
Abstract
Background:
Pancreatic cancer has poor prognosis and high mortality. Currently, the therapy of pancreatic cancer remains a challenge. In this study, we compared the antitumor activity of the recombinant antitumor antiviral protein (RAAP), an improved interferon, with gemcitabine, a classic chemotherapy agent used for pancreatic cancer treatment.
Methods:
The proliferation of Bx-PC3 pancreatic cancer cells was evaluated by an MTT assay. Cell cycle arrest and apoptosis were evaluated by flow cytometry and annexin V-FITC/propidium iodide double staining, respectively. The expressions of matrix metalloproteinase (MMP)-2, MMP-9, caspase-3, caspase-8, and caspase-9 genes were evaluated by reverse transcription–polymerase chain reaction and the Western blot analysis. A xenograft pancreatic cancer model was established by inoculating Bx-PC3 cells into athymic nude mice. The antitumor activity of RAAP and gemcitabine was tested in the xenograft tumor model.
Results:
RAAP significantly inhibited proliferation, induced cell cycle arrest, and induced apoptosis in Bx-PC3 cells in vitro and delayed tumor growth in vivo. The antitumor activity of 20 ng/mL of RAAP was a little more effective than 10 μM of gemcitabine. The antitumor activity of RAAP was associated with its role in inducing caspase-3 and caspase-8 expression as well as downregulating MMP-2 expression.
Conclusions:
RAAP can effectively suppress human pancreatic cancer cell growth in vitro and in vivo. The antitumor efficacy of RAAP is similar to gemcitabine.
Introduction
Patients with pancreatic cancer have very poor prognosis with a median survival of less than 6 months. 1,2 A report from the American Cancer Society revealed that pancreatic cancer is the fourth leading cause of cancer-related deaths in the United States. Approximately 43,100 patients were diagnosed with pancreatic cancer, and 36,800 patients died in the United States in 2010. 3 A major reason for high mortality is that most pancreatic cancers are discovered at an unresectable stage. Only 15%–20% of patients have a resectable tumor. For late-stage patients, palliative chemotherapy is the only treatment available. 4 Interferons (IFN) have also been used for pancreatic cancer therapy. 5 However, the use of IFN alone provided little benefit to the patient's survival. A combined administration of IFN with targeting of XIAP, 6 histone deacetylase inhibitors, 5 or recombinant poxvirus vaccines 7 have been tested and were found to be more effective than IFN alone.
Gemcitabine is a nucleoside analog that has been used for first-line chemotherapy of advanced pancreatic cancer for the past decade. 8,9 Gemcitabine arrests tumor growth through inducing tumor cell apoptosis. The apoptotic role of gemcitabine is mediated through replacing one of the building blocks of nucleic acid during replication and inactivating ribonucleotide reductase (RNR) irreversibly by binding to the active site of the enzyme. Once RNR is inhibited, the cell cannot produce the deoxyribonucleotides required for DNA replication and repair, leading to cell apoptosis. 10 However, gemcitabine has been known to cause severe side-effects, such as hematological toxicity. 4 Despite the many advances made in diagnosis and therapy of cancers in the past decades, the overall survival of patients with pancreatic cancer has not been improved significantly. 11 Therefore, there is an urgent need for new agents to improve pancreatic cancer therapy.
The recombinant antitumor antiviral protein (RAAP), also named Novaferon, is an improved IFN produced by DNA shuffling technology.
12
RAAP is a new protein drug developed by Genova Biotech, and its antitumor efficacy was established in the pancreas, intestines, liver, lung, breast, ovary, and prostate cancers (
Materials and Methods
Cell culture
Bx-PC3, a human pancreatic cancer cell line, was obtained from the American Type Culture Collection (ATCC) and grown in the RPMI 1640 medium supplemented with L-glutamine, antibiotics, and 10% fetal bovine serum (Gibco Laboratories). Cells were cultured at 37°C in 5% CO2.
Animal
Thirty female athymic nu/nu mice (7 weeks old) were provided by the Animal Center of the Central South University. The animals were housed socially (5 mice per cage) in a room under standard lighting conditions and temperature. Water and food were provided ad libitum. Animal experiment was conducted under an approved protocol from the Central South University and performed in accordance with the animal care guidelines of the Chinese Council.
MTT assay
Bx-PC3 cells (2×103 per well) were seeded in 96-well plates 24 hours before treatment. Cells were then incubated with RAAP (10 and 20 ng/mL), gemcitabine (10 μM), or saline for 6, 12, 24, or 36 hours. Twenty microliters of the MTT solution (5 mg/mL) was added to each well, and cells were incubated for another 4 hours at 37°C. The supernatant was removed gently and 100 μL of DMSO was added, followed by shaking for 10 minutes at room temperature. The absorbance was measured at 590 nm.
Cell cycle analysis by flow cytometry
To determine the effect of RAAP on cell cycle arrest, cells were grown to 50% confluency and then treated with RAAP (10 and 20 ng/mL), gemcitabine (10 μM), or saline for 24 hours. Cells were harvested by digestion with 0.25% trypsin-EDTA, washed with cold phosphate-buffered saline (PBS), and then fixed with 70% ethanol at −20°C overnight. After washing the cells with cold PBS again, cells were resuspended with the propidium iodide (PI) staining solution containing PI (20 μg/mL) and RNase (100 μg/mL), and incubated for 30 minutes at room temperature in the dark. Stained cells were analyzed using fluorescence-activated cell sorting. The number of gated cells in G1, G2/M, or S-phase was presented as a percentage.
Annexin V-FITC/PI double staining
Apoptosis was assessed using the annexin V-FITC/PI double-staining kit, according to the manufacturer's instructions. Briefly, cells were treated with RAAP (10 and 20 ng/mL), gemcitabine (10 μM), or saline for 24 hours. Cells were then harvested and rinsed with cold PBS twice. After resuspending cells with the binding buffer, 5 μL of annexin V and 1 μL of PI working solution (100 μg/mL) were added to each 100 μL solution of cells and incubated for 15 minutes at room temperature in the dark. The stained cells were then analyzed using flow cytometry.
Reverse transcription–polymerase chain reaction
Bx-PC3 cells were treated with RAAP (10 and 20 ng/mL), gemcitabine (10 μM), or saline for 24 hours. Total RNA was extracted using the TRIzol reagent (Invitrogen), according to the manufacturer's instructions. Reverse transcription (RT) was performed using the RevertAid™ First Strand cDNA Synthesis Kit (Fermatas China). GAPDH, matrix metalloproteinase (MMP)-2, MMP-9, caspase-3, caspase-8, and caspase-9 genes were amplified using primers listed in Table 1. The polymerase chain reaction (PCR)-amplified products were visualized on a 1.5% agarose gel containing 0.5 μg/mL of ethidium bromide.
MMP, matrix metalloproteinase.
Western blot
Bx-PC3 cells were treated with RAAP (10 and 20 ng/mL), gemcitabine (10 μM), or saline for 24 hours. Cells were then rinsed with cold PBS twice and harvested in 0.2 mL of the RIPA buffer (10 mM Tris, pH 7.4, 150 mM NaCl, 1% Triton ×100, 1% deoxycholate, 0.1% SDS, and 5 mM EDTA) containing 1× protease inhibitor cocktail (Sigma-Aldrich). To detect protein expression in tumors, tumor tissues were homogenized in 10 volume of the RIPA buffer and then sonicated for 15 seconds. The protein concentration was measured using the BCA Protein Assay Kit (Beyotime). Twenty micrograms of total protein were run on a 15% SDS-PAGE gel and then transferred to PVDF membranes. After blocking with 5% BSA for 2 hours, membranes were then incubated with a primary antibody (anti-MMP-2, anti-MMP-9, anticleaved caspase-3, anti-caspase-8, or anti-VEGF) overnight at 4°C. After washing with PBS, membranes were incubated with a HRP-labeled anti-mouse or anti-rabbit antibody (1:5000 dilution) for 2 hours. Immunoreactive proteins were detected using an enhanced chemiluminescence reagent (Pierce), according to the manufacturer's instructions. For loading control, membranes were stripped and reprobed with beta-actin (1:5000 dilution). The antibodies were purchased from Promega.
Tumor growth inhibition study
Approximately 5×106 Bx-PC3 tumor cells were subcutaneously injected into the right hind limbs of mice. After the tumors grew to 7 mm in diameter, mice were randomly divided into three groups: control, gemcitabine, or RAAP treatment group. Each treatment group consisted of 10 animals. Mice in the control group were given a saline injection. Mice in the gemcitabine group were intratumorally injected with gemcitabine (0.8 mg/kg, every other day). Mice in the RAAP group were intratumorally injected with RAAP (10 μg/kg, every other day). Animals were euthanized 1 day after last injection. Tumor specimens were snap-frozen and kept at −80°C.
Statistical analysis
Data were analyzed using Statistical Package for the Social Science, version 13.0. Differences between groups were detected with the two-tailed Student's t-test. All data are presented as mean±standard error of the mean. A p<0.05 was considered statistically significant.
Results
RAAP inhibited Bx-PC3 proliferation in a time- and dose-depended manner
A previous study demonstrated that 10 μM of gemcitabine can significantly inhibit Bx-PC3 proliferation. 13 In this study, cells were incubated with 10 ng/mL RAAP, 20 ng/mL of RAAP, or gemcitabine (10 μM). Cell proliferation was measured by the MTT assay. Both Gemcitabine and 10 ng/mL of RAAP significantly inhibited cell proliferation starting at 12 hours after treatment. Twenty nanograms per milliliter of RAAP significantly inhibited Bx-PC3 growth in a time-dependent manner and exhibited more significant inhibition on cell proliferation than either 10 ng/mL of RAAP or gemcitabine alone (Fig. 1).

Inhibition of cell proliferation. Gemcitabine significantly reduced the growth rate in Bx-PC3 cells at 12, 24, and 36 hours compared to control. Both concentrations of RAAP reduced the cell growth rate compared to control. *p<0.05, **p<0.01 versus other three groups. High concentration of RAAP was more effective than gemcitabine at 36 hours (# p<0.05). n=5. RAAP, recombinant antitumor antiviral protein.
RAAP reduced the S+G2 percentage in Bx-PC3 cells
Cancer cell proliferation is controlled by the cell cycle, apoptosis, or both. Cells in the S+G2 phase are currently undergoing DNA replication. Cell cycle arrest in treated Bx-PC3 cells was measured by cytometry. The percent of cells in the S+G2 phase in gemcitabine-treated (10 μM) Bx-PC3 cells was 28%, which was significantly lower than in untreated cells (44%, p<0.05) (Fig. 2). After treatment with RAAP (10 or 20 ng/mL) for 24 hours, the percent of cells in the S+G2 phase decreased to 30% and 24%, respectively (Fig. 2, p<0.05). Our data also demonstrated that treatment with RAAP reduced the percent of cells in the G1 phase (proliferative fraction). These results suggested that RAAP induced cell cycle arrest in Bx-PC3 cells.
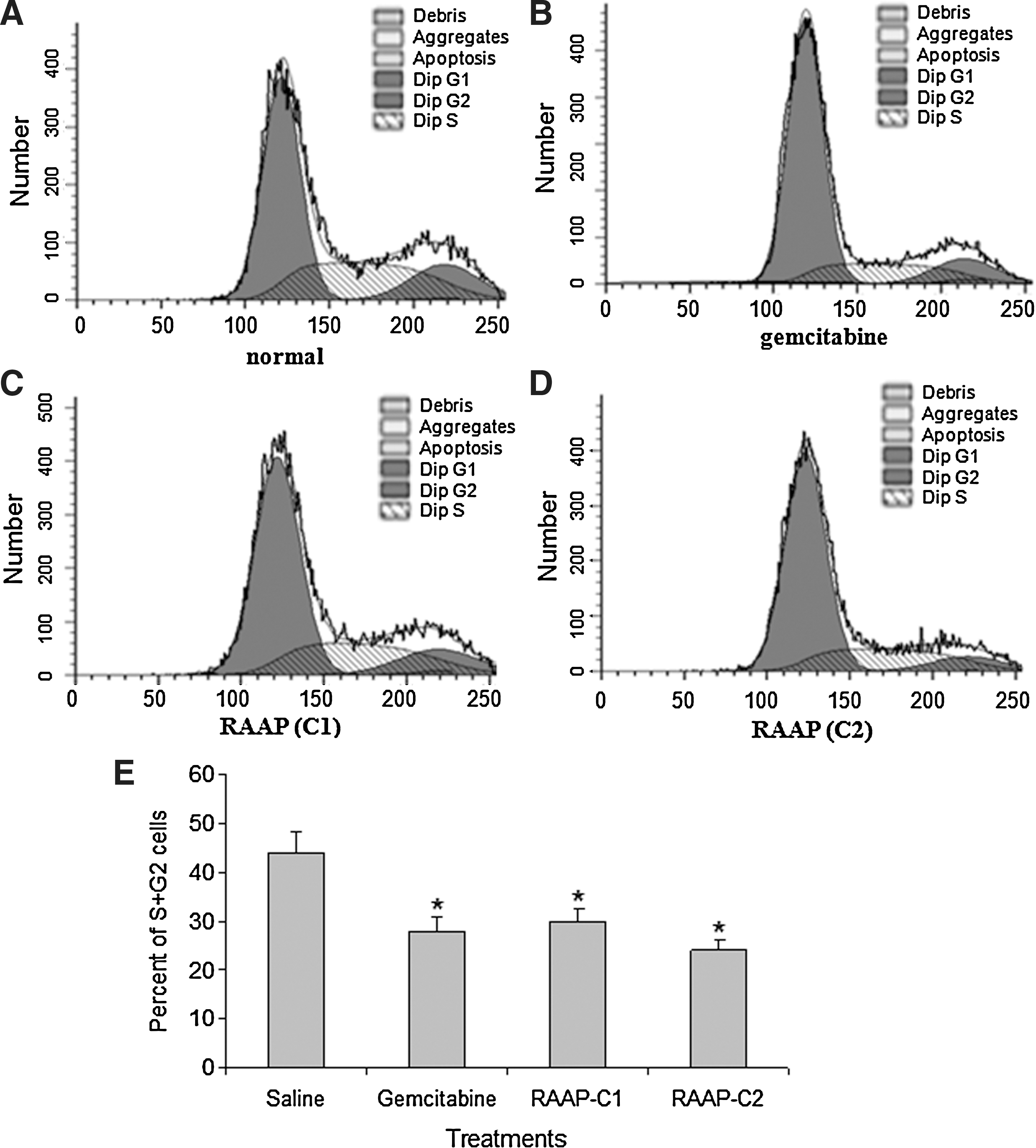
Cell cycle arrest in gemcitabine and RAAP-treated Bx-PC3 cells. Cell cycles were analyzed using flow cytometry after propidium iodide staining.
RAAP induced apoptosis in Bx-PC3 cells via death receptor pathway
Induction of cell apoptosis is another important mechanism to establish effective therapy in cancers. We further tested whether RAAP could induce apoptosis in Bx-PC3 cells. Apoptotic cells were detected by annexin V-FITC/PI double staining. Gemcitabine significantly increased the percent of cells in the R5 stage (early apoptosis) in Bx-PC3 cells (8.12%) compared to the untreated control (6.05%, p<0.05). However, 10 and 20 ng/mL of RAAP induced 10.16% and 15.69% of cells to go into the R5 stage, respectively (Fig. 3, p<0.05). Both gemcitabine and RAAP induced significant increases in the percent of cells in the R3 stage (later stage of apoptosis) (Fig. 3). Twenty nanograms per milliliter of RAAP was more effective than 10 μM of gemcitabine (p<0.05). To investigate apoptotic signaling, the mRNA expression of caspase-3, -8, and -9 was detected by RT-PCR. RAAP significantly increased the expression of caspase-8 and caspase-3 mRNA (Fig. 4A, B, p<0.05), but had no effect on caspase-9 expression (Fig. 4C, p>0.05). These results suggested that RAAP induced apoptosis in Bx-PC3 cells via the death receptor pathway.
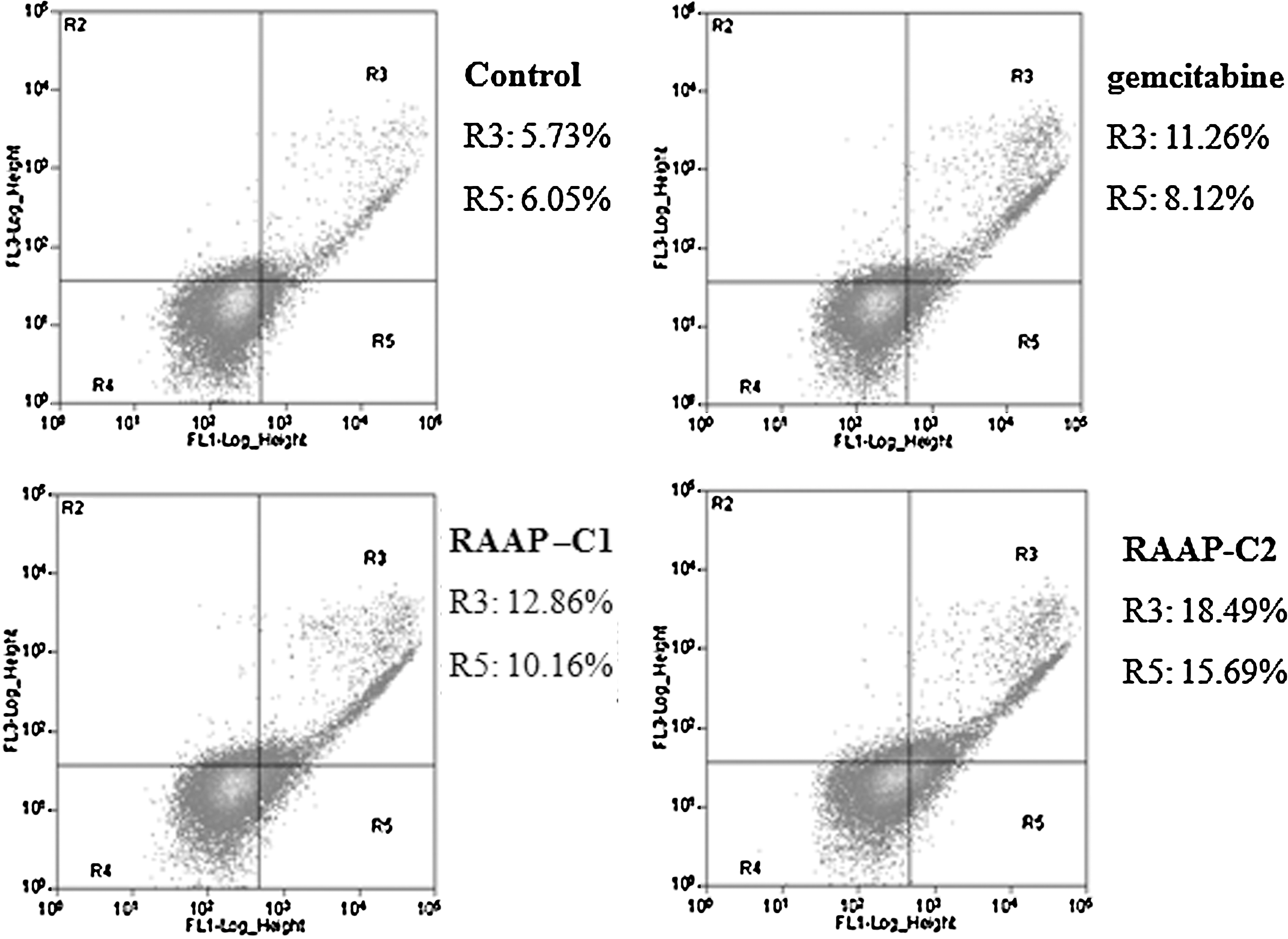
Apoptosis in Bx-PC3 cells. Compared with the control group, gemcitabine and RAAP treatment increased the proportion of early stage apoptosis (R5) and later stage apoptosis (R3). n=5.
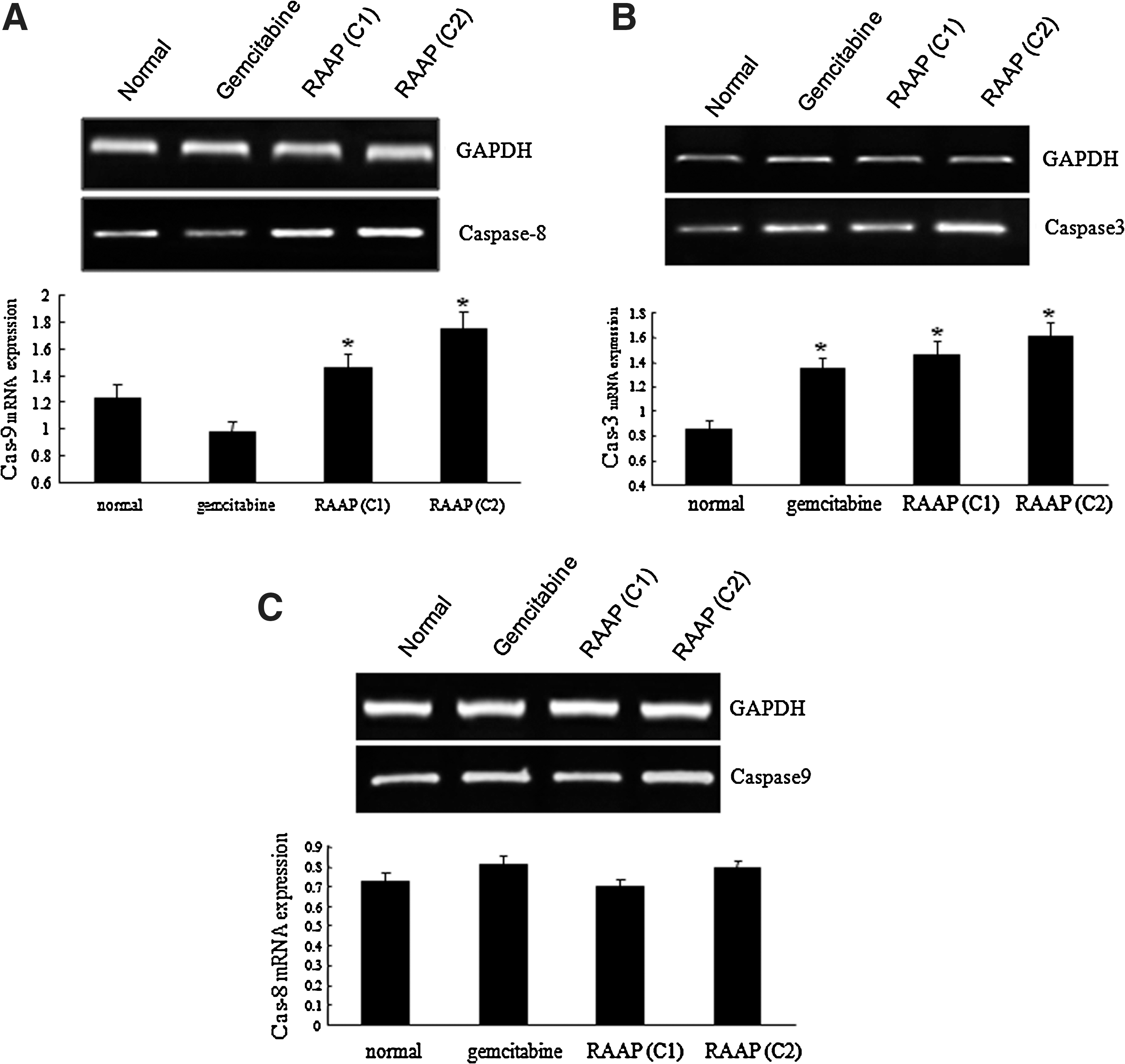
Caspase mRNA expression detected by RT-PCR.
RAAP downregulated the expression of MMP-2 in Bx-PC3 cells
MMPs are involved in the regulation of tumor growth, apoptosis, cell adhesion, and angiogenesis. 14,15 We therefore measured MMP-2 and MMP-9 mRNA and protein expressions in Bx-PC3 cells after treatments. Results showed that RAAP (10 and 20 ng/mL) significantly downregulated MMP-2 mRNA (Fig. 5A, B) and protein (Fig. 5D, E, p<0.05). RAAP stimulated MMP-2 expression in a dose-dependent way. However, RAAP had no significant effect on MMP-9 mRNA (Fig. 5A, C) and protein expression (Fig. 5D, F, p>0.05).
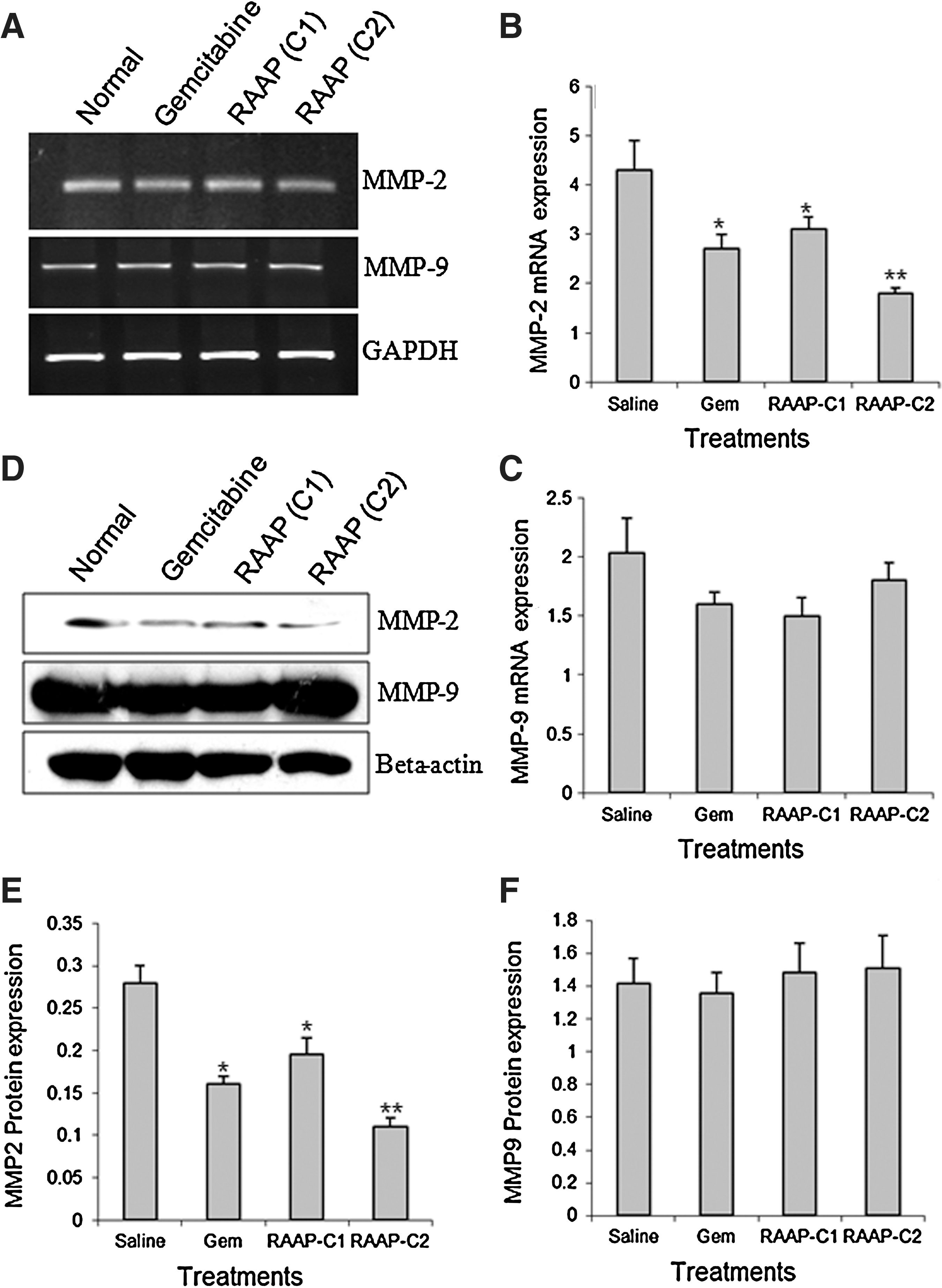
MMP-2 and MMP-9 expression in treated Bx-PC3 cells.
RAAP suppressed tumor growth in xenograft pancreatic tumor model
After Bx-PC3 tumor reached about 7 mm in diameter, mice were given gemcitabine (0.8 mg/kg, every other day), RAAP (10 μg/kg, every other day), and saline injection into tumor for 28 days. Tumors were weighed. As shown in Figure 6A, gemcitabine and RAAP treatments significantly decreased tumor weight (0.163±0.075 g and 0.146±0.093 g, respectively), compared to saline-injected tumor (0.400±0.082 g). We further investigated the protein levels of MMP-2, cleaved caspase-3, caspase-8, and VEGF-A by Western blot. Gemcitabine significantly increased the level of cleaved caspase-3, but had no effect on caspase-8 expression. In contrast, RAAP significantly elevated caspase-8 and cleaved caspase-3 levels (Fig. 6B, C). Both gemcitabine and RAAP significantly decreased the levels of MMP-2 and VEGF-A proteins (Fig. 6C, D).
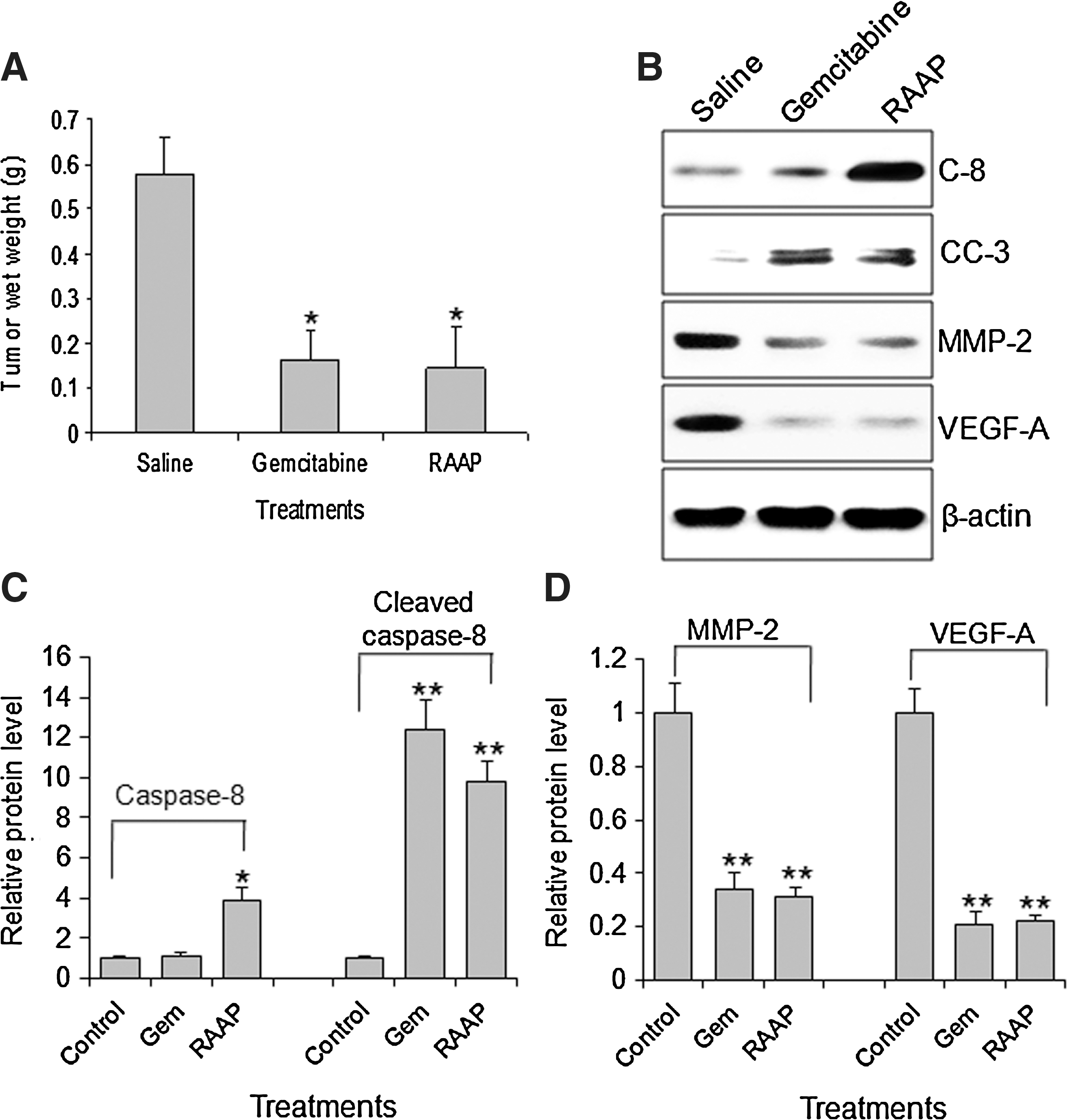
Inhibition of tumor growth in the xenograft tumor model and associated molecular changes. Tumor tissues were harvested 28 days after initial treatments.
Discussion
Gemcitabine is a classic chemotherapy agent and possesses a wide range of antitumor activity. Evidence of gemcitabine inhibiting tumor cell growth was obtained from many in vitro and in vivo studies on a broad range of cancer cells. In the clinic, the antitumor activity of gemcitabine has been extensively demonstrated in a variety of cancers, including pancreatic cancer. 16 –18 In this study, we used gemcitabine as a positive control to evaluate the therapeutic effect of RAAP. We demonstrated that RAAP significantly inhibited Bx-PC3 cell proliferation, induced cell apoptosis, and cell cycle arrest in vitro in a dose-dependent manner and delayed tumor growth in the xenograft tumor model. The activity of 20 ng/mL of RAAP was more efficient than 10 μM gemcitabine. We further demonstrated that the antitumor effect of RAAP was associated with the induction of caspase-3 and caspase-8 expression and inhibition of MMP-2 expression.
Previous studies demonstrated that gemcitabine induces cell death in Bx-PC3 cells with an IC50 of 10 μM. 13 Treatment of pancreatic cancer cells with 10 μM of gemcitabine significantly induced cell cycle arrest and apoptosis at 24 hours. 19 In this study, we confirmed the antitumor activity of 10 μM of gemcitabine in Bx-PC3 cells in vitro and in vivo. Although 10 ng/mL of RAAP also showed a significant anticancer activity, it is less efficient than 10 μM of gemcitabine. In contrast, 20 ng/mL of RAAP exhibited more efficacy than 10 μM of gemcitabine in inducing apoptosis and inhibiting cell proliferation, but showed similar effect in inducing cell cycle arrest and inhibiting tumor growth.
Caspase-3 is one of the most widely studied effector caspases and an activated caspase-3 plays pivotal roles in both death receptor pathways induced by caspase-8, and the mitochondrial pathway involving caspase-9. Interestingly, RAAP significantly increased the expression of caspase-8 and caspase-3 mRNA in vitro, but showed no effect on caspase-9 expression. We further demonstrated that RAAP significantly increased cleaved caspase-3 (activated form) and caspase-8 levels in tumor tissues. These results suggested that RAAP induced apoptosis in Bx-PC3 cells via the death receptor pathway. Previous studies revealed that IFN can activate the death receptor signaling pathway. 20,21 Furthermore, the apoptotic effects of IFNs on pancreatic cancer cells can be blocked by caspase-8 inhibition. 22 Also, IFN gamma induces cell apoptosis in human pancreatic carcinoma cells via the caspase-1-dependent pathway. 23 RAAP is an analog of IFN, but its activity is significantly higher than IFN. It is therefore not surprising that RAAP may inhibit tumor growth in a similar mechanism to the IFNs.
Metastatic tumor dissemination is the main cause of cancer-related death. The interactions between a tumor and the extracellular matrix (ECM) play a critical role in tumor metastasis. MMPs are the main enzymes involved in ECM degradation. While MMPs are mainly considered to be important in tumor invasion, increasing evidence suggests that MMPs are also involved in the regulation of tumor growth, apoptosis, cell adhesion, and angiogenesis. 14,15 Gemcitabine has been revealed to downregulate the expression of MMP-2, 24 and thus inhibiting the invasion of cancer cells. In this study, RT-PCR and Western blot revealed that gemcitabine downregulated MMP-2 expression. Similarly, RAAP inhibited MMP-2 mRNA and protein expression in a dose-dependent manner in vitro. Both gemcitabine and RAAP had no effects on MMP-9 expression. Interestingly, both gemcitabine and RAAP significantly inhibited MMP-2 and VEGF-A expression in tumor tissues, suggesting that downregulation of MMP-2 expression inhibited tumor angiogenesis.
In conclusion, our study suggested that RAAP could effectively inhibit pancreatic cancer cell growth. RAAP's antitumor activity stems from activating the death receptor pathway and upregulating MMP-2 expression. RAAP is more effective than gemcitabine in the antitumor activity in pancreatic cancer cells.
Footnotes
Disclosure Statement
There are no existing financial conflicts.