
Abstract
Cyclin-dependent kinase-like 1 (CDKL1) is a member of cell division control protein 2 (CDC2)-related serine–threonine protein kinase family, and it is likely to occur in malignant tumors, plays an important impact on the progress. This study aimed to evaluate the expression of CDKL1 in breast cancer and regulation in cancer cell growth. In the work, the CDKL1 mRNA level in fresh biopsy tissues from 186 breast cancer patients, with 98 benign tissues as negative control, and CDKL1 protein in 30 paraffin-embedded tissues from primary breast cancer patients were detected by the real-time reverse transcriptase–polymerase chain reaction (RT-PCR) assay and immunohistochemical staining, respectively. The roles of CDKL1 in cell growth were analyzed with CDKL1 short hairpin RNA (shRNA) inhibitor-transfected cells. CDKL1 was overexpressed in breast cancer patients and had a positive detection efficiency of 77% (144/186), which showed statistically significant difference compared with estrogen receptor (ER), progesterone receptor (PR), P53, vascular endothelial growth factor (VEGF), and E-cadherin (E-cad) (p<0.05). Inhibiting CDKL1 function with shRNA, MCF-7 cells exhibited obvious accumulation at the G2/M phase and increased sensitivity to cell cycle chemotherapeutic drugs. The results suggested that the CDKL1 gene could be a potential tumor marker for diagnosis and a gene target for therapy.
Introduction
Breast cancer has been remaining an important public health problem for women in the world. Although the long-term survival of breast cancer has not varied since 1980s, its prevalence rate has increased. 1 To improve the long-term survival, early diagnosis and treatment have attracted a considerable interest. A potential strategy for improving the survival rate and achieving early diagnosis and treatment will be to enhance the efficiency of diagnosis and the sensitivity of chemotherapy. Thus, probing the new biomarkers or therapeutic targets for breast cancer diagnosis and chemotherapy has become a pressing project.
Cells generally utilize the checkpoint mechanisms to coordinate cell cycle progression and cellular response to cell cycle irregularities, thereby maintaining accurate transmission of the genetic material during cell division. 2,3 Cell division control protein 2 (CDC2) is a key molecule in mitosis. Cyclin-dependent kinase-like 1 (CDKL1, CDC2-related kinase) is a member of CDC2-related serine–threonine protein kinase family. 4 CDKL1 is identified as a human homolog of cyclin-dependent kinases (CDKs) by low-stringency cloning studies, and named after the amino acid sequence corresponding to the PSTAIRE motif of CDC2 (subdomain III). 5,6 However, up to now, the function of CDKL1 in cell regulation has not been well known.
In this study, we first reported that CDKL1 was overexpressed in Chinese breast cancer patients. The CDKL1 mRNA levels in fresh biopsy tissues from both patients with breast cancer and benign controls and the correlations between the CDKL1 expression and clinicopathological variables were analyzed. To further explore the CDKL1 biological function, CDKL1 shRNA (short hairpin RNA)-interfering sequences were inserted into the pGPU/GFP plasmids to interfere with CDKL1 expression in MCF-7 cells. The changes in cell cycles and cell proliferation of CDKL1-silencing MCF-7 cells indicated that CDKL1 was involved in cell cycle regulation.
Materials and Methods
Patients and sample collection
One hundred eighty-six fresh biopsy tissues from patients with breast cancer, who were diagnosed at Nanjing Medical University Cancer Hospital (Nanjing, Jiangsu, P.R. China), between 2008 and 2010, and 98 benign tissues that were 5-cm away from the cancer tissues were enrolled in this study. The diagnosis, clinical stages, and pathological features of primary tumors of the patients were defined according to the criteria of the American Joint Commission on Cancer. The clinicopathological characteristics of the patients, including age, tumor size, histology grade, lymph node metastasis, and menopause, are summarized in Table 1. Patients who received preoperative treatment, such as radiation therapy and chemotherapy, were excluded from this work. Fresh biopsy tissues from 186 primary breast cancer patients and 98 benign tissues were collected in compliance with and approved by the Institutional Review Board and with informed consent from all the participants. Thirty paraffin-embedded tissue samples were obtained from the Department of pathology, which have been fixed in formalin, and routinely processed and embedded in paraffin.
Real-time reverse transcriptase–polymerase chain reaction
Total RNA was extracted using the Total RNA Isolation System (Promega) in accordance with the manufacturer's instructions, and analyzed by gel electrophoresis. The purity of the extracted RNA was determined by the ratio of absorbency at 260 and 280 nm. cDNA was synthesized from 1 μg of total RNA using reverse transcriptase, and was used as template for real-time polymerase chain reaction (PCR) assay (LightCycler1.5; Roche) with CDKL1-L primers (Table 2). Each analysis contained cDNA derived from 100 ng total RNA.
CDKL1, cyclin-dependent kinase-like 1.
The cDNA of MCF-7 cells was subjected to PCR amplification of CDKL1 mRNA, and the products were cloned into pMD18-T vector (TaKaRa). The exact sequence of the inserted plasmids was analyzed with CDKL1-L primers. Serial dilutions (from 1 to 1×106 copies/μL) of the recombinant CDKL1-pMD18-T vector were used to obtain the standard curve. CDKL1 expression was undetectable when the crossing point (Cp value) for CDKL1 was 36 cycles and above. All samples were measured in duplicate. After PCR, a melting curve was constructed by increasing the temperature from 65°C to 95°C, with a temperature transition rate of 0.1°C/s. To ensure that the correct product was amplified in the reaction, all samples were separated by 1% agarose gel electrophoresis. Single- and sharply defined melting curves with narrow peaks were obtained by Tm-calling analysis. The real-time PCR amplification efficiency was calculated from the given slopes in LightCycler software.
Immunohistochemical staining
Paraffin-embedded tissue sections were rehydrated before staining. To reduce nonspecific hydrophobic interactions between the primary antibodies and the tissue, the section was first blocked with one to three drops of serum-blocking reagent for 15 minutes, and then the resulting sample was incubated sequentially with anti-CDKL1 mAb (Abcam) and horseradish peroxidase-conjugated rabbit anti-mouse IgG Ab (DAKo). After incubation with 3, 3′-diaminobenzidine solution, visualized staining of tissue was observed under a microscope using a bright-field illumination.
Construction of interfering plasmids and transfection
Four shRNA-interfering sequences, named shRNA-381, shRNA-709, shRNA-649, and shRNA-963, were synthesized and inserted into pGPU/GFP plasmids, respectively. One independent interfering sequence was synthesized and used as a negative control.
MCF-7 cells were maintained in an RPMI-1640 medium, supplemented with 10% fetal bovine serum. Cell cultures were incubated in air containing 5% CO2. After the formation of a confluent monolayer, the cells were incubated in phosphate-buffered saline (PBS) containing 0.53 mmol/L ethylenediaminetetraacetic acid and 0.05% trypsin for 5 minutes at 37°C. Before transfection, the cell viability was assessed by trypan blue staining.
Five experimental groups were designed in six-well culture plates: one negative control group and four experimental groups, which were transfected with shRNA-381, shRNA-709, shRNA-649, shRNA-963, respectively. Lipofectamine 2000 (Invitrogen) was used as a medium according to the manufacturer's instructions.
Relative real-time PCR
We used real-time PCR and the 2−ΔΔCp method to detect the levels of CDKL1 mRNA with CDKL1-S primers (Table 2). β-Actin was used as a reference gene. Samples were amplified with a thermal denaturation at 94°C for 30 seconds, followed by 40 cycles of denaturation at 94°C for 10 seconds, annealing at 52°C for 15 seconds, and extension at 72°C for 15 seconds. After PCR, a melting curve was constructed by increasing the temperature from 65°C to 95°C, with a temperature transition rate of 0.1°C/s.
Western blot and flow cytometric assay
MCF-7 cells were harvested after transfection for 48 hours and lyzed in an RIPA buffer. Twenty-five micrograms of the sample was first electrophoresed on a 12% sodium dodecyl sulfate–polyacrylamide gel and then transferred to a polyvinylidene difluoride membrane. After saturated for 1 hour with Tris-buffered saline (pH 7.6) containing 5% skim milk and 0.1% Tween-20, the membrane was incubated with anti-CDKL1 mAb at 1:500 dilution, followed by horseradish peroxidase-conjugated rabbit anti-mouse IgG Ab at 1:200 dilution, and visualized by chemiluminescence (Thermo).
After transfection for 48 hours, MCF-7 cells were trypsinized, centrifuged at 1000 rpm for 5 minutes, and washed twice with PBS. The cells were fixed in 70% ethanol at−20°C. After the cells were resuspensed in 1 mL propidium iodide solution (100 μg/mL) for 30 minutes at 37°C, the suspension was filtered through a nylon monofilament mesh screen with 300 μm, and the obtained suspension was used for the flow cytometric analysis.
Cell proliferation assay
MCF-7 cells were harvested after transfection for 24 hours, stained with trypan blue, counted, and cultured in a 96-well plate at 4000 cells/well. After cultivating for 12 hours, the chemotherapeutic drugs, 5-fluorouracil (25 μg/mL), epirubicin (1 μg/mL), and docetaxel (10 μg/mL), were added in the wells, respectively. Five duplicates were made at each drug. After further cultivating for 48 hours, 20 μL MTT (5 mg/mL) was added in each well and incubated for 4 hours. One hundred fifty microliters of dimethyl sulfoxide was then added in the well to dissolve the solid matter, and the absorbance was detected at 590 nm.
Statistical analysis
The differences in clinicopathological variables were investigated using the McNemar test. p<0.05 values were considered to be statistically significant.
The data from drug sensitivity test were expressed as mean±standard deviation. The difference between the experimental groups and control group was assessed by the Student's t-test. It was considered to be statistically significant only when p<0.01.
Results
Breast cancer patients express high levels of the CDKL1 gene
Table 1 shows the clinicopathological characteristics of the 186 female patients, including age, tumor size, histology grade, lymph node metastasis, and menopause. The average age of these patients was 51 years, and 90 patients (48%) were aged below 50 years, and 114 patients (61%) were premenopausal. The tumor sizes of 165 patients (89%) were diagnosed larger than 2 cm, and 21% (39 out of 186) tumors were classified as grade I and 79% (147 out of 186) as histology grade II or III. Eighty-seven patients (47%) had tested tumor positivity for infiltrated auxiliary lymph nodes of breast cancer at the time of diagnosis. The positive detection efficiency of estrogen receptor (ER), progesterone receptor (PR), human epidermal growth factor receptor-2, P53, vascular endothelial growth factor (VEGF), epidermal growth factor receptor (EGFR), and E-cadherin (E-cad) was 44%, 27%, 71%, 48%, 92%, 76%, and 66%, respectively (Table 3).
p<0.05 was considered to be statistically significant.
ER, estrogen receptor; PR, progesterone receptor; C-erbB2, human epidermal growth factor receptor-2; VEGF, vascular endothelial growth factor; EGFR, epidermal growth factor receptor; E-cad, E-cadherin.
Real-time reverse transcriptase–polymerase chain reaction (RT-PCR) assay showed that in 186 fresh biopsy tissues, 144 cases (77%) had positive amplification. All of the 98 benign tissues had no amplification by comparing with the standard curve. CDKL1 had a statistically higher positive detection efficiency compared with ER, PR, P53, VEGF, and E-cad (p<0.05) (Table 3).
Breast cancer tissues overexpress CDKL1 protein in the cell nucleus
We used immunohistochemical staining to detect CDKL1 expression in a small cohort of 30 paraffin-embedded tissue samples of breast cancer with different histology grades (Fig. 1). The results showed that CDKL1 protein was overexpressed in the nuclei of tumor cells.
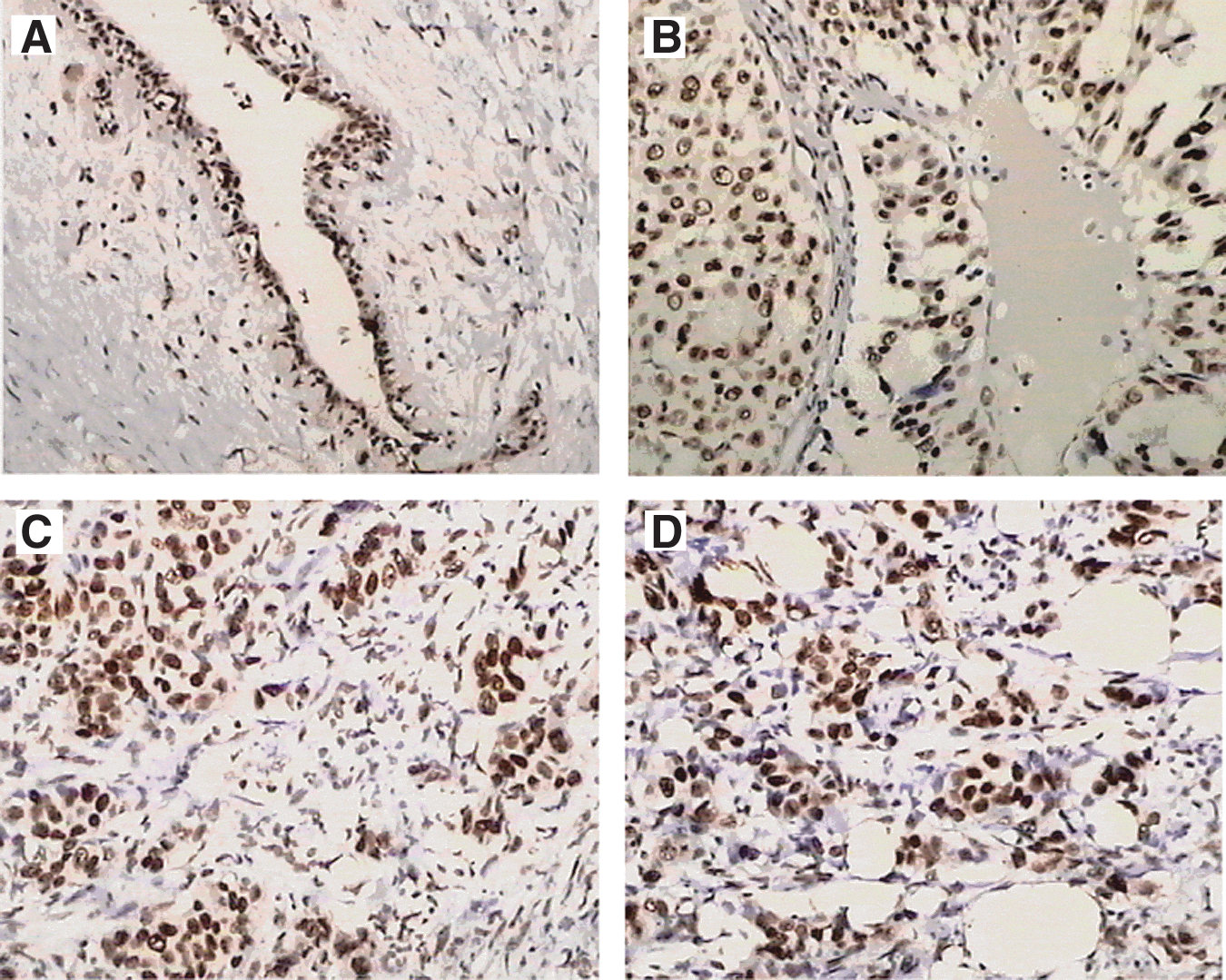
CDKL1 protein expression in breast cancer tissues by immunohistochemical staining. Positive staining of antibody to CDKL1 was exhibited in breast tumors of histology grade I
Effective inhibition of the expression of CDKL1 in MCF-7 cells
We used inverted fluorescence microscope to detect GFP expression after transfecting interfering plasmids into MCF-7 cells for 24 hours. The fluorescent intensity of the transfected cells was related to the transfection efficiency. As shown in Figure 2, the transfection efficiency was more than 70%.
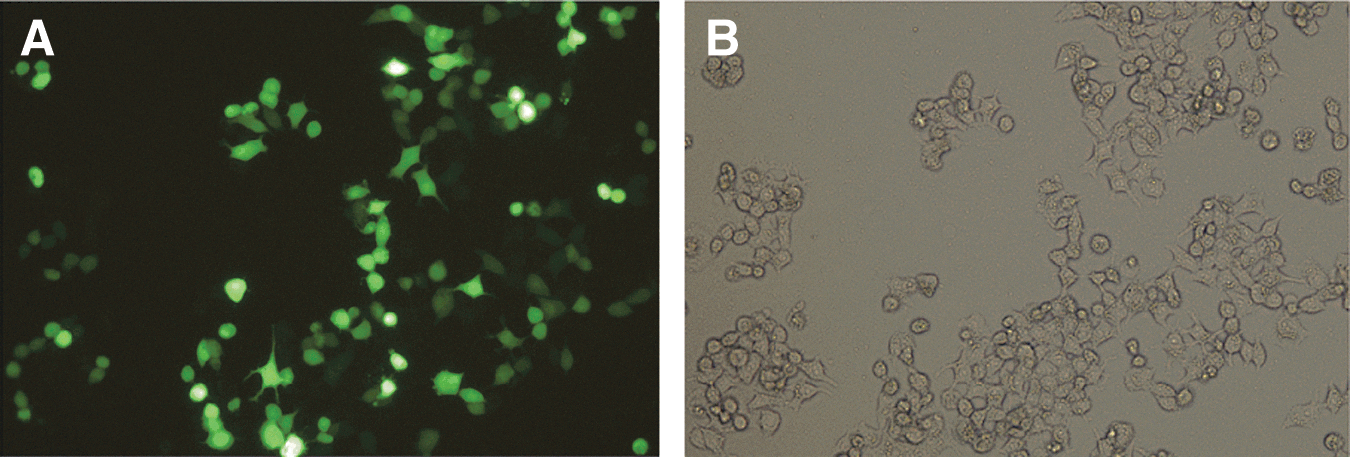
Inverted fluorescence microscopic images of MCF-7 cells after transfection. The transfection efficiency was evaluated by comparing fluorescent-field
By the relative real-time PCR assay, the levels of CDKL1 mRNA in MCF-7 cells transfected with shRNA-381, shRNA-649, shRNA-709, or shRNA-963, which were normalized to the negative control 1.00 (0.97–1.03), were 0.75 (0.73–0.77), 0.39 (0.37–0.41), 0.19 (0.14–0.25), and 0.39 (0.32–0.49), respectively (Table 4). shRNA-709 is the most effective interfering sequence for CDKL1 mRNA, as it inhibits the expression of CDKL1 mRNA, mostly.
cDNA was reversely transcribed from
Cp, crossing point; shRNA, short hairpin RNA.
Western blot was used to detect the CDKL1 protein expression in MCF-7 cells after interference. As shown in Figure 3, all these four shRNA-interfering sequences had a slight effect on the β-actin expression, whereas some of them could clearly downregulate the protein expression. Similar to the interference effect on CDKL1 mRNA expression, shRNA-709 also inhibited CDKL1 expression mostly, indicated that shRNA-709 is the most effective interfering sequence for CDKL1 in MCF-7 cells.
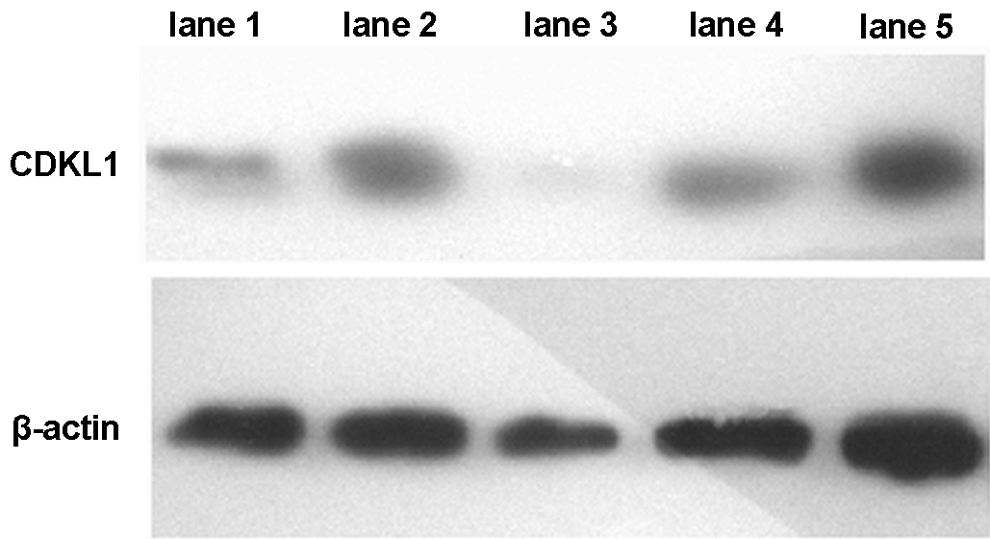
CDKL1 detection by Western blot. Cells interfered with shRNA-709 had the lowest CDKL1 protein expression. Lane 1: shRNA-381; lane 2: shRNA-649; lane 3: shRNA-709; lane 4: shRNA-963; and lane 5: negative control. shRNA, short hairpin RNA.
Silencing CDKL1 affects the cancer cell growth
As shRNA-709 is an effective interfering sequence and can effectively decrease CDKL1 expression in MCF-7 cells, we investigated the effect of the CDKL1 gene silencing on cancer cell growth. The cell cycle was observed by a DNA content analysis (Fig. 4). Compared with the negative control (Fig. 4A), MCF-7 cells interfered with shRNA-709 (Fig. 4B) exhibited obvious accumulation in the G2/M phase (15.329% vs. 5.065%).
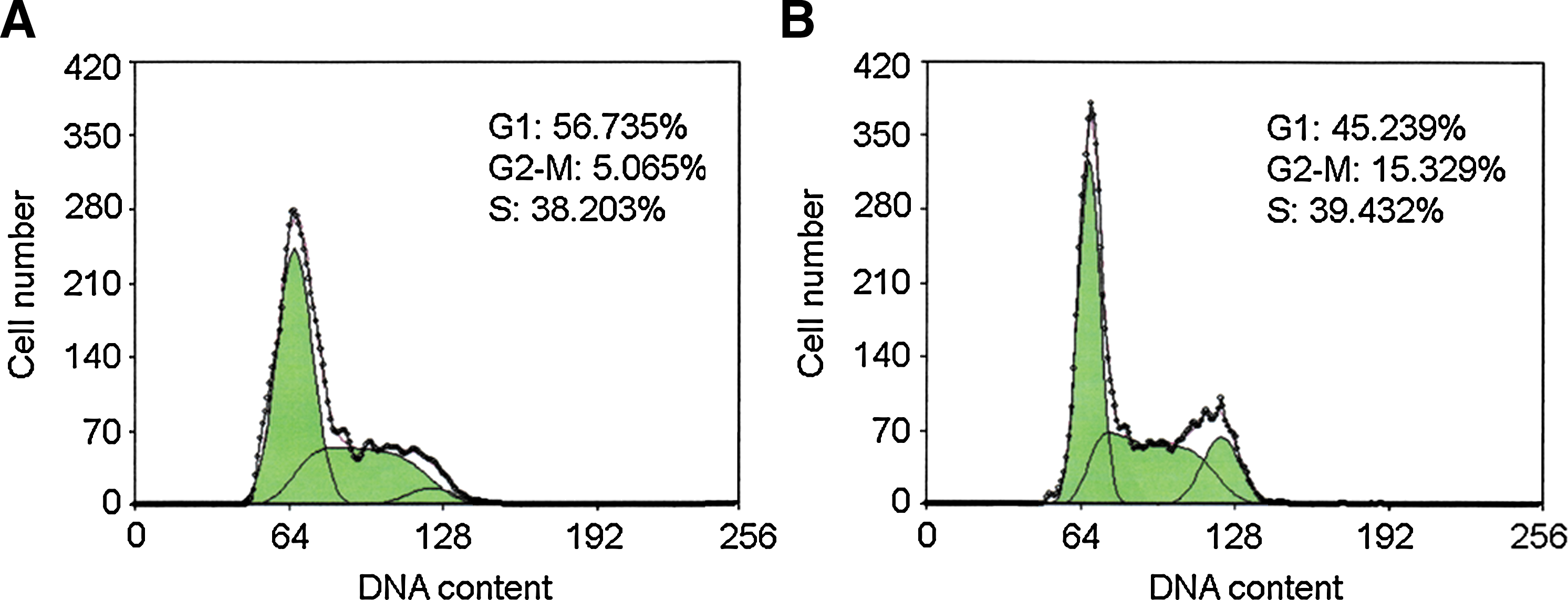
CDKL1 DNA content detected by flow cytometry. Compared with negative control
Silencing CDKL1 increases the sensitivity of breast cancer cells to chemotherapeutic drugs
The MTT assay was carried out to evaluate the relationship between CDKL1 expression and the proliferation of breast tumor cells. As shown in Figure 5, when MCF-7 cells were treated with epirubicin, 5-fluorouracil, and docetaxel for 48 hours, the growth inhibition rates of negative control groups, which transfected with the negative control sequence, were (57.40±3.04)%, (77.88±1.61)%, and (60.71±3.71)%, respectively. Correspondingly, the growth inhibition rates of CDKL1-silencing MCF-7 cells were (76.81±4.16)%, (85.43±1.00)%, and (76.69±0.90)%, respectively (Table 5). Compared with negative control groups, the growth rates of CDKL1-silencing MCF-7 cells decreased significantly when treated with the same cell cycle chemotherapeutic drugs (t=11.73 [p<0.01] for epirubicin, t=9.63 [p<0.01] for 5-fluorouracil, and t=9.32 [p<0.01] for docetaxel).

Histogram of the growth inhibition rates for negative control and shRNA-709-transfected MCF-7 cells. By the Student's t-test, the growth inhibition rates of CDKL1-interfered MCF-7 cell were higher than that of the control group (p<0.01). Error bars represent means±standard deviation for five duplicates.
Breast cancer line MCF-7 was transfected with a plasmid containing either shRNA-709 or a negative control. Epirubicin, 5-fluorouracil, and docetaxel were added at the final concentrations of 1, 25, and 10 μg/mL. Five duplicates were examined for each chemotherapeutic drug.
Discussion
Cancer is a disease of inappropriate cell proliferation, which arises when cells evade normal constraints on their growth or division. 7 –9 Oncogenic transformation often entails derangement of the mechanisms that ensure the stable inheritance of genes and chromosomes during mitotic cell division. 10,11 CDKs play critical roles in both the commitment to cell division and the quality control mechanisms that safeguard genome integrity, and they represent obviously, but potentially risky, therapeutic targets in human cancers. 12,13 Among them, CDC2 is a core molecule on regulation. CDKL1 is a member of the CDC2-related serine–threonine protein kinase family that is likely to occur in malignant tumors, and plays an important impact on tumor progress.
This study showed that CDKL1 is overexpressed in Chinese breast cancer tissue for the first time, and CDKL1 is a more sensitive diagnostic biomarker than ER, PR, P53, and E-cad (Table 3). CDKL1 expression is significantly associated with histology grades of tumors: the higher histology grades or the higher degree of differentiation, the more CDKL1 expression. However, this conclusion needs to be confirmed by further statistical analysis. Our future work will explore the relationship between the expression level and malignant degree.
RNA interference (RNAi)-based therapies have shown great potential for the treatment of diseases such as viral infections, genetic disorders, and cancers. 14 –16 Selection of appropriate gene targets is an important parameter in the potential success of RNAi cancer therapies. Candidate targets include genes associated with cell proliferation, metastasis, angiogenesis, and drug resistance. Importantly, silencing of such genes does not affect the functions of normal cells. 17,18 As different CDKs are specialized to perform discrete functions during distinct cell cycle intervals, 19,20 we investigated whether silencing CDKL1 expressed in breast cancer cells would significantly affect their growth and be sensitive to chemotherapeutic drugs by comparing cell physiology changes before and after interference. We designed and synthesized shRNA-interfering sequences according to the CDKL1 mRNA sequence in NCBI (GeneBank: NM004196). The recombinant shRNA–pGPU/GFP plasmids were sequenced and identified. Real-time RT-PCR and Western blot assay were performed to examine the expression level of CDKL1 mRNA and protein, respectively (Table 4 and Fig. 3). shRNA-709 was indentified to be the most efficient interfering sequence for the subsequent experiments.
It has been known that spatial and temporal sequential activation of CDKs pushes cell progression in the order from the G1 phase through the S phase, G2 phase to M phase, completing cell division, and realizing cell proliferation. CDKs together with their coordinating regulatory subunits cyclins constitute the core mechanism of cell cycle control. The combination of CDK1 and cyclin B1 is responsible for initiating the M phase. 21 In our study, when the CDKL1 function was inhibited, the population of MCF-7 cells in the G2/M phase increased (Fig. 4). This phenomenon prompted that CDKL1 was involved in cell cycle regulation. Beside, downregulation of CDKL1 enhanced the effects of 5-fluorouracil, epirubicin, and docetaxel in MCF-7 cells. In other words, the sensibility of breast cancer cells to chemical therapy increased (Table 5 and Fig. 5). Our work provided the evidence that downregulation of CDKL1 gene expression by shRNA interference is an alternative strategy with potential clinical application in human breast adenocarcinoma. It provided a way to explore a new molecular target for breast cancer clinical therapy.
Conclusions
To summarize, our work verified that the CDKL1 is overexpressed in breast cancer tissue for the first time. CDKL1 has significant high positive detection efficiency compared with ER, PR, P53, and E-cad. The cancer cells treated with shRNA were also more sensitive to chemotherapeutic drugs. As a novel molecule involved in cell cycle regulation, CDKL1 could be not only a more sensitive diagnostic biomarker for breast cancer but also a new gene target for breast cancer therapy.
Footnotes
Acknowledgments
This work was financially supported by the National Basic Research Program of China (2010CB732400), National Natural Science Foundation of China (21075055), Programs for Six Peak Talents (66) and Medicine Leading Talents of Jiangsu Province (LJ201131), and the Youth Project of Jiangsu Cancer Hospital (ZQ200902).
Disclosure Statement
The authors declare that they have no competing interests.