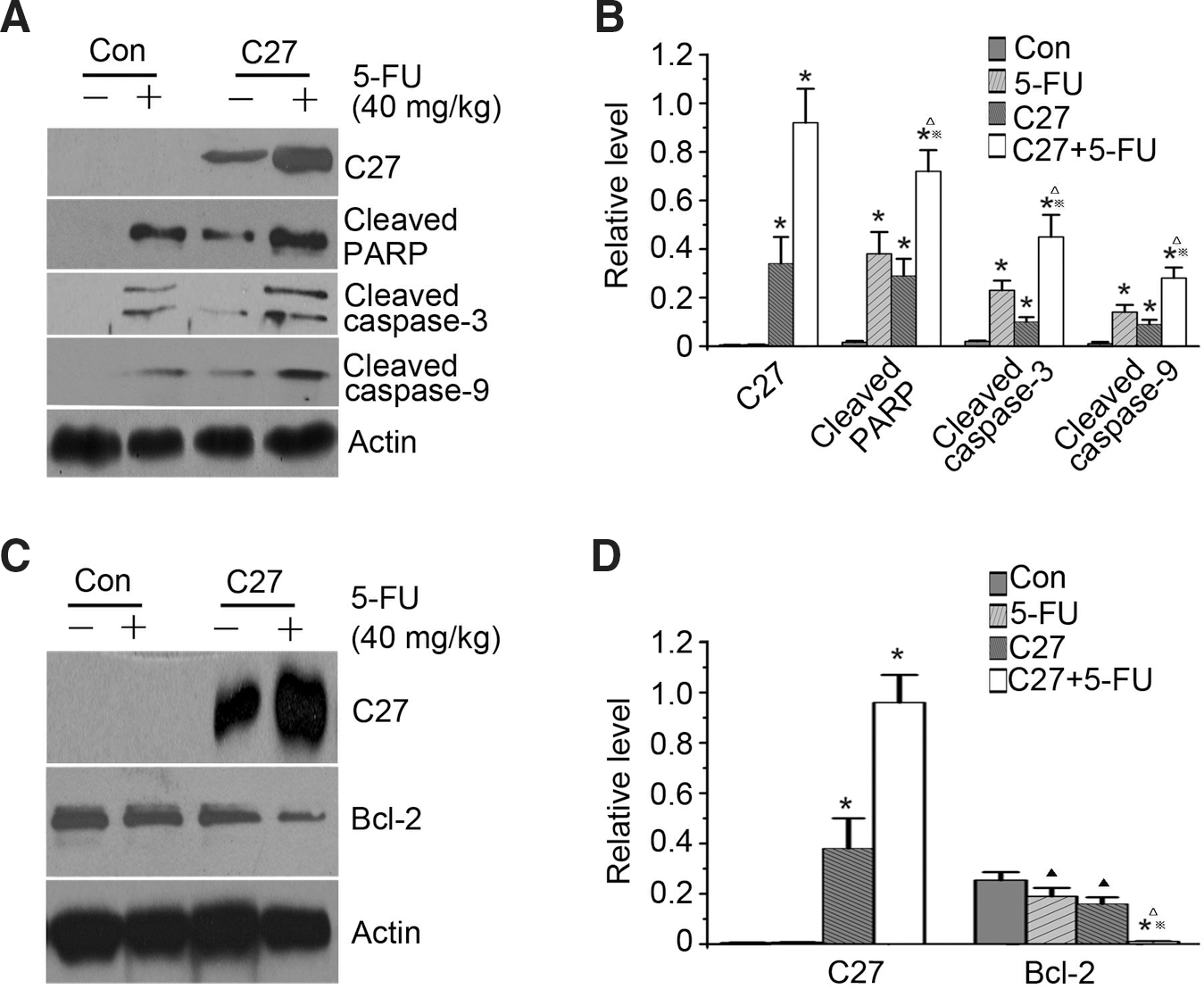
Immunoblotting analysis of cleaved PARP, caspases, and Bcl-2 protein.
Discussion
The motivation of this study was to determine if the Egr-1 promoter can be activated by 5-FU to increase hTERTC27 expression, and if 5-FU can be used as an inducer to increase Egr-promoter-driven hTERTC27 polypeptide overexpression and a chemotherapeutic agent on NPC cancer. Our results showed the Egr-1 promoter, which is sensitive to reactive oxygen intermediates, 18 was activated by 5-FU to drive hTERTC27 overexpression. At appropriate concentrations of 5-FU, we observed the synergistic antitumor effect of the hTERTC27 polypeptide regulated by the Egr-1 promoter and 5-FU on nasopharyngeal carcinoma.
As we know, 5-FU is one of the most widely used drugs in NPC chemotherapy 1,2 because it can effectively inhibit the activity of thymidylate synthase, block the DNA synthesis, arrest the cell cycle, and promote apoptosis. However, the severe side effects of 5-FU and its limited sensitivity to cancer cells still bring great obstacles to cancer therapy. Different strategies have been designed to decrease the dosage and increase the antitumor sensitivity of 5-FU. Therefore, it is very important to combine 5-FU chemotherapy and gene therapy to make the tumor cells sensitive to 5-FU treatment and enhance its antitumor effects. 19 –21
Recently, a variety of therapeutic approaches are being investigated to target telomere or telomerase, since many tumors express high levels of telomerase. 6,22 –24 We have newly constructed the hTERTC27 polypeptide and previously reported that this polypeptide rendered the HeLa cells more sensitive to H2O2 treatment. Moreover, our previous study showed that overexpressed hTERTC27 induced telomere dysfunction in HeLa cells and inhibits cervical carcinoma and glioblastoma tumor growth and tumorigenicity. 16 –18 However, hTERTC27 gene therapy alone cannot quickly inhibit the tumor growth because of its limited expression level. Here, our results showed that hTERTC27 remarkably inhibited cell proliferation, promoted apoptosis of C666-1 cells in vitro, and obviously slowed down the NPC tumor growth in vivo as well. The IC50 value of 5-FU in hTERTC27-overexpressed NPC cells was significantly lower than that in control cells, which indicated that a low concentration of 5-FU treatment in combination with hTERTC27 polypeptide achieves the same antitumor effect as a higher 5-FU concentration. This strategy can decrease the therapeutic dosage and the side effects of 5-FU.
To further optimize the therapeutic strategy of the hTERTC27 gene therapy and 5-FU chemotherapy, we introduced the Egr-1 promoter as a bridge to amplify the antitumor effect of hTERTC27 and 5-FU. The Egr-1 promoter increased hTERTC27 expression in NPC cells due to reactive oxygen intermediates caused by 5-FU, and the high expression of hTERTC27 rendered the cells more sensitive to 5-FU and made the cells generate more reactive oxygen intermediates and further led to a higher hTERTC27 protein level. By recycling, the combined therapy of hTERTC27 driven by Egr-1 promoter and 5-FU exerted a more efficient antitumor effect just as found in our study. Also, the combined treatment of hTERTC27 and 5-FU significantly promoted apoptosis in C666-1 cells. An in vivo mouse model also showed that hTERTC27 driven by the Egr-1 promoter and 5-FU synergistically inhibited tumor growth and local infiltration.
The Bcl-2 family proteins have been shown to play important roles in apoptotic cell death. 25 Bcl-2 inhibits cell apoptosis. Here, we found that Bcl-2 expression was obviously diminished in tumors of combined treating group. Caspase-3 and caspase-9 are involved in cell apoptosis. A classical substrate of activated caspases is PARP, which is a DNA-repair enzyme needed for normal cells. 26 Interestingly, we found that obviously elevated activated caspase-3 and 9 expressions and cleaved PARP with combined therapy of C27 and 5-FU. These results confirmed the synergistic antitumor effects of the hTERTC27 polypeptide driven by Egr-1 promoter and 5-FU on nasopharyngeal carcinoma.
In summary, 5-FU activated the Egr-1 promoter to overexpress hTERTC27 polypeptide, which enhanced the sensitivity of NPC cells to 5-FU treatment. Our study proposes that the combinational therapy of 5-FU-Egr-1-hTERTC27 is adaptable to treating nasopharyngeal cancer.
Footnotes
Acknowledgments
This work was supported by the grants from Shenzhen/Hong Kong Innovation Circle (CXQ2008043), National Natural Science Foundation of China (NSFC) (30870921, 81170632, and 30900335), and Doctoral Training Fund 20100001110047, International Science & Technology Cooperation Program of China 2012DFA11070.
Disclosure Statement
The authors declare no conflict of interest.