
Abstract
Laryngeal carcinoma, as a malignant tumor that occurs in the head and neck region, is widely treated by radiation, but in some cases, the cancer is radioresistant to the radiotherapy. The reason for the radioresistant response needs to be clinically understood. We designed our present study to identify the molecules that may be involved in this radioresistant response. In this study, we initially established the inherent radioresistant (Hep-2max) and radiosensitive (Hep-2min) cell lines from the parental laryngeal cancer cell line Hep-2. Furthermore, using microarray analysis, we identified a novel inherent radioresistance-related gene, phosphoprotein associated with glycosphingolipid-enriched microdomains1 (PAG1). We showed that siRNA directed against PAG1 in a radioresistant (Hep-2max) cell line dramatically enhanced the radiosensitivity and IR-induced cell death. On the contrary, ectopic expression of PAG1 in radiosensitive (Hep-2min) cell lines led to radioresistance and suppressed the IR-induced cell death. Taken together, our results indicate that the PAG1 gene may be a novel, promising radiosensitization target for laryngeal carcinoma.
Introduction
Head and neck cancers are the sixth most prevalent cancers in the world with a global incidence of ∼500,000 cases per year. 1 Cigarette smoke is one of the key causative agents for these cancers. Laryngeal carcinoma is dominated by squamous cell carcinomas, of which 67% arise in the glottic region, 31% in the supraglottis, and 2% in the subglottis. 2 Normally, patients with laryngeal carcinoma are treated with surgery, or radiotherapy alone, or adjuvant treatments. Although most patients with early-stage laryngeal cancer can be cured with radiotherapy, these outcomes have not improved over the past 2 decades, which is partly because of tumor radioresistance, including intrinsic radioresistance, which may exist before treatment, and radioresistance may greatly reduce the efficiency of radiotherapy for laryngeal carcinoma.
Increasing numbers of earlier reports suggest that radioresistance is often correlated with mutations or alterations of expression of specific genes, such as p53 and epidermal growth factor receptor. 3,4 Although such studies have contributed to a partial understanding of the mechanisms of cellular radioresistance, the molecular parameters that determine how a cell becomes more or less sensitive to DNA damage induced by radiation are poorly understood. The identification of molecular parameters to predict the response to radiotherapy before treatment is therefore urgently needed in clinical practice. 5
Previous studies have demonstrated that global gene expression profiling of radioresistant cancer cell sublines can elucidate the molecular mechanisms and pathways related to inherent radioresistance and can improve the efficacy of radiotherapy for cancers. 5 –7 Also, using the microarray technology, differentially expressed genes of tumor cells that have acquired radioresistance during radiation treatments were also identified in cancer cell lines, including esophageal, nasopharyngeal, and oral squamous cell carcinoma. 8 –10 Although ionizing radiation can lead to the development of radiation resistance and altered gene expression profiles, 11 the information on the mechanism underlying the inherent radioresistance in laryngeal carcinoma cells is still lacking.
In our present study, we focused on studying the molecular mechanisms underlying the inherent resistance to radiation in laryngeal carcinoma. Initially, we established a radioresistant and radiosensitive subline derived from one parent cell line Hep-2, and subsequently identified a novel radioresistance-related gene by using microarray analysis, phosphoprotein associated with glycosphingolipid-enriched microdomains1 (PAG1), which promotes the radioresistance in laryngeal carcinoma cells.
Materials and Methods
Cell lines and cell culture
The Hep-2 cell lines were purchased from the Cell Bank of the Shanghai Institute of Life Sciences (the Chinese Academy of Science). The cells were cultured in the DMEM (Gibco) supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin. Cultures were maintained at 37°C in a humidified incubator with 5% CO2.
Establishment of sublines
Single-cell clones were obtained using the limited dilution method. The logarithmic growth phase of Hep-2 cells was seeded on a 96-well plate (Corning) at 0.5 cells per well and then incubated at 37°C in a humidified incubator with 5% CO2. After the cells had grown for 10 days, they were treated with trypsin and subcultured into 24-well plates (Corning). This was repeated until confluence was reached at 20 days after single-cell cloning. Finally, six sublines were obtained, namely Hep-2-1, Hep-2-2, Hep-2-3, Hep-2-4, Hep-2-5, and Hep-2-6.
RNAi transfections
Three pairs of double-stranded siRNA oligonucleotides against the PAG1 gene were designed. Their sequences were PAG1-1: sense 5′-GCAGCAAGGCAGAGAUAUUdTdT-3′, antisense 3′-dTdTCGUCGUUCCGUCUCUAUAA-5′; PAG1-2: sense 5′-CUGGACAGUUAGUGAAUAAdTdT-3′, antisense 3′-dTdTGACCUGUCAAUCACUUAUU-5′; PAG1-3: sense 5′-GUGUUAAUGUAGAGAGUAUdTdT-3′, antisense 3′-dTdTCACAAUUACAUCUCUCAUA-5′. These siRNAs were synthesized by RIBOBIO Company. Approximately 2×105 cells were seeded per well in a six-well tissue culture dish, the day before transfection. siRNAs were transfected using RNAiMAX (Invitrogen). Cells were harvested for analysis after 48 hours before the time of transfection.
Transfection and expression of the human PAG1
The human PAG1 cDNA (NM_018440.3) was subcloned in-frame with a c-terminal Flag-tag downstream of the cytomegalovirus promoter into the pReceiver-M13 vector, and the pReceiver-M13-Flag vector was used as a control (GeneCopoeia). Cells seeded at 2.5×105 cells per well in a six-well tissue culture dish and were transfected with 2 μg plasmid DNA using Lipofectamine™ 2000 (Invitrogen). Cells were collected 48 hours after transfection.
Microarray analysis
To compare the differentially expressed genes in the three groups, an aliquot (2 μg) of the total placental RNA was used to synthesize double-stranded cDNA, which was subsequently transcribed into biotin-tagged cRNA using the MessageAmp II aRNA Amplification Kit (Ambion). The cRNA was then fragmented to produce strands of 35–200 bases in length in accordance with protocols (Affymetrix). The fragmented cRNA was hybridized to the Affymetrix GeneChip Human Genome U133 Plus 2.0 Array, containing 47,000 transcripts. Microarray hybridization was performed at 45°C with a rotation lasting for 16 hours using Affymetrix GeneChip Hybridization Oven 640. The arrays were washed and stained (streptavidin–phycoerythrin) at Affymetrix GeneChip Fluidics Station 450, and then later scanned on Affymetrix GeneChip Scanner 3000 to analyze the hybridization data. The scanned images obtained were assessed first by visual inspection and then were analyzed utilizing Affymetrix GeneChip Operating Software (GCOS 1.4).
qRT-PCR
qRT-PCR was conducted on two genes differentially expressed in the microarray. Total RNA was extracted using TRIzol reagent (Invitrogen), and 1 μg DNase-treated RNA was reverse transcribed with the Revert Aid™ First Strand cDNA Synthesis Kit (MBI Fermentas) according to the manufacturer's instructions. Amplification reactions were conducted using the SYBR Green/ROX qPCR Master Mix (Fermentas) with an ABI PRISM 7300 system. Gene-specific qRT-PCR primers were sense GAGTCCACCTACACCTC CATTC and antisense GCCTTTTCTTCCTCTCTGTTGA for PAG1; sense GCATCAACTTTGTGGCAATG and antisense ATAGAGGCCGATTTCCTTGGT for IL18; and sense GGTG GTCTCCTCTGACTTCAACA and antisense GTTGCTGTAG CCAAATTCGTTGT for GAPDH. The cycling conditions composed of 4-minute polymerase activation at 95°C and 40 cycles, 95°C for 30 seconds, 60°C for 30 seconds, and 72°C for 1 minute. Cycle threshold (Ct) values were obtained from the ABI PRISM 7300 system. The fold change of relative mRNA expression was determined using the 2-ΔCt method. 12 Each set of qRT-PCR was performed in triplicates and repeated thrice.
Western blotting
Cells were harvested and lyzed in a radioimmunoprecipitation buffer (Beyotime). Total protein (30 μg) was separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred onto polyvinylidene difluoride membranes. The membranes were incubated with a primary antibody, anti-Flag (1:500 dilution; ZSGB-BIO) and anti-β-actin (1:500 dilution; Beyotime), and then incubated a with horseradish peroxidase-conjugated secondary antibody (1:2000 dilution; ZSGB-BIO). Bands were visualized by employing the ECL Plus Detection System (Tiangen). The level of β-actin was used as a loading control.
Clonogenic assays
A defined number of cells were irradiated with 0, 1, 2, 4, and 6 Gy at room temperature using X-rays (4 Gy/min, SSD=98 cm, 15×15 cm2). After irradiation, cells were incubated at 37°C for 14 days and then fixed for 15 minutes with 3:1 methanol–acetic acid and stained for 15 minutes with 0.5% crystal violet (Sigma) in methanol. After staining, colonies with a cut-off of 50 viable cells were counted. The surviving fraction was calculated by using the equation (mean colony counts)/(cells plated)×(plating efficiency), where the plating efficiency was defined as (mean colony counts)/(cells plated for irradiated controls). Experiments were performed in triplicate.
Apoptosis measurement with flow cytometry
Apoptosis of cells was evaluated by double staining with the fluorescein isothiocyanate label Annexin V and propidium iodide (PI), according to the manufacturer's manual (Invitrogen). Cells were quickly washed twice in cold phosphate-buffered saline and resuspended in a binding buffer, and the mixtures were gently vortexed and incubated for 15 minutes at room temperature in the dark. After 1 hour, cells were analyzed with a flow cytometer (Beckman Coulter).
Statistical analyses
Data are shown as mean±standard deviation. Statistical Program for Social Sciences (SPSS) for Windows version 13.0 software (SPSS, Inc.) was used for statistical analyses. One-way analysis of variance was used to compare between the groups. p<0.05 was considered significant.
Results
Establishment of radioresistant and radiosensitive laryngeal carcinoma cells
The human laryngeal carcinoma cell line Hep-2 was used to develop sublines by limiting dilution assays in a preliminary experiment. 13 We obtained six sublines, which were named as Hep-2-1, Hep-2-2, Hep-2-3, Hep-2-4, Hep-2-5, and Hep-2-6, respectively. We first determined the cell survival rate of these six sublines by a classic clonogenic assay, after its exposure to various doses of radiation. As shown in Figure 1A, Hep-2-2 shows a markedly increased surviving cell fraction after irradiation, when compared to the parental cell line Hep-2 and other sublines, whereas Hep-2-6 showed relatively a decreased surviving cell fraction after irradiation. If the level of survival cells increases, a decrease in apoptotic cell death after irradiation is expected. This is indeed the case, as shown in Figure 1B and C. After 48 hours of irradiation, the apoptotic cells of Hep-2-6 were 22.96% markedly increased when compared to other sublines, whereas the level of apoptotic cells of Hep-2-2 were significantly lesser than that of other sublines (p<0.05). Taken together, these results indicate that Hep-2-2 is highly radioresistant, whereas Hep-2-6 is highly radiosensitive among the six sublines. In other words, we have successfully established the most radioresistant Hep-2-2 and radiosensitive Hep-2-6 cells from the parent cell line Hep-2, and also have designated Hep-2-2 and Hep-2-6 as Hep-2max and Hep-2min, respectively.
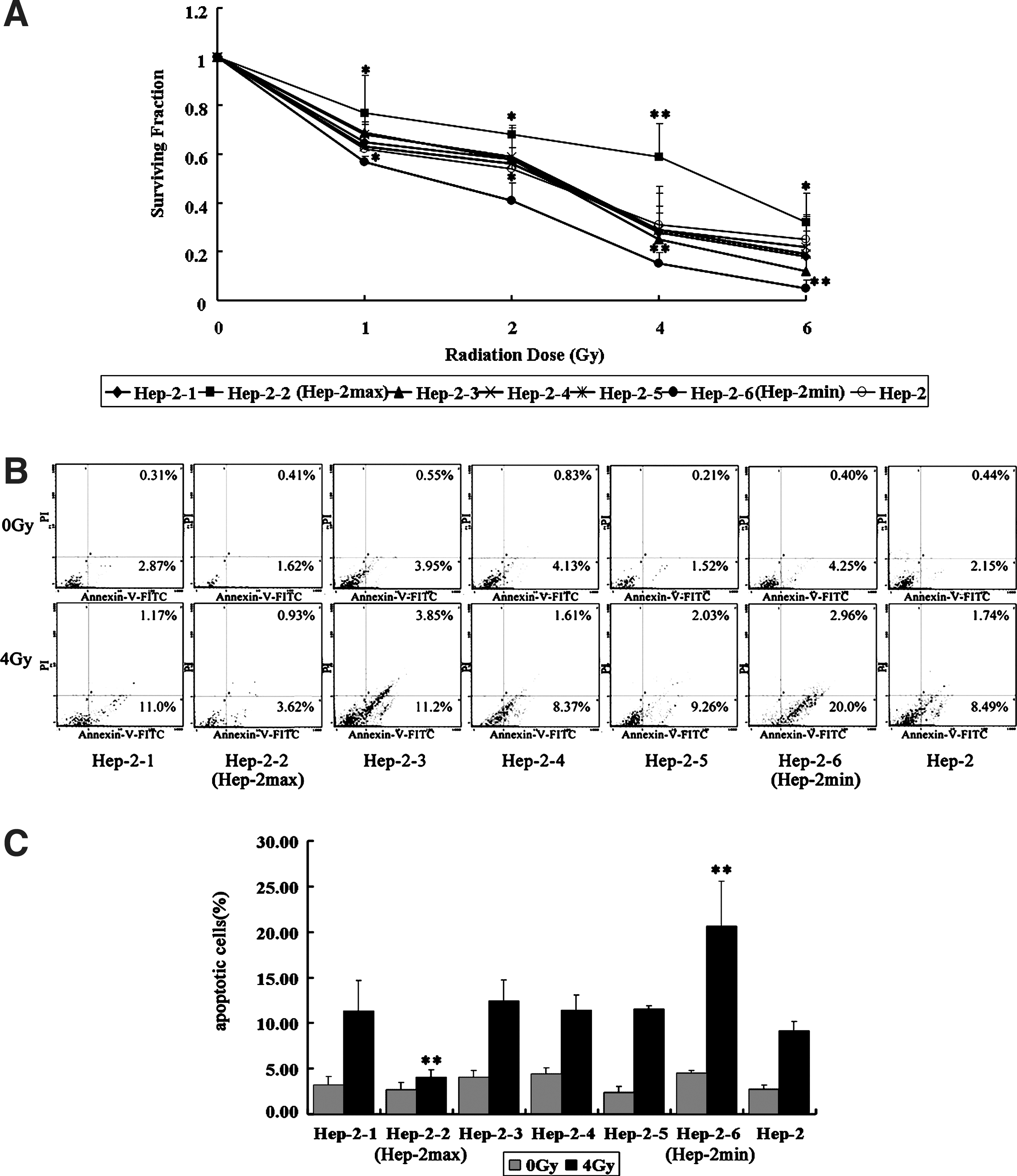
Radioresistant and radiosensitive sublines established from the parental cells.
Identification of differentially expressed genes in Hep-2max and Hep-2min cells
To screen the radioresistant-related genes, we used a cDNA microarray to identify differentially expressed genes in Hep-2max, Hep-2min cells, and the parent cell line Hep-2. After analysis of microarray data, 20 genes were certified to be significantly differentially expressed between the groups, with a threshold fold change over 2.0 or under 0.5. The list of 20 differentially expressed genes was used for unsupervised hierarchical clustering, and the results (Fig. 2A) were analyzed and visualized by the TreeView program. Finally, we focused on those genes whose expression was upregulated in Hep-2max cells and simultaneously downregulated in Hep-2min cells. As shown in Figure 2A, two genes PAG1 and IL18 comply with such conditions.
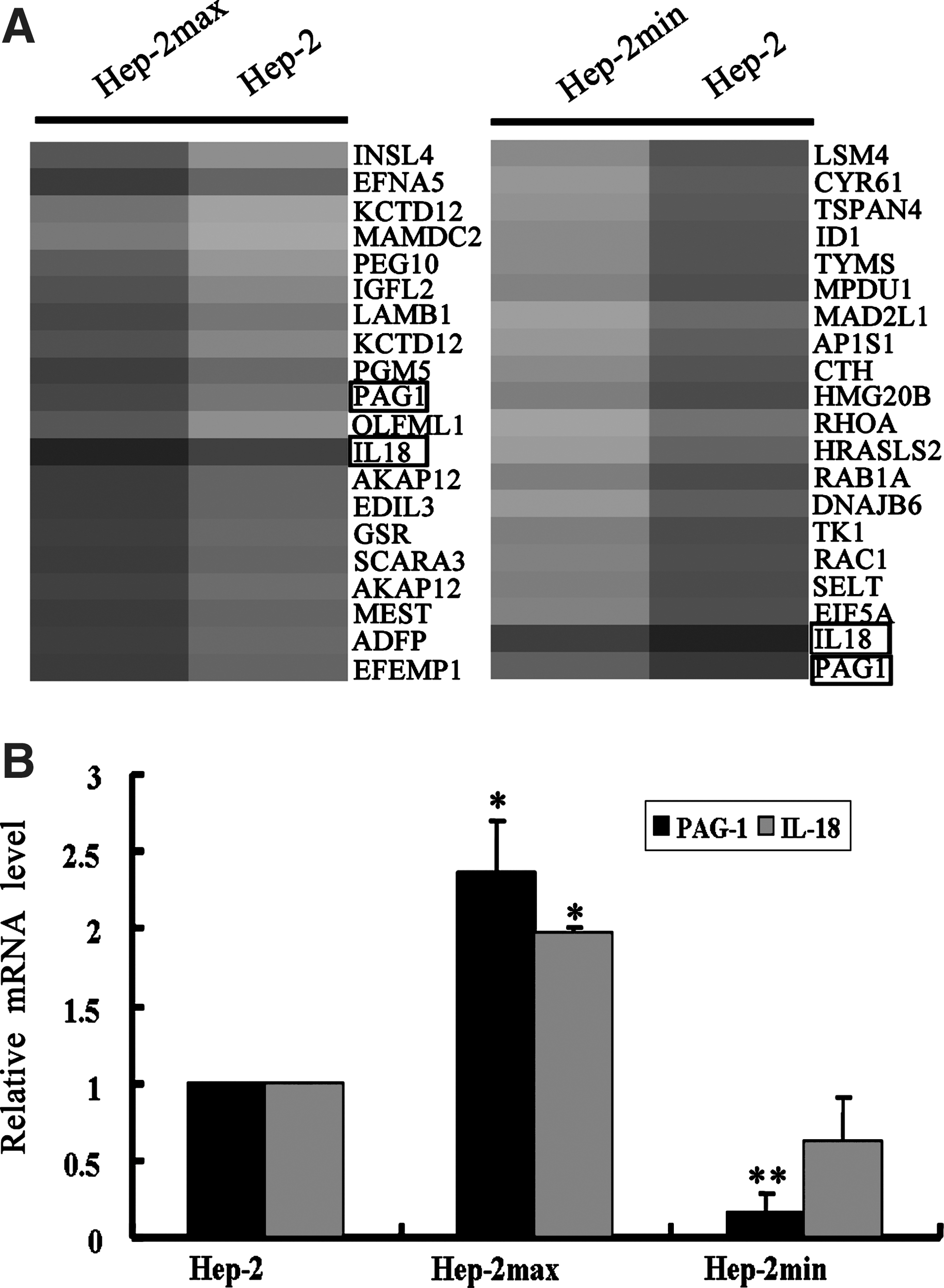
Identification of differentially expressed genes between the sublines and the parental cell lines.
To independently validate the results obtained from our microarray results, we further use the qRT-PCR to detect the expression of PAG1 and IL18 in Hep-2, Hep-2max, and Hep-2min cells. As shown in Figure 2B, the mRNA level of PAG1 was increased in Hep-2max and decreased in Hep-2min, on comparison with the parental cell line Hep-2, respectively. Its tendency of change was consistent with the microarray data. The difference was statistically significant (p<0.05). However, the expression of another gene IL18 in Hep-2min was not significantly downregulated, when compared with that in the Hep-2 (p>0.05). These results strongly suggest that PAG1 may be a novel molecule that is involved in the radioresistance of human laryngeal carcinoma cells.
Silencing of PAG1 enhances the radiosensitivity and IR-induced cell death in Hep-2max cells
To analyze whether PAG1 expression affects radiation-induced survival and cell death, we first synthesized three siRNAs specifically targeting different PAG1-coding regions and examined the knockdown effect of the PAG1-specific siRNA. As shown in Figure 3A, the interference effects of siRNA-PAG1-2 and siRNA-PAG1-3 were the strongest among the three siRNAs. Therefore, siRNA-PAG1-2 and siRNA-PAG1-3 were selected in the present study.

The role of PAG1 in the Hep-2max cell lines with response to siRNA transfection.
Next, to show the ability of the PAG1 siRNA transfection to sensitize Hep-2max cells, clonogenic survival assays were performed for Hep-2max cells. As shown in Figure 3B, the survival fractions for the siRNA-PAG1-2 and siRNA-PAG1-3 groups were significantly lower than those for the scrambled control siRNA (p<0.05 at the various doses). Consistent with this notion, the percentage of radiation-induced cell death in the siRNA control and siRNA-PAG1 groups was significantly higher in siRNA-PAG1-2 (31.4%) and siRNA-PAG1-3 (25.08%), as indicated by a 5.3- and 4.2-fold increase, respectively, when compared with the siRNA control group (5.85%, p<0.01) (Fig. 3C). Collectively, these results indicate that the depletion of PAG1 clearly increases the radiosensitivity of Hep-2max cells.
Overexpression of PAG1 increases the radioresistance and suppresses IR-induced cell death in Hep-2min cells
Since PAG1-knockdown Hep-2max cells exhibit an increased radiosensitivity, we summarized that the ectopic expression of PAG1 may be associated with the radioresistance. We further investigated whether PAG1 overexpression could enhance radioresistance. To test this possibility, PAG1-overexpressing Hep-2min cells and their respective control cells were treated with X-ray irradiation and analyzed with clonogenic survival assays and flow cytometry. We first detect the expression of exogenous PAG1, and western blotting analysis confirmed PAG1 overexpression in Hep-2min cells (Fig. 4A). As shown in Figure 4B, the survival fractions were increased when the PAG1 protein is overexpressed in Hep-2min cells. In addition, the IR-induced apoptotic cells (7.25%) in PAG1-overexpressing Hep-2min cells were significantly lower than in control cells (23.91%, p<0.01) (Fig. 4C). These results strongly suggest that the overexpression of PAG1 results in the radioresistance in Hep-2min cells.
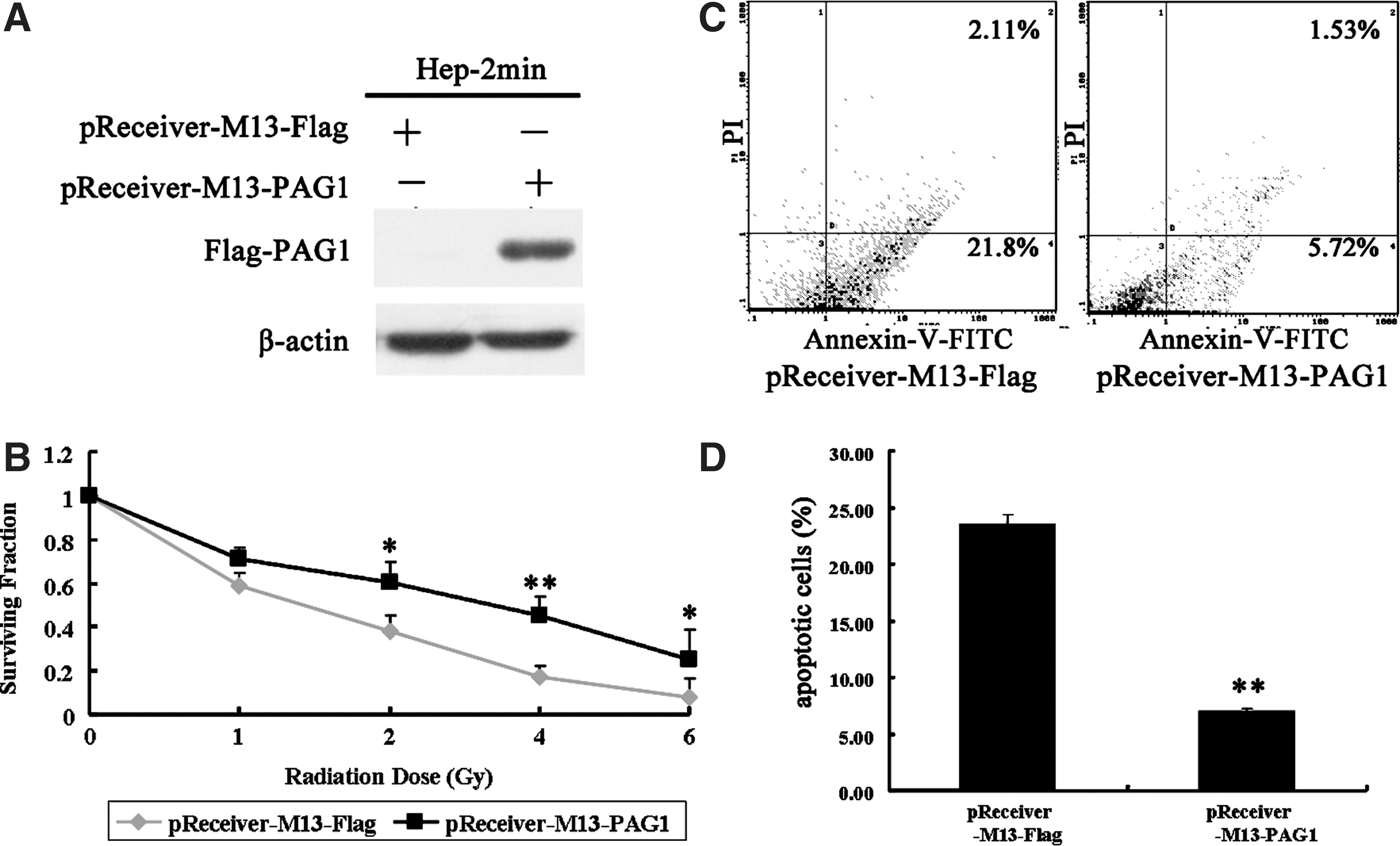
Effect of the overexpression of PAG1 in Hep-2min cells.
Discussion
In the present study, we have successfully established the inherent radioresistant and radiosensitive laryngeal carcinoma cells from the parental cell lines. Furthermore, we identified a novel molecule PAG1, which promotes the inherent radioresistance in the laryngeal carcinoma cells. To our knowledge, this is the first evidence that PAG1 may be implicated in inherent radioresistance in human laryngeal carcinoma cells.
Numerous lines of evidence have been presented that cancer cells will exhibit acquired radioresistance during radiotherapy, such as esophageal, nasopharyngeal, and oral squamous cell carcinoma. 8 –10 However, there are few reports about the inherent radioresistance in these cancers. In our present study, we initially construct the inherent radioresistant and radiosensitive laryngeal carcinoma cells (Fig. 1A, B). Furthermore, using microarray analysis, we found that one of the genes, PAG1, might be involved in cancer inherent radioresistance. Therefore, our results provided a good cell model for studying the inherent radioresistance of human laryngeal carcinoma.
Phosphoprotein associated with PAG1, which is also known as Csk-binding protein, was first fully characterized in 2000. 14 It was initially recognized as a ubiquitously expressed adaptor protein, recruiting cytoplasmic C-terminal Src-kinase to the close proximity of plasma membrane-anchored Src-kinases, thereby allowing Csk to impose its inhibitory potential on these kinases. 15 –17 Subsequently, PAG1 has been reported to involve in immune reactions and tumorigenesis. 18,19 For example, in human nonsmall cell lung cancers, PAG1 expression is significantly downregulated, and the overexpression of PAG1 results in the suppression of tumor growth by promoting c-Src inactivation. 20,21 According to the current opinions, these findings place PAG in a position of a tumor suppressor candidate. Some studies have revealed many tumor-related genes associated with radioresistance. 22,23 However, there is no report on PAG1, which participates in cancer radioresistance. Here, we found that the depletion of PAG1 enhanced the radiation sensitivity (Fig. 3A–C), whereas ectopic expression increased the radioresistance in human laryngeal carcinoma cells (Fig. 4A–C), indicating that PAG1 might be a novel radioresistance-related protein. These findings suggest that PAG1 inhibition may be a promising strategy to increase the radiotherapy effect in laryngeal carcinoma. However, how PAG1 involved in the cancer radioresistance and its detail mechanisms need to be further investigated in future. In addition, although our findings show that the expression level of IL18 in Hep-2min cells was not significantly decreased, we cannot exclude the possibility that IL18 participates in cancer radioresistance, which remains to be addressed in future studies.
In conclusion, we have demonstrated that PAG1 acts as a radioresistant factor in laryngeal carcinoma cells. This may shed new light on understanding of molecular mechanisms related to cancer-inherent radioresistance. Given that PAG1 plays an important role in radiosensitivity, we speculate that PAG1 may be a promising radiosensitization target for cancer radiotherapy, especially for the laryngeal carcinoma.
Footnotes
Acknowledgments
This work was supported by the National Natural Scientific Foundation of China Grant (No. 30800282).
Disclosure Statement
No institutional or commercial affiliations would pose a conflict of interest regarding the publication of this article.