
Abstract
Introduction
Telomerase is a ribonucleoprotein complex responsible for the maintenance of the length of telomeres, which are shortened during cell division. 1 Telomerase expression has been directly linked to tumor development, and inhibition of telomerase in human tumor cells may arrest tumor growth. 2 The human telomerase catalytic subunit human telomerase reverse transcriptase (hTERT) is expressed in many human cancer cells derived from multiple tissues, but infrequently in normal cells. 3 –5 Thus, hTERT is an attractive candidate target for tumor immunotherapy.
Immunotherapy targeting the hTERT subunit of telomerase has been shown to induce robust immune responses in cancer patients after vaccination with a single hTERT peptide. The hTERT:540–548 peptide (p540, ILAKFLHWL) was previously identified by investigators from two laboratories as an immunodominant HLA-A*0201-restricted T-cell epitope. 6,7 In both studies, it was reported that this peptide could induce CTL that recognized the peptide in vitro as well as cells that endogenously expressed telomerase, including breast, colon, lung, melanoma, and prostate cancers, and cells transfected with the full-length hTERT gene.
Although hTERT:540–548 peptide has been identified as an effective HLA-A*0201-restricted T-cell epitope, an effective T-cell-based cancer treatment, including vaccines, against hTERT will likely require the identification of other MHC class I-restricted epitopes in this antigen. 8 Thus, the search of CTL epitopes of hTERT needs to be extended to other epitopes.
The goal of this study was to identify immunogenic peptides of hTERT restricted by HLA-A*0201 molecule, which accounts for about 15% of the Chinese population. 9 As we reported here, the immunogenicity of six hTERT peptides that were predicted to bind to HLA-A*0201 molecules was analyzed. Through molecule simulation, it was found that two of the peptides (p540 and a new peptide p675, LLGASVLGL) were able to bind HLA-A*0201 molecule more strongly. The two peptide-specific CTL lines effectively recognized HLA-matched, telomerase-positive tumor cell lines, indicating that they are naturally processed and expressed epitopes, and p675 could serve as a new potential immunogen for cancer vaccines.
Materials and Methods
Peripheral blood samples
Peripheral blood mononuclear cells (PBMC) were obtained by density-gradient centrifugation from HLA-A2+ volunteers who gave written informed consent on protocols approved by the Institutional Review Board of Jinling Hospital. Donors were screened for HLA-A*0201 expression by PCR-SSP and DNA sequencing.
Peptide epitope prediction and molecular dynamics simulations
hTERT peptides that are predicted to bind to HLA-A*0201 were identified using three computer algorithms for epitope prediction: SYFPEITHI (
Peptide synthesis and HLA-A*02 binding
Peptides were synthesized by Shanghai ShineGene Molecular Biotechnology Co., Ltd. Purity was >95% as assessed by HPLC and mass spectrometry. All peptides were dissolved in a mixture of water and dimethyl sulfoxide. Binding to HLA-A*02 was confirmed using a TAP-deficient T2-cell-binding assay to measure cell surface stabilization of HLA-A*02 after overnight incubation with peptide in a serum-free medium. Surface HLA-A*02 was analyzed by flow cytometry.
Cell lines
Human lung adenocarcinoma cell lines A549 (HLA-A*0201+hTERT+) and SPC-A1 (HLA-A*0201−hTERT+) and the human breast cancer cell line MCF-7 (HLA-A*0201+hTERT+) 10,11 were maintained in our laboratory. These cell lines were grown in RPMI 1640 or DMEM (Hyclone) supplemented with 10% fetal bovine serum (Hyclone).
ELISPOT assay
ELISPOT analysis was performed after a 1-week in vitro culture of separated mononuclear cells from patients with peptide (10 μg/mL) and IL-2 (100 IU/mL). We cultured 3×105 to 3×104 cells per well, depending on the availability of the patient material, together with TAP-deficient HLA-A*0201+ T2 cells. After an overnight incubation, the activated cells were assayed in 20-hour ELISPOT cultures (96-well nitrocellulose microtiter plates; Millipore) for IFN-γ production examination. Briefly, microtiter plates were coated overnight with 50 μL per well of anti-IFN-γ monoclonal antibody (BD Pharmingen) at 10 μg/mL in a carbonate buffer, pH 9.6. Activated cells were added in at least triplicates. The plates were incubated in 37°C, 5% CO2 for 20 hours. After that, the plates were processed using biotinylated anti-IFN-γ (5 μg/mL, biotin rat anti-IFN-γ; BD Pharmingen), streptavidin-conjugated horseradish peroxidase (1:1000; DakoCytomation), and an alkaline phosphate conjugate substrate kit (BioRad). The number of spots was determined by a digitalized ELISPOT counter (Immunospot; CTL, Inc.).
Expansion of CD8+ T-cells and flow cytometry examination
For CTL line generation, PBMC from HLA-A*0201+ normal healthy donors were stimulated with hTERT-derived peptides in vitro in 96-well plates in the presence of interleukin-2 (5 and 10 IU/mL, thereafter) as described previously. By the limited dilution method, single-CTL-cell clone against hTERT-derived peptide was expanded by PHA (1–2 μg/mL) in the presence of high-dose interleukin-2 (100 IU/mL). PE-labeled pentamer against peptide-specific T-cell receptor (Proimmune) and PE-Cy5-labeled CD8 monoclonal antibody (BD Biosciences) were used to identified the expanded CTL clone by flow cytometry on FACSCaliber and CELLQuest software (BD Biosciences).
Cytotoxicity assay
The cytotoxicity of CTLs against target cells was quantified using the lactate dehydrogenase (LDH) release assay (Promega) according to the manufacture's instruction. Briefly, 1×103 cells/mL target cells (A549, SPC-A1, and MCF-7) were plated in 96-well plates in triplicate and incubated for 4 hours with CTLs at various E-to-T ratios. Maximal release of LDH was performed by completely lyzing the target cells. Target cells or effector cells alone were used as negative controls (spontaneous release). The killing rate was determined according to the formula [(experimental value−effector control value−target spontaneous value)/(target maximal value−target spontaneous value)]×100%.
Tumor inhibition assay
Specific pathogen-free BALB/c nude mice (male, ∼6 weeks; Model Animal Research Center of Nanjing University) were bred in our research institute. The mice were maintained under a 12-hour dark and 12-hour light cycle with food and water ad libitum. Viable A549 and SPC-A1 cells were harvested and diluted to a concentration of 107/mL in a phosphate buffer solution, and then 0.1 mL of the suspension was injected subcutaneously in the proximal of the left leg. The tumor volume was estimated daily by the formula 0.52×length×width 2 , in which the length and perpendicular width were measured by a caliper. When the tumor grew to about 100 mm3, the mice were randomly distributed into four groups (A549, A549-CTL, SPC-A1, and SPC-A1-CTL) (n=5) and treated with two doses of CTL (1×106) intravenously. Normal saline was used as control. The tumor growth rate was calculated by the formula (Tumor volume after treatment/Tumor volume before treatment-1). After the tumor growth was significantly different in groups, the tumor samples were excised and examined by hematoxylin and eosin staining. All mouse experiments were in accordance with the guidelines as approved by the Institute Animal Care and Use Committee of Jingling Hospital.
Statistical analysis
Data were expressed as mean±standard error of the means. Statistical Package for the Social Science (SPSS) version 13.0 was used for statistical analysis. Statistical significance was determined by one-way analysis of variance, and LSD was applied for multiple comparisons of means. p<0.05 was designated as statistically different.
Results
Peptide epitope prediction
Six peptides were predicted in hTERT with high scores by three computer algorithms (Table 1). Their binding energies to HLA-A*0201 were further analyzed by molecular simulation (Fig. 1). Of the six epitope peptides, p540 (ILAKFLHWL) and p675 (LLGASVLGL) were identified with lower binding free energies than the other four peptides (Table 2).

Molecule simulations of six predicted peptides (the binding of the peptides and HLA-A*0201). hTERT peptides predicted to bind to HLA-A*0201 were identified using three computer algorithms: SYFPEITHI, BIMAS, and NetCTL. The simulations of binding of the candidate peptides and HLA-A*0201 were then analyzed on the plate of Discovery Studio 2.5 (Accelrys). hTERT, human telomerase reverse transcriptase.
hTERT, human telomerase reverse transcriptase.
ETotal, total energy; EElect, electrostatic energy; EVdW, Van der Waals energy.
p540 and p675 strong binding to HLA-A*02
According with the molecular simulation, two peptides (p540 and p675) could bind with HLA-A*02 more firmly than the other four predicted peptides, as the two peptides could increase the fluorescence intensity of HLA-A*02 on the surface of T2 cell (Table 3).
p540 and p675 elicited stronger IFN-γ secretion of patients' PBMC
The reactivity of NSCLC patients against selected peptides was also assessed using IFN-γ ELISPOT assay. Ten HLA-A*0201+ patients with nonsmall-cell lung cancer were analyzed, and seven patients show similar results. As shown in Figure 2, peptides p540 and p675 could elicit more IFN-γ secretion than the other four peptides.

ELISASPOT assay of one HLA-A*0201+ patient's PBMC. Peptide p540 and p675 could elicit more IFN-γ secretion than the other four peptides. PBMC, peripheral blood mononuclear cells. # p<0.05 compared with other peptides; *p<0.05 compared with the control.
Expansion of CD8+ T-cells and flow cytometry examination
We further expanded the CTL clones against the peptides p540 and p675. After three cycles of expansion, the purities of CTL clones were about 96.08% and 96.24%, respectively (Fig. 3).
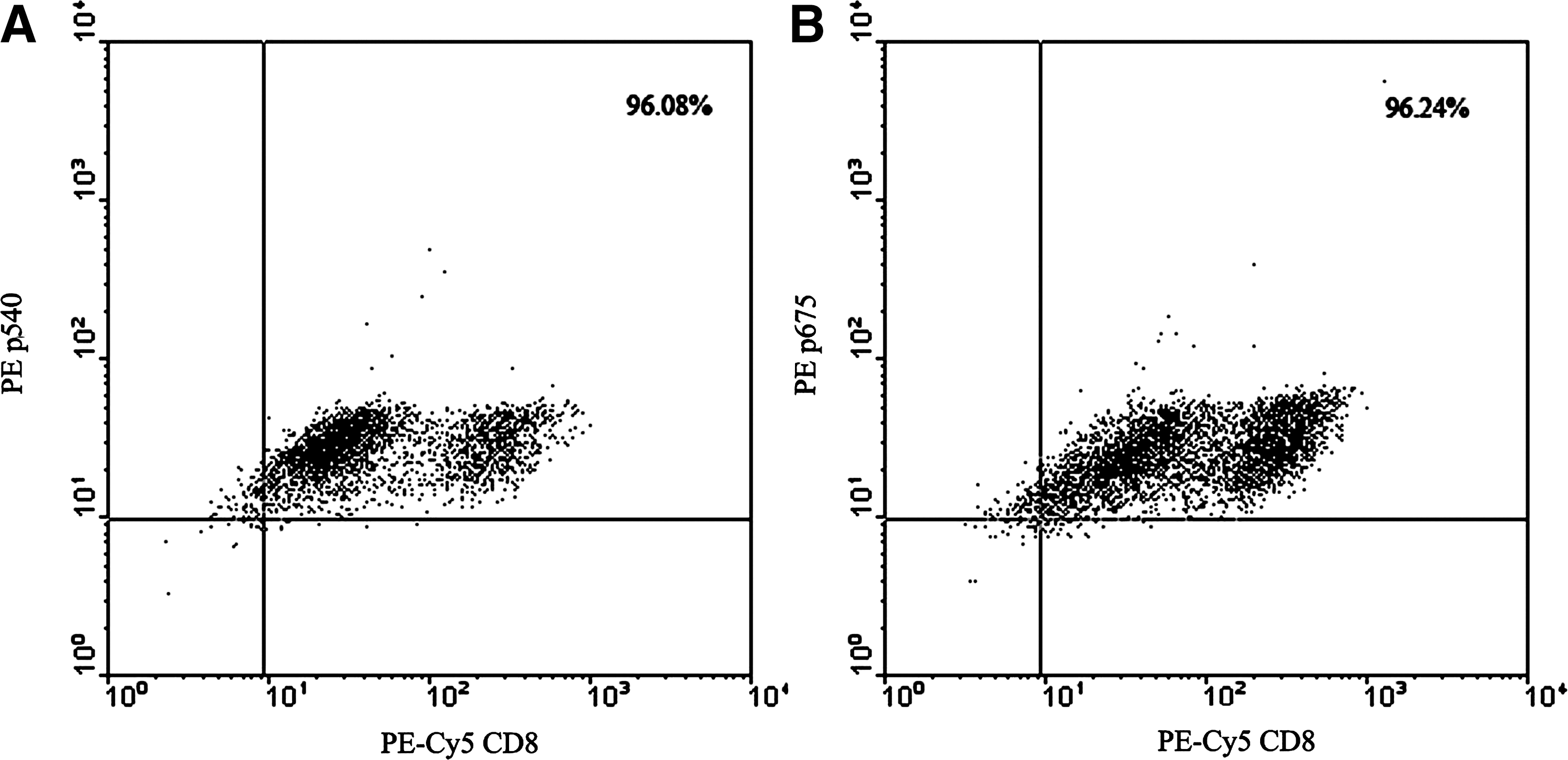
Flow cytometry examinations of expanded CTL clones against the peptides p540
CTL clone against p675 elicited significant cytotoxic effect HLA-A*0201 restrictly
We then assessed the cytotoxic effect of the CTL clone against p675 by LDH release assay. As shown in Figure 4, the CTL clone against p675 could lyze the target tumor cells with an HLA-A*0201-restricted manner, and HLA-A*0201-positive A549 and MCF-7 cells were more sensitive to the CTL cells.
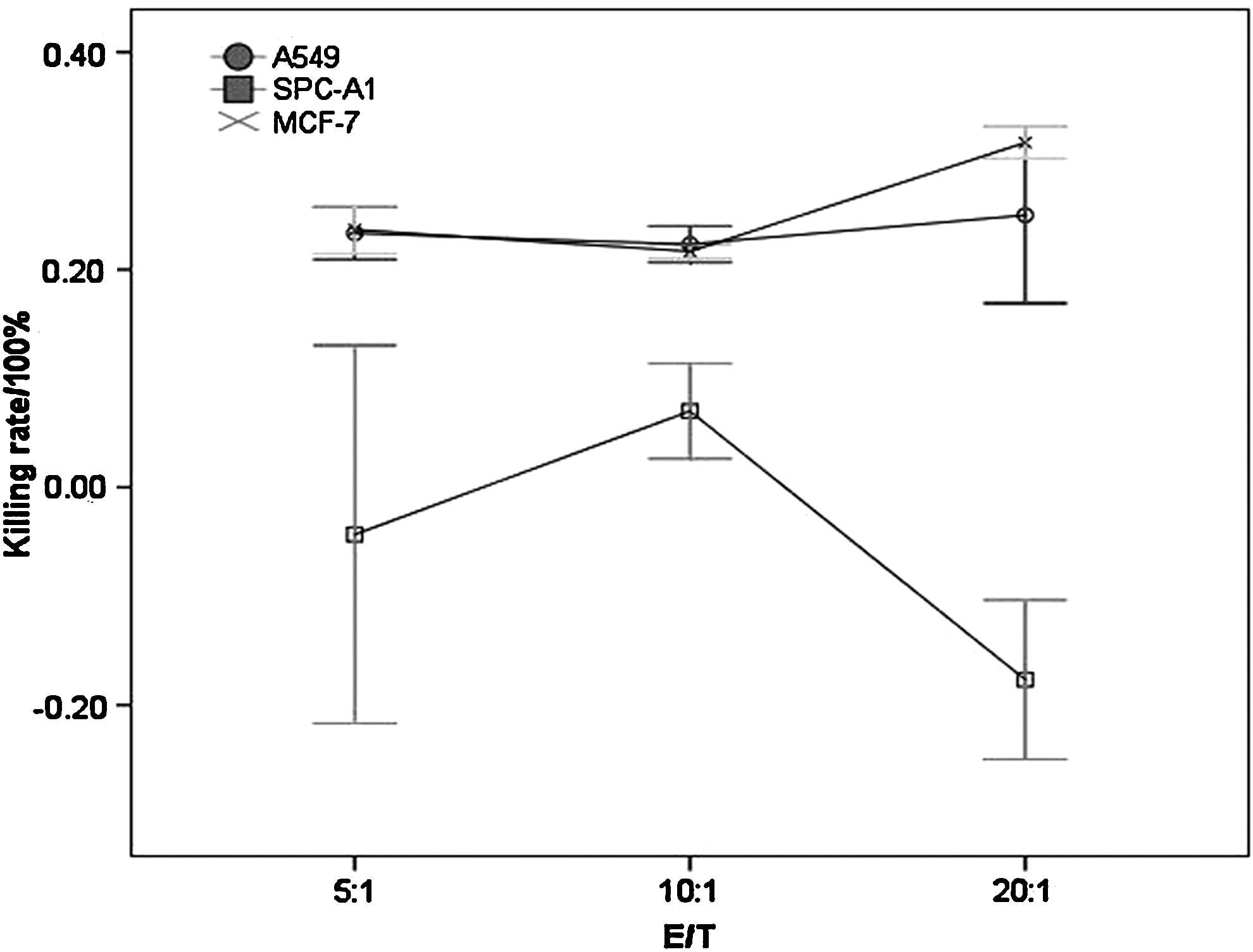
Cytotoxic effects of the CTL clone against p675. A549 (HLA-A*0201+hTERT+), SPC-A1 (HLA-A*0201−hTERT+), and the human breast cancer cell line MCF-7 (HLA-A*0201+hTERT+) were selected as target cancer cells. The cytotoxic effects of the CTL clone against p675 were analyzed by LDH release assay. The killing rate was determined according to the formula [experimental value−effector control value−target spontaneous value)/(target maximal value−target spontaneous value)]×100%. Of three effector/target ratio (5:1, 10:1, and 20:1), HLA-A*0201-positive A549 and MCF-7 cells were all more sensitive to the CTL cells than HLA-A*0201-negative SPC-A1 cells. LDH, lactate dehydrogenase.
CTL clone against p675 could inhibit tumor growth HLA-A*0201 restrictly
In accordance with the cytotoxic effect, the CTL clone against p675 could also inhibit the tumor growth with an HLA-A*0201-restricted manner. The growth of HLA-A*0201-positive A549 lung cancer was significantly suspended by the CTL clone against p675 (Fig. 5). Although weakly, the CTL clone against p675 could also inhibit the growth of HLA-A*0201-negative SPC-A1 lung cancer in vivo. However, the CTL could not inhibit the growth of other HLA-A*0201-negative lung cancer models (Supplementary Fig. S1). Further histochemistry examination indicated that the effect of tumor inhibition by the CTL clone against p675 might be related with the infiltration of lymphocytes in the tumor section (Fig. 6).
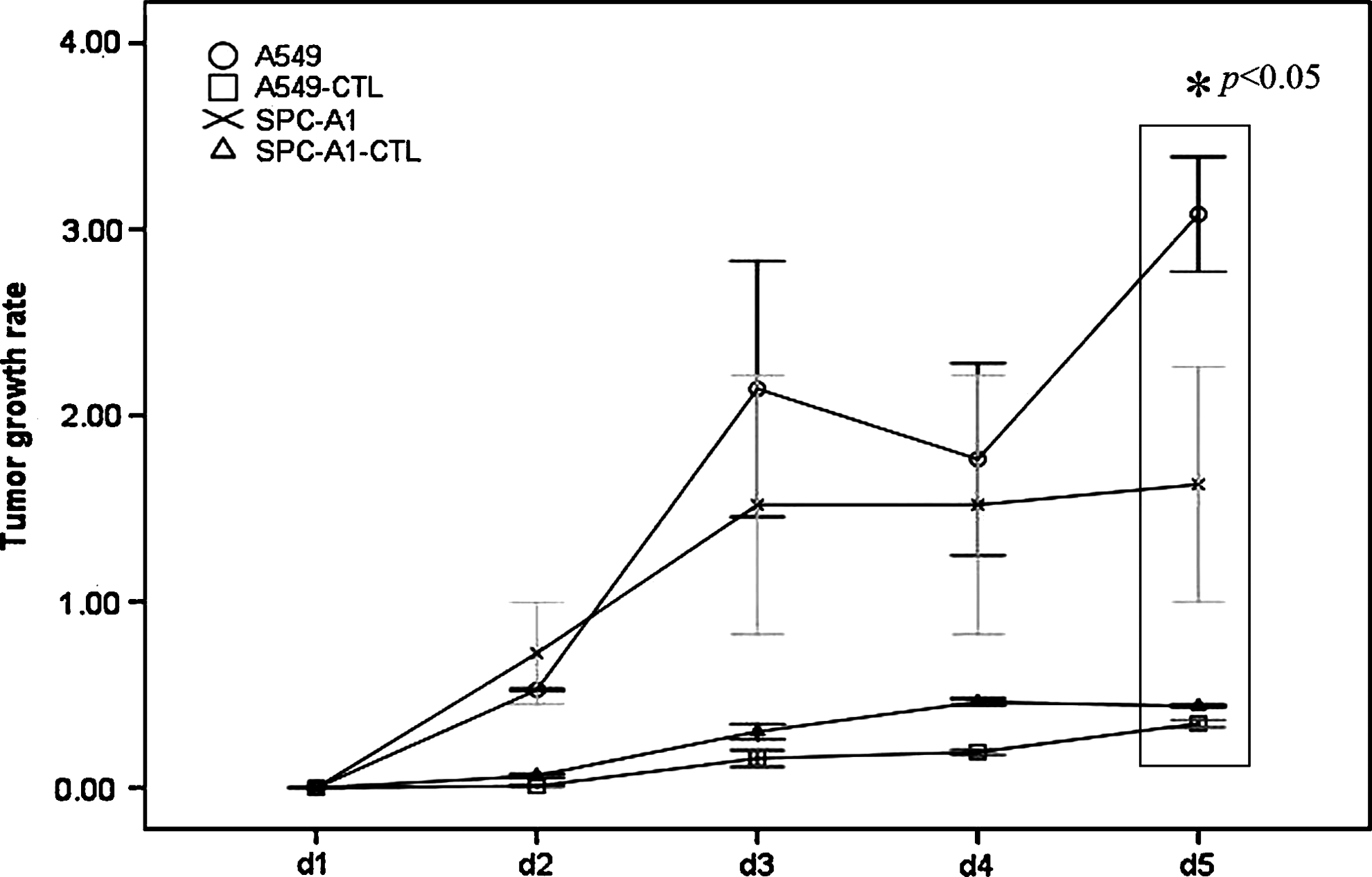
Tumor growth suppression effects of the CTL clone against p675. Xenograft models of A549 (HLA-A*0201+hTERT+) and SPC-A1 (HLA-A*0201−hTERT+) were initiated for the analysis of tumor growth suppression effects. Tumor growth rate was calculated by the formula (Tumor volume after treatment/tumor volume before treatment-1). The CTL clone against p675 could significantly inhibit the tumor growth of A549 lung cancer (HLA-A*0201-positive), compared with the effects on HLA-A*0201-negative SPC-A1 lung cancer.
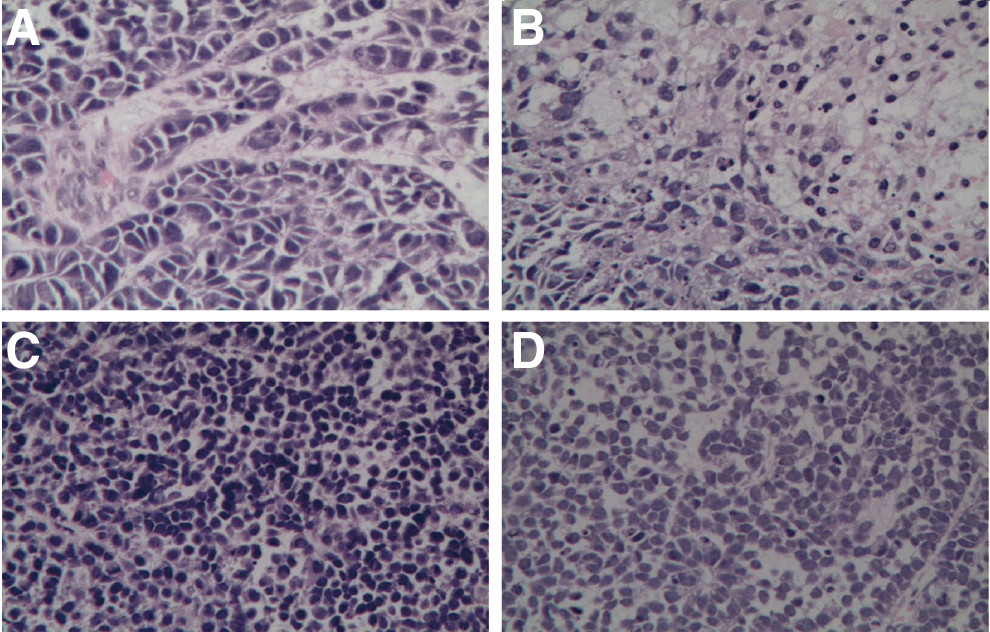
The infiltration of lymphocytes in the tumor section after adoptive transfer of CTL cells against p675.
Discussion
The identification of tumor-associated antigens and epitopes has enabled the development of novel immunotherapies for the treatment of patients with cancer. 12 Multiple tumor-associated antigens and specific class I MHC-restricted epitopes have been identified that are capable of being recognized by CTLs, including melanocyte differentiation antigens (e.g., MART-1, gp100, and tyrosinase-related proteins), cancer testis antigens (e.g., NY-ESO-1 and the MAGE family of proteins), and mutated proteins (e.g., MUM-1 and CDK-4). 13
The ribonucleoprotein telomerase has also been suggested to be a tumor-associated antigen. The telomerase catalytic subunit hTERT 4 is the rate-limiting component in the telomerase complex and is most closely correlated with telomerase activity. More than 85% of human cancers have telomerase activity and express hTERT, 5 whereas most normal adult human cells do not maintain the lengths of their telomeres. Therefore, telomerase was considered to be an attractive candidate target antigen for the development of immunotherapies for the treatment of patients with a variety of human cancers.
The search for immunogenic hTERT-derived peptides is a formidable task, but a necessary step to develop immunotherapy that targets hTERT on tumor cells in the widest assortment of the human population. For the most common HLA-A*0201 type, previous studies identified the immunogenic CTL epitope p540–548, which was also verified in the present study and used as positive control. 10
However, the processing of epitope peptides in hTERT was rather complex in cancer cells, and some reports indicated that the epitope p540 was not always presented on the cancer cell surface. 14 It is urgently needed for identifying other candidate T-cell epitopes of hTERT.
Bioinformatic analyzation and molecular dynamics provide powerful and flexible tools for the prediction and analysis of molecular and macromolecular systems. 15 A reliable approach to obtaining predictions of peptide–MHC binding is based on so-called molecular dynamic simulation. This technique attempts to calculate the free energy of binding for a given molecular system, which is closely related to experimentally observable quantities such as equilibrium constants. Essentially, all that is required is the experimentally determined MHC peptide complex. In the present study, we utilized this tool to analyze the binding of the predicted peptides and HLA-A*0201, and all the simulated results were in accordance with the experimental data. Peptide p675 could bind with HLA-A*0201 more firmly compared with other predicted peptides, indicating that p675 could be a new effective epitope in hTERT.
PBMC of nonsmall-cell lung cancer patients could be reactivated by p540 and p675, rather strongly as indicated by ELISPOT assay. However, the expansion and activity of the specific cytotoxic T cells were inhibited by various immunosuppressive conditions in the tumor microenvironment. 16 In vitro expansion of these CTLs might be a convenient method to deal with this condition. In the present study, the CTLs could be expanded more than 90%, and these expanded CTLs show significant tumor suppression effects.
However, the activated CTLs went to senescence about 1 week later. As a result, the expanded CTLs could only elicit a limited antitumor effect. Further studies are needed to clone the T-cell receptor of these epitope peptide-specific CTLs for gene therapy. 17
In conclusion, besides peptide p540, we identified p675 as a new HLA-A*0201-restricted immunogenic T-cell epitope, and the expanded CTL clone against it could suppress tumor growth effectively.
Footnotes
Acknowledgment
The work was supported by the National Natural Science Foundation of China (No. 30872979).
Disclosure Statement
The authors declare no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.