
Abstract
Introduction
Breast cancer is the most frequently diagnosed cancer and also the leading cause of cancer death among women, accounting for 23% of total cancer cases and 14% of cancer deaths. 1 In China, breast cancer ranks second among all female malignancies in terms of incidence, and ranks between fourth and fifth in terms of mortality. Radiotherapy remains an essential component of the treatment strategy, particularly in the context of breast-conserving therapy for those with early disease. 2 Complications of radiotherapy affecting the surrounding organs can be severe, in particular, epithelial and connective tissue fibrosis, which adversely affect physical and psychological well-being. A radiation sensitizer could, therefore, be of benefit for this group of patients.
In the search for strategies that can improve tumor control while limiting toxicity to normal tissues, the combination of radiation with epigenetic therapies seems an attractive option. Epigenetics is the study of inheritable changes in gene expression or a cellular phenotype caused by mechanisms other than changes in the underlying DNA sequence. 3 Within the mechanisms of relevance to epigenetics, DNA hypermethylation plays a prominent role in the inactivation of genes involved in cell-cycle regulation, DNA repair, apoptosis, cell signaling transcription, and other cellular processes. 4,5 DNA methyltransferase (DNMT) inhibitors can restore the expression of genes silenced by DNA methylation. Preliminary clinical trials of 5-aza-2′-deoxycytidine (DAC), one of the DNMT inhibitors, have revealed antineoplastic activity in patients with leukemia, myelodysplastic syndrome, and solid tumors. 6 –8 The potential to increase the DAC chemosensitivity of cancer cell lines has been proved in vitro. 9 –11 However, there are a few reports on the radiosensitivity associated with exposure to DAC. 12 –15 The present study, therefore, examined the effect of DAC on the radiation sensitivity of breast cancer cells.
Materials and Methods
Cell culture and treatment
Two breast cancer cell lines purchased from the Cell Bank of Type Culture Collection of the Chinese Academy of Sciences were used in the study, MDA-MB-231 and MDA-MB-435. The cells were cultured at 37°C with 5% CO2 and 95% air in RPMI1640 supplemented with 10% (v/v) fetal bovine serum (FBS). The cells were harvested for analysis by trypsinization.
The DNMT inhibitor, DAC, was obtained from Sigma, dissolved in phosphate-buffered saline (PBS) at a stock concentration of 10 mmM, and stored at −20°C. We treated cells at the confluency of 60% using 2.5 μM or 5 μM DAC/PBS in the regular growth medium (RPMI1640 supplemented with 10% (v/v) FBS) for treatment. The incubation time with DAC was 96 hours. Controls were treated with the same amount of PBS. Both drugs and media were refreshed every 24 hours.
Irradiation
Cell monolayers growing in vitro with complete culture medium were irradiated using 6 MV X-rays from linear accelerators (ELEKTA Synergy) with a dose rate of 2 Gy/minute and 1.5 cm bolus as compensators and then incubated at 37°C until they were trypsinized and collected at different time intervals after exposure.
Methylation-specific polymerase chain reaction
DNA was extracted from the cell lines using the TIANamp Genomic DNA Kit (TIANGEN BIOTECH). On bisulfite conversion with the CpGenome DNA Modification Kit (CHEMICON International), the methylation status of ER, PR, HIC-1, BRCA1, 14-3-3σ, and E-cadherin was assessed using a methylation-specific polymerase chain reaction (MSP). Polymerase chain reaction (PCR) products were visualized on a 1.5% or 2% agarose gel with ethidium bromide (Biorad) staining. Primer sequences and annealing temperatures are listed in Table 1. DNA from normal peripheral lymphocytes served as a negative control. The same DNA was treated with M.SssI CpG methylase (New England Biolabs) in the presence of S-adenosylmethionine (New England Biolabs) that served as a positive control. Water was substituted for DNA as a blank control.
Reverse-transcription PCR
Total cellular RNA was isolated from cultured cells with Trizol (Invitrogen Life Technologies). For cDNA synthesis, 2 μg total RNA was used in a 20 μL reaction mixture using the Quantscript RT Kit (TIANGEN BIOTECH). The primers used in the PCR reactions for the genes analyzed are summarized in Table 2. The amplified products were subjected to electrophoresis on a 1% agarose gel containing ethidium bromide. Negative controls for PCR were generated under the same conditions without RNA. The expression of GADPH was used to examine the integrity of RNA in each sample and to standardize the amount of cDNA added to each of the PCR reactions.
Colony-forming assays
For colony-forming assays, cells with 5 μM DAC, 2.5 μM DAC, or without DAC were seeded in six-well tissue culture dishes. The seeded cell number increased with the dose of irradiation: 100 cells for the control (nonirradiated cells), 200 cells for the 2 Gy dose, 600 cells for 4 Gy, and 4,000 cells for 6 Gy, in the MDA-MB-231 cells. For MDA-MB-435 cells, the number seeded was 100 cells for the control (nonirradiated), 200 cells for 2 Gy, 600 cells for 4 Gy, and 2,000 cells for 6 Gy. Cells were exposed to the indicated doses of radiation, and 14 days after radiation treatment, the cells were washed and stained with crystal violet, and colonies containing at least 50 cells were counted. Plating efficiency was calculated by dividing the average number of cell colonies per well by the number of cells plated. Survival fractions (SF) were calculated by normalization to the plating efficiency of appropriate control groups. We used GraphPad Prism (GraphPad Software) to fit the cell survival curve in accordance with a standard linear-quadratic (LQ) model (SF=exp(-αD-βD 2 )), in order to evaluate the radiosensitization of DAC in the two breast cancer cell lines.
MTT analysis
Cells with or without DAC (5 μM) treatment were trypsinized and seeded into 96-well plates at 2000 cells/well with a complete culture medium. After allowing the cells time to attach, the plates were irradiated with a dose of 4 Gy and then incubated at 37°C. At different time intervals (1, 2, 3, 4, and 5 days) after irradiation, the culture medium was removed, and 3-(4,5)–dimethylthiahiazo-(-z-y1)-3,5-di-phenytetrazoliumromide (MTT) dye (5 mg/mL) was added to each well, followed by incubation for a further 4 hours. The reaction was terminated by the addition of dimethyl sulfoxide, and absorbance was measured at a wavelength of 490 nm in a microplate reader (BioRad Model 680). Experiments were repeated thrice.
Cell-cycle analysis
The cell-cycle phase distribution was evaluated using flow cytometry. After incubation with DAC (5 μM) or PBS (control treatment) for 96 hours, cells (1×106 cells/well in six-well plates) were irradiated at a dose of 4 Gy, and after 24 hours, the cells were harvested, fixed in cold 70% ethanol, and incubated overnight at 4°C. The fixed cells were then resuspended in 100 μL and stained with propidium iodide (50 μg/mL) after treatment with 100 μL of Ribonuclease A (RNase A) (10 mg/mL). The stained cells were analyzed for DNA content using an FACScan (Becton Dickinson).
Γ-H2AX formation assay
Immunofluorescence was used to detect γ-histone H2A (γ-H2AX) foci formation. Briefly, the cells (1×103 cells/cover slip) on cover slips in six-well plates with a complete culture medium were irradiated at a dose of 4 Gy. After 1 hour, 6 hours, 12 hours, and 24 hours, respectively, the cells were washed with PBS for 5 minutes and then fixed with 4% paraformaldehyde for 10 minutes. After washing with PBS, the cells were incubated in 0.1% (v/v) Triton X-100 for 10 minutes and then washed with PBS twice for 5 minutes each time. Blocking buffer (5% bovine serum albumin, BSA) was then applied for 1 hour. Primary antibody for γ-H2AX (Upstate/Millipore) was added to label the cells for 2 hours, which were then washed with PBS thrice, for 10 minutes on each occasion. A secondary antibody (FITC-conjugated anti-mouse immunoglobulin G [IgG] Jackson) was added for 1 hour, and the cells were then washed thrice with PBS. Finally, we mounted the slides by applying fluorescent mounting medium with 4, 6-diamino-2-phenylindole (DAPI) (ZSGB-BIO) and cover slips.
Statistical analysis
Statistical analysis was performed using SPSS 11.5 software for Windows (SPSS). All analyses used two-sided hypothesis tests. Differences between experimental groups were determined using the one-way analysis of variance (ANOVA). Probability values of p<0.05 were regarded as statistically significant. All statistical tests were two sided.
Results
Effect of DAC on DNA methylation and gene expression
The breast cancer cell lines were evaluated for the effect of DAC on radiosensitivity. As an indicator of demethylation, initial experiments used MSP analysis to determine the effects of DAC on the DNA methylation status of three genes, ER, PR, and HIC-1, which have been frequently reported to be hypermethylated in these cell lines. 16 As can be seen in Figure 1, the promoter regions of ER, PR, and HIC-1 were heavily methylated in both MDA-MB-231 cells and MDA-MB-435 cells before DAC treatment, whereas DAC treatment resulted in an increase in the level of unmethylated ER, PR, and HIC-1 in both cell lines. Consistent with the changes in methylation, DAC exposure increased the expression of ER, PR, and HIC-1 in both cell lines (Fig. 2). These results indicate that DAC induces the demethylation and reactivation of silenced genes in both the breast cancer cell lines evaluated.
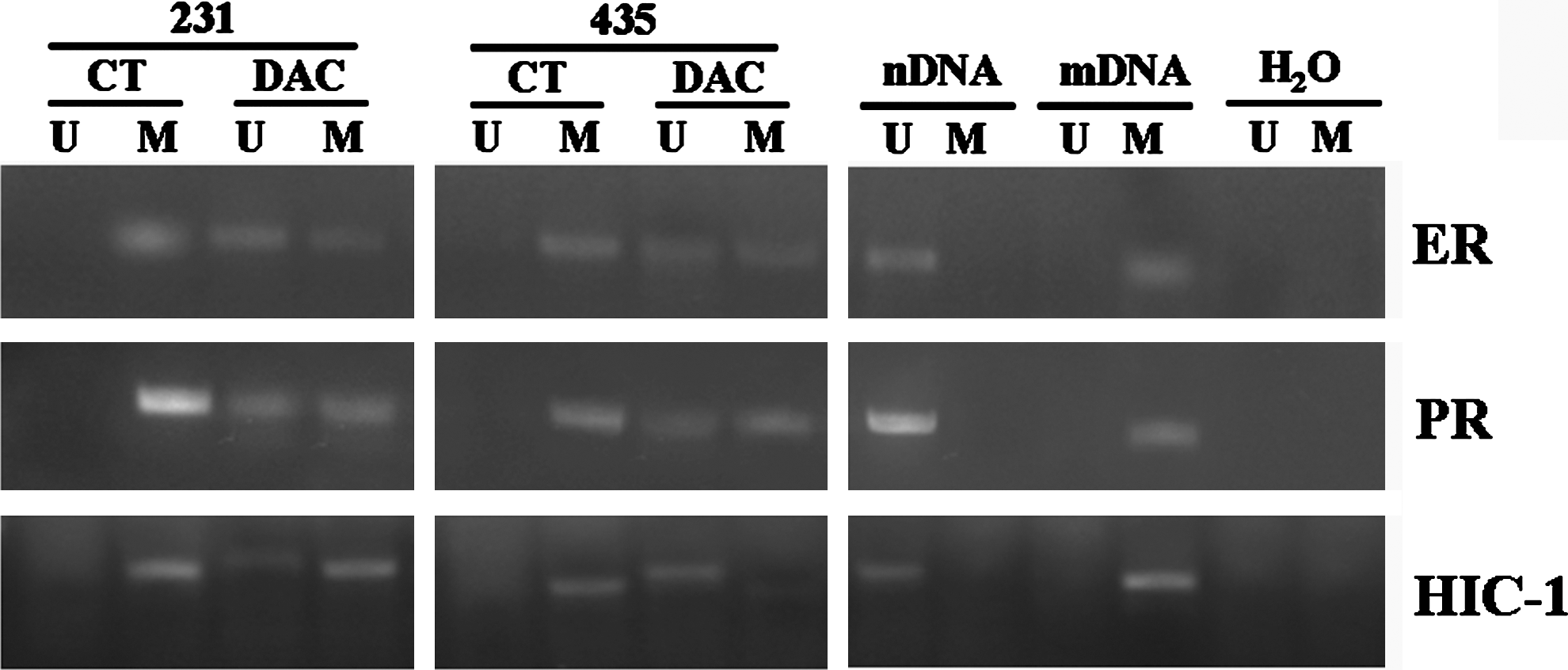
DNA methylation status of ER, PR, and HIC-1 after treatment with DAC: The promoter regions of ER, PR, and HIC-1 were heavily methylated in both MDA-MB-231 cells and MDA-MB-435 cells before DAC treatment, whereas DAC treatment resulted in an increase in the level of unmethylated ER, PR, and HIC-1 in both cell lines. 231: MDA-MB-231; 435: MDA-MB-435. CT: untreatment group; DAC: DAC treatment. U: unmethylated DNA; M: methylated DNA. nDNA: negative control; mDNA: positive control; H20: blank control. DAC, 5-aza-2′-deoxycytidine.
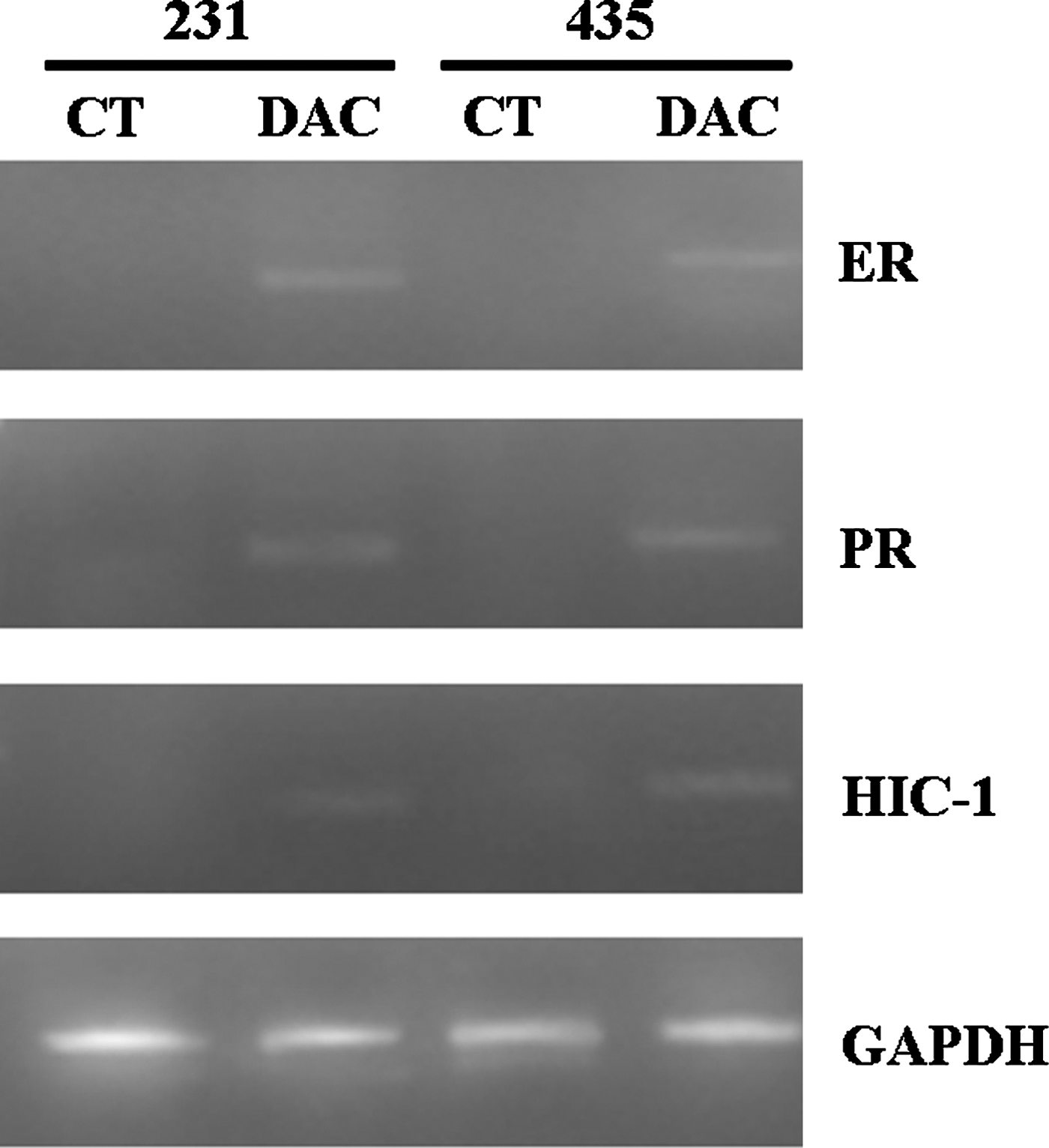
mRNA expression of ER, PR, and HIC-1 after treatment with DAC: The mRNA expression of ER, PR, and HIC-1 was increased by DAC treatment in both MDA-MB-231 cells and MDA-MB-435 cells. 231: MDA-MB-231; 435: MDA-MB-435. CT: untreatment group; DAC: DAC treatment.
DAC enhances radiosensitivity of breast cancer cells
The radiosensitivity of MDA-MB-231 and MDA-MB-435 cells in the 5 μM DAC group, the 2.5 μM DAC group, and the group without DAC was assessed using a colony-forming assay. In the MDA-MB-231 cell line, the SF in the 5 μM DAC treatment group was lowest with 0.398+0.026 for 2 Gy, 0.076+0.003 for 4 Gy, and 0.015+0.002 for 6 Gy, while the SF in the 2.5 μM DAC treatment group was lower than that observed in the DAC-free treatment group, but higher than that of the 5 μM group, with an SF of 0.409+0.021, 0.104+0.010, and 0.033+0.002 for 2 Gy, 4 Gy, and 6 Gy in the 2.5 μM group, respectively, and 0.531+0.022, 0.220+0.005, and 0.057+0.001 in the DAC-free group, respectively. There was a statistically significant difference in the SF between all groups (p<0.01). As shown in the cell survival curves fitted by an LQ model, the SF decreased most rapidly with increases in the irradiation dose in the 5 μM DAC treatment group. Most values of the LQ model parameter (β, α/β, and SF2) in the 5 μM group were statistically significantly different from those in the 2.5 μM group or the control group (p<0.05), indicating that DAC at a concentration of 5 μM enhanced MDA-MB-231 cellular sensitivity to ionizing radiation (Fig. 3 and Table 3).

The cell survival curves fitted by an L-Q linearity Quadri-model using the colony-forming assay in MDA-MB-231: The survival fraction decreased most rapidly with increases in the irradiation dose in the 5 μM DAC treatment group, indicating that 5 μM DAC enhanced MDA-MB-231 cellular sensitivity to ionizing radiation.
p<0.05, versus CT.
p<0.05, versus DAC (2.5 μM).
DAC, 5-aza-2′-deoxycytidine.
In MDA-MB-435 cells, the SF in the 5 μM DAC treatment group remained below that of the 2.5 μM group and the untreated group after irradiation with 2 Gy, 4 Gy, and 6 Gy. There was a statistically significant difference in the SF between all groups (p<0.01). The cell survival curves fitted by an LQ model revealed that the SF decreased most rapidly with increases in irradiation doses in the 5 μM DAC treatment group. The values of β, α/β, and SF2 on exposure to the 5 μM DAC were statistically significantly different from those of the control treatment group (p<0.05), indicating that 5 μM DAC enhanced the radiosensitivity of MDA-MB-435 cells (Fig. 4, Table 4).

The cell survival curves fitted by an L-Q linearity Quadri-model using colony-forming assay in MDA-MB-435: The survival fraction decreased most rapidly with increases in irradiation doses in the 5 μM DAC treatment group, indicating that 5 μM DAC enhanced the radiosensitivity of MDA-MB-435 cells.
p<0.05, versus CT.
The combination of DAC and irradiation induced growth suppression in vitro
We carried out an MTT assay on the two breast cancer cells that had been treated with or without DAC and then irradiated with 4 Gy. The absorbance in the DAC combined with the irradiation treatment groups was significantly lower than that in the DAC treatment group, the irradiation only group, and the control treatment group, in both the MDA-MB-231 and the MDA-MB-435 cell lines within 5 days (p<0.05), indicating that treatment with DAC caused a decrease in cell viability and proliferation after irradiation (data not shown).
DAC induced G2/M cell-cycle arrest after irradiation
We next determined the effect of DAC on cell-cycle phases in the breast cancer cells using Fluorescence Activating Cell Sorter analysis. We found that irradiation alone increased the percentage at G2/M phase, and this was similar to previously published results. 17,18 DAC exposure alone also induced a slight G2/M arrest, but not to the same degree as that observed with irradiation, while the combination of DAC followed by irradiation enhanced G2/M arrest significantly, both in the MDA-MB-231 cells and in the MDA-MB-435 cells (Figs. 5 and 6).
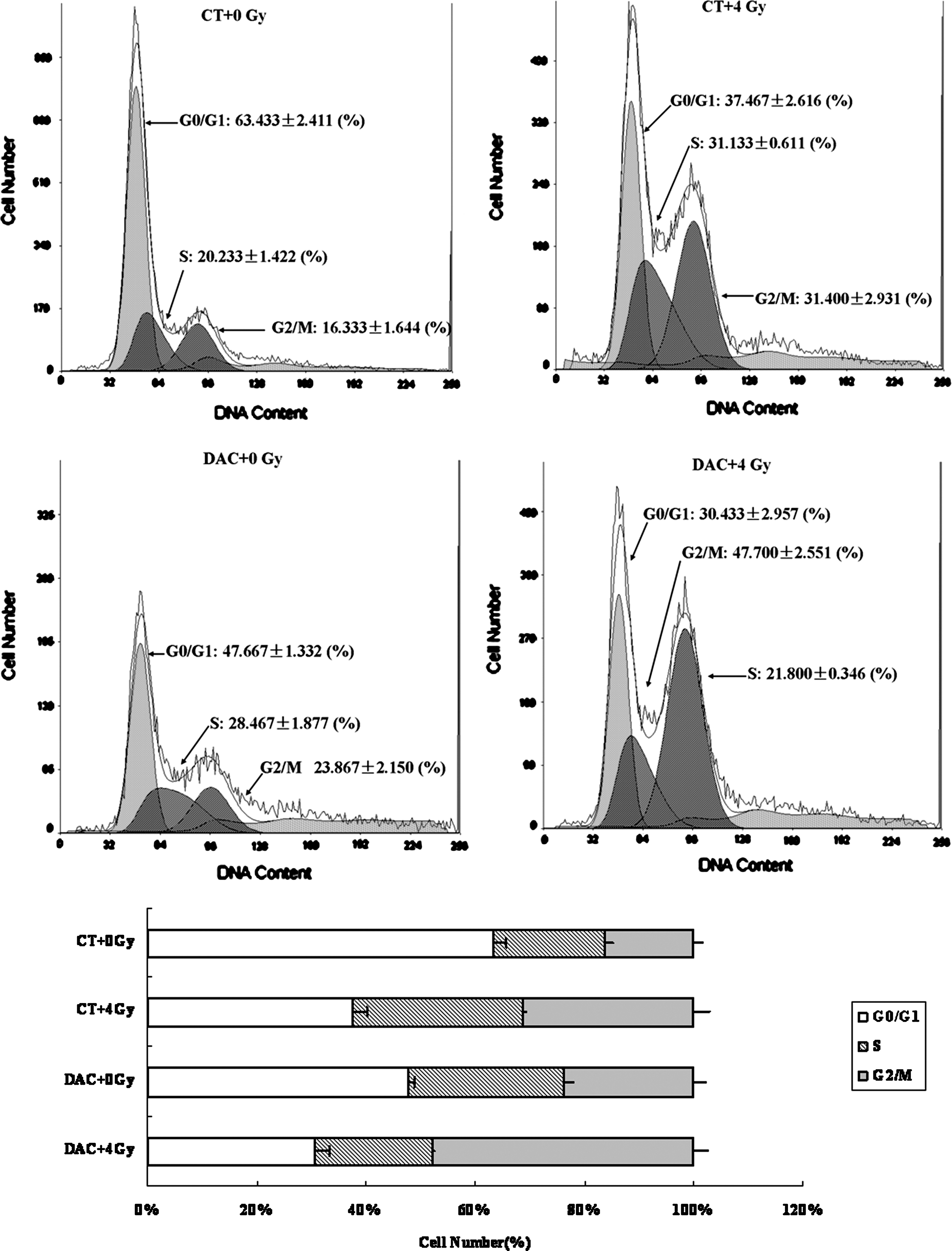
Cell-cycle analysis in MDA-MB-231: Irradiation alone increased the percentage in the G2/M phase. DAC exposures alone also induced a slight G2/M arrest, but not to the same degree as that observed with irradiation, while the combination of DAC followed by irradiation enhanced G2/M arrest significantly in MDA-MB-231 cells.
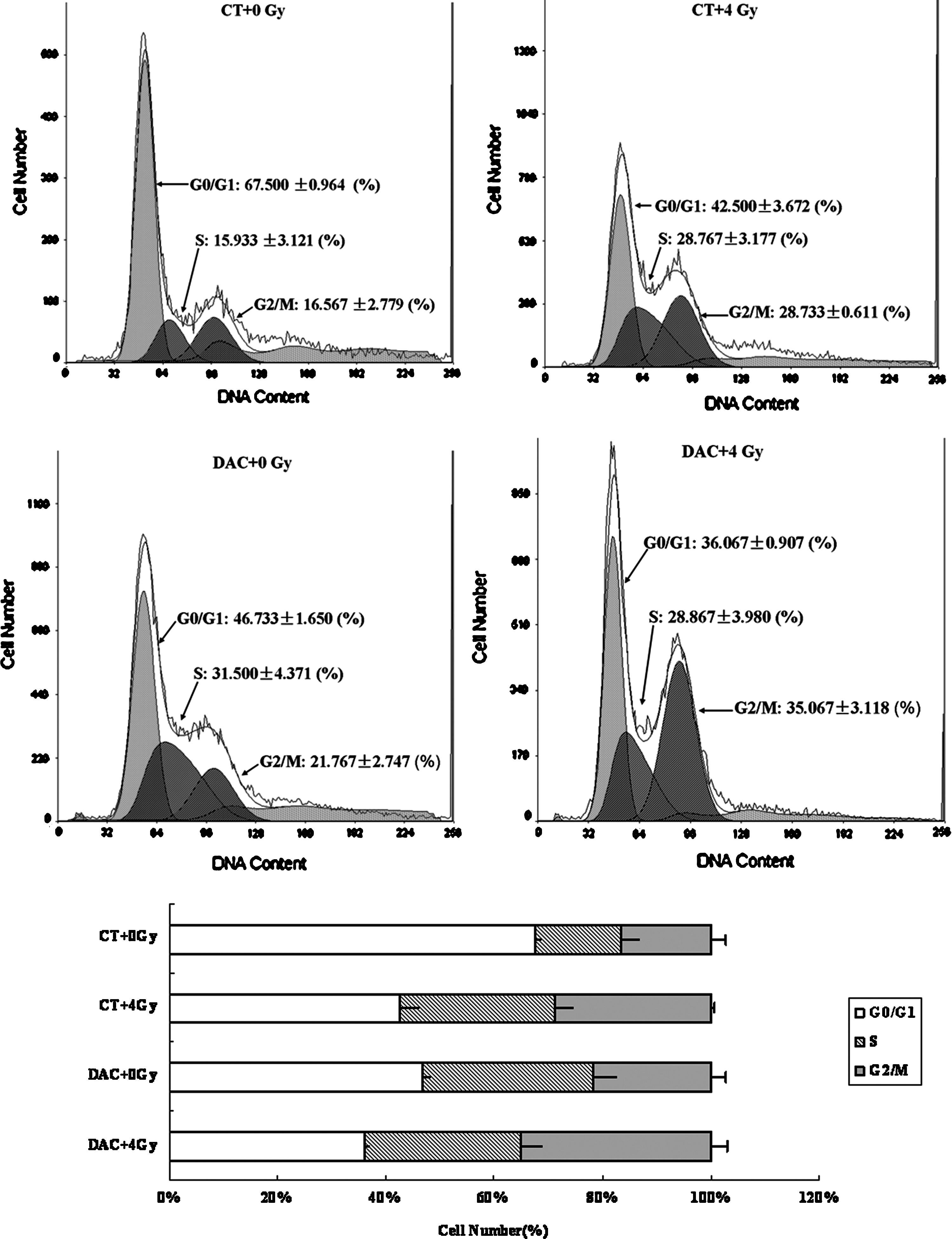
Cell-cycle analysis in MDA-MB-435: Irradiation alone increased the percentage during the G2/M phase. DAC exposures alone induced a slight G2/M arrest, while the combination of DAC followed by irradiation enhanced G2/M arrest significantly in MDA-MB-435 cells.
DAC retardation of DNA repair
Phosphorylation of the histone H2AX is an indicator of DNA double-stranded breaks (DSB). γ-H2AX foci form at a 1:1 ratio to DSB at the site of the broken ends of DNA, making the number of foci a reliable marker for DSB formation, and their subsequent disappearance a measure of repair. The rate of DSB repair predicts the relative sensitivity of a cell to radiation. Here, the repair of DSB produced by irradiation was tracked by counting the number of phosphorylated γ-H2AX foci per cell at several time points after irradiation. In both cell lines, few γ-H2AX foci (less than 3) could be observed in the cells without irradiation, indicating no or rare cells broken. In the irradiation groups in both cell lines, although all cells demonstrated a roughly equivalent number of initial DSBs, the repair of irradiation-induced DSB was impeded in the combined DAC and irradiation group compared with the irradiation alone group. Radiation induced a significant increase in γ-H2AX foci at 1 hour after irradiation in both irradiation groups, which in DAC untreated cells decreased at 6 hours and returned to baseline levels by 24 hours after irradiation. However, for the cells exposed to the combination of DAC and radiation, the number of γ-H2AX foci remained increased, when compared with the radiation-only group, at 6, 12, and 24 hours after irradiation (Figs. 7 and 8).

Analysis of DNA repair on the basis of repair foci after irradiation (MDA-MB-231): Radiation induced a significant increase in γ-H2AX foci at 1 hour after irradiation in both irradiation groups, which in the radiation-only group decreased at 6 hours and returned to baseline levels by 24 hours after irradiation. However, for the cells exposed to the combination of DAC and radiation, the number of γ-H2AX foci remained increased, when compared with the radiation-only group, at 6, 12, and 24 hours after irradiation.

Analysis of DNA repair on the basis of repair foci after irradiation (MDA-MB-435): Repair of irradiation-induced DSB was impeded in the combined DAC and irradiation group compared with the irradiation-alone group. The γ-H2AX foci in the radiation-only group decreased at 6 hours and returned to baseline levels by 24 hours after irradiation. However, for the cells exposed to the combination of DAC and radiation, the number of γ-H2AX foci remained increased, when compared with the radiation-only group, at 6, 12, and 24 hours after irradiation.
Effects of DAC on genes correlated with the radiosensitivity
The methylation status of genes correlated with radiosensitivity (BRCA1, 14-3-3σ, and E-cadherin) in both tumor cell lines, with or without DAC treatment, was determined using the MSP method. In MDA-MB-231 cells, the promoter regions of BRCA1 and 14-3-3σ were unmethylated in the control (untreated) group, and the methylation status was not affected by DAC treatment either for BRCA1 or for 14-3-3σ. However, the E-cadherin gene was completely methylated in the control, and the DAC treatment resulted in an increased level of unmethylation. In the MDA-MB-435 cells, BRCA1 was partially methylated, with 14-3-3σ and E-cadherin completely methylated before DAC treatment, while the methylation status changed as DAC treatment induced complete BRCA1 demethylation, and partial 14-3-3σ and E-cadherin demethylation (Fig. 9).

DNA methylation status of BRCA1, 14-3-3σ, and E-cadherin after treatment with DAC: In MDA-MB-231 cells, the BRCA1 and 14-3-3σ were unmethylated in the control group, and the methylation status was not affected by DAC treatment while the E-cadherin gene was completely methylated in the control and the DAC treatment resulted in an increased level of unmethylation. In the MDA-MB-435 cells, BRCA1 was partially methylated, with 14-3-3σ and E-cadherin being completely methylated before DAC treatment, while DAC treatment induced complete BRCA1 demethylation, and partial 14-3-3σ and E-cadherin demethylation. 231: MDA-MB-231; 435: MDA-MB-435. CT: untreatment group; DAC: DAC treatment. U: unmethylated DNA; M: methylated DNA. nDNA: negative control; mDNA: positive control; H20: blank control.
Discussion
Attempts are being made to enhance the radiosensitivity of breast cancer while protecting normal tissue at the same time. The combination of epigenetic drugs with radiotherapy is particularly interesting in this context, and has demonstrated improved efficacy both in vitro and in vivo in solid tumors other than breast cancer. 12 –15 The data presented here suggest an association between DAC-induced demethylation and radiosensitization in breast cancer cells.
Initially, two breast cancer cell lines were investigated to evaluate the effect of DAC in combination with irradiation. Based on colony-forming assays, which are considered the gold standard for evaluating radiosensitivity, the combination of DAC and radiation yielded a lower SF, and there were significant radiosensitization effects in both the cancer cell lines. In addition, DAC demonstrated synergism of growth inhibition with irradiation, as absorbance in the combined treatment group was less than that observed in any of the other groups. These results suggest that DAC might be a useful radiation sensitizer in breast cancer.
Two doses of DAC (5 μM and 2.5 μM) were used in combination with irradiation in the current study. Yang et al. 19 reported a maximal ER gene re-expression with 2.5 μM DAC for MDA-MB-231 cells and 0.6 μM DAC for MDA-MB-435 cells. Fan et al. 20 obtained different results, with more ER induced with 2.5 μM DAC than with 5 μM, but more growth inhibition was observed with 5 μM. A concentration of DAC 1 μM was used in combination with irradiation by Qiu et al. 13 in gastric cancer cells, and the results demonstrated synergistic effects with the combination. In the current study, 5 μM DAC had a greater radiosensitization effect than 2.5 μM DAC, both in MDA-MB-231 and in MDA-MB-435 cells. Therefore, 5 μM DAC was used in subsequent experiments.
To further explore potential mechanisms, the effect of DAC in combination with irradiation on the cell cycle was initially examined. We found that DAC resulted in an accumulation of radiation-induced G2/M phase arrest in both the cancer cell lines. The activation of the G2 cell-cycle checkpoint, which rapidly arrests cells in G2 allowing time for DNA repair before progression into mitosis, is also considered a determinant of radiosensitivity. Moreover, the G2 checkpoint is considered a target for potential radiosensitizing agents. 21 In our study, since cells in the G2/M phase are the most sensitive to irradiation, the arrest during the G2/M phase caused by DAC might contribute to the enhanced radiosensitivity observed in breast cancer.
A further process involved in determining cellular radiosensitivity is DNA repair, specifically the repair of DNA DSBs. Although these breaks are resealed by complex repair processes, both the rate and extent of DSB repair correlates with cell radiosensitivity, which is to say that impeded DSB repair, as evidenced by the retention of γ-H2AX foci over the first 24 hours after irradiation, correlates with enhanced radiosensitization. 22 During the investigation of the DNA repair processes after irradiation in our study, we found that the repair of radiation-induced DSBs was impeded in the combined DAC and irradiation group compared with the irradiation alone group in both the cell lines.
Although DAC-enhanced radiosensitivity involving the inhibition of DNA damage repair has been reported in many studies, 14,15 the specific process that results in a putative DNA repair defect remains to be determined. However, Dote et al. 14 found that treatment with another DNA methylation inhibitor, zebularine, also enhanced radiosensitivity and involved an inhibition of DNA damage repair. The authors speculate that the radiosensitization induced by zebularine may involve a mechanism similar to that which mediates the sensitization induced by histone deacetylase (HDAC) inhibitors. DNA methylation and histone deacetylation are not independent epigenetic mechanisms. They have a very close relationship and influence each other. 12 Both processes act toward silencing gene expression through the modulation of transcription factor activity and by condensing local chromatin structure. Moreover, DNA methylation acts toward recruiting HDAC to gene promoter regions. HDAC inhibitors enhance tumor cell radiosensitivity associated with a prolongation of γ-H2AX expression and an inhibition of DNA DSBs. Thus, it could be hypothesized that the radiosensitization induced by DAC may involve a similar mechanism to that which underlies the sensitization induced by HDAC inhibitors. Alternatively, since the prolonged expression of radiation-induced γ-H2AX foci may reflect the end result of disparate processes and events leading to the maintenance of unrepaired DSBs, distinctly different mechanisms may be involved. Clearly, additional investigations are required to define the molecular processes responsible for DAC-mediated radiosensitization. However, these results do suggest that enhanced radiosensitivity induced by DAC involves an inhibition of DNA damage repair.
Although DAC enhanced radiosensitivity in both MDA-MB-231 and MDA-MB-435 cells, the degree of radiosensitization varied between the two cell lines. DAC enhanced radiosensitivity to a greater extent in MDA-MB-231 cells than in MDA-MB-435 cells. Thus, we conclude that a different degree of enhanced radiosensitivity induced by DAC may be due to the initially different methylation status in the breast cancer cells. Reversal of the methylation status of genes about radiosensitivity has been found in many other tumor cell types 23,24 although they do not agree with De Schutter's study, 15 which did not find a relationship between drug-induced alterations in epigenetic status and radiosensitization in head-and-neck squamous cell carcinoma cell lines treated with DAC. The authors suggest that this result may indicate that DAC works by mechanisms other than its demethylating activity.
In this study, three genes (BRCA1, 14-3-3σ, and E-cadherin) both of which were related with radiosensitivity and frequent hypermethylation in breast cancer were determined using the MSP method. Among these, BRCA1 is a classic tumor suppressor gene in which mutations are associated with hereditary breast cancers, while promoter methylations seem to be more important in sporadic breast cancer 25 and DAC treatment can result in BRCA1 re-expression and growth inhibition. 25,26 BRCA1 is also related to radioprotection by repairing DNA damage. 27 14-3-3σ has been shown to be associated with growth regulation and signal transduction as well as promoting repair DNA damage caused by irradiation and leading to radioresistance. 28 Hypermethylation of the gene was often observed in breast cancer, and DAC could result in its demethylation as well as the reactivation of gene expression. 29 Loss of E-cadherin expression in breast cancers results from hypermethylation of the gene promoter region. 30 DAC could modify the methylation status in the E-cadherin and induce its re-expression, inhibit tumor growth, and reduce the potential for metastasis. 31 E-cadherin modulates radioresistance with low-cell-surface E-cadherin, which is more resistant to radiation. 32 We found that different methylation patterns were present in the two cell lines, BRCA1 and 14-3-3σ, which were associated with radioprotection, were unmethylated in MDA-MB-231 cells while showing partial and complete methylation in MDA-MB-435 cells. Treatment with DAC had no effect on BRCA1 and 14-3-3σ methylation status in MDA-MB-231 cells, but rather induced demethylation of the two genes in MDA-MB-435 cells. With regard to the E-cadherin gene, which is associated with radiosensitization, there was no difference in either cell line. So, the demethylation of both radioresistant genes (BRCA1 and 14-3-3σ) and radiosensitive gene (E-cadherin) after administering DAC made a less degree of radiosensitization by DAC in MDA-MB-435 cells than in MDA-MB-231 cells. These studies suggest that DAC can enhance radiosensitization in breast cancer cells specifically by demethylating radiosensitizing genes, and that the different radiosensitization effects in different breast cancers cells may be correlated with the primary methylation status of the radiosensitizing genes. Furthermore, the result was that a less significant increase in unrepaired DNA damage in MDA-MB-435 cells than in MDA-MB-231 cells after DAC treatment may partly attribute to the effect of BRCA1 and 14-3-3σ in repairing DNA damage. However, the inapparent difference in the repair of DSB before DAC treatment between the two cell lines also suggests that other genes may participate in the repair of DNA damage induced by irradiation. So, to elucidate the mechanism of DAC-mediated radiosensitization, an exploration of other genes is needed, and more research is required to clearly prove the effect of these three genes in DAC-mediated radiosensitization.
The aim of this study was to explore whether treatment with DAC influences the response to irradiation in breast cancer cells. The next step is to undertake an in vivo study in order to clarify the potential benefit of DAC in radiation therapy. As has been reported, both demethylating agents and HDAC inhibitors have a synergistic effect, and, thus, the combined effect of both on radiosensitization needs further exploration. Furthermore, it had been shown that DAC has no synergistic effect with irradiation on growth inhibition in normal fibroblast cells, and the administration of drugs that can increase radiosensitivity might, thus, serve to avoid complications in normal adjacent cells in gastric cancer. 13 However, the evaluation of drug safety is imperative before clinical application in breast cancer. In 2006, DAC (decitabine) was approved by the U.S. Food and Drug Administration (FDA) for the treatment of de novo and secondary MDS (treatment-related MDS, t-MDS) and has gained popularity, given its favorable side-effect profile and its potential to improve survival. In the clinical trials, decitabine was generally safe and well tolerated. At its therapeutic dose, decitabine has favorable and manageable side effects, making it amenable to the elderly patient population in the treatment of MDS. 33 There is hope that DAC might also be useful in the treatment of solid tumors, but further studies are required to evaluate the safety and efficacy of DAC in solid tumors, especially in breast cancers. Finally, although DAC is a powerful demethylating agent, it has a number of drawbacks such as instability in aqueous solutions, which makes it difficult to be administered and quite toxic both in vitro and in vivo. Furthermore, the effects of the drug are transient, and DNA is gradually remethylated once the drug is stopped. Cytidine deaminase renders the drug inactivate by converting it into a 5-azauridine compound. 34 More work needs to be done for the optimization and reconstruction of the effects of DAC.
In conclusion, our results indicate a possible role for DAC as a radiosensitizer in breast cancer cells, a function that correlates with G2/M arrest, retardation of the repair of radiation-induced DSB, and reversal of the methylation status of genes that are associated with radioprotection or radiosensitization. These findings suggest that treatment strategies which incorporate DAC may be considered promising for the sensitization of breast cancer cells to irradiation.
Footnotes
Acknowledgments
This work was supported by the Applied and Basic Research Funds of Yunnan Province, China (No. 2008ZC075M and 2009ZC148M).
Disclosure Statement
No competing financial interests exist.