
Abstract
VB6-845 is a recombinant immunotoxin comprised of deBouganin (a de-immunized plant toxin) genetically linked to an epithelial cell adhesion molecule (EpCAM)-targeting humanized Fab fragment (4D5MOCB). EpCAM is highly expressed on a wide range of epithelial tumors but has limited expression on most normal epithelia and therefore represents an excellent target for immunotherapy. A comprehensive preclinical evaluation was performed to determine the safety and suitability of VB6-845 as a systemically administered drug for the treatment of solid tumors. Efficacy studies in mice demonstrated that VB6-845 specifically and potently targeted EpCAM-positive tumors. In a dose-ranging study in Sprague–Dawley rats, single doses of VB6-845 were well-tolerated resulting in a no-observable adverse effect level (NOAEL) of 100 mg/kg whereas repeated doses of VB6-845 resulted in vascular leak-associated symptoms particularly at higher dose levels. However, much higher doses in Cynomolgus monkeys were well-tolerated when given as a 3-hour infusion mimicking the intended route of administration in the clinic. In addition, VB6-845 proved to be minimally immunogenic in monkeys. The toxicological data obtained in Cynomolgus monkeys indicated an excellent safety profile with a NOAEL value of 30 mg/kg (equivalent to a 10 mg/kg dose in humans). These results are supportive of an exploratory Phase I trial.
Introduction
The intent of targeted therapy is to provide significant clinical benefit to the patient while minimizing side-effects related to nonspecific toxicities. Immunotoxins represent one such form of targeted therapy. Basically, immunotoxins are comprised of two functional entities; an antibody moiety that binds to an antigen on the surface of cancer cells, and following internalization, a toxin that blocks translation and inhibits protein synthesis thereby inducing apoptosis. 1 –4 The heightened expression of the target antigen on the cancer cell relative to normal cells provides a higher level of safety not found with chemotherapy. 5 –7 Immunotoxins have been evaluated clinically with some success, particularly in the treatment of patients with hematologic cancers. 8 –10 This success against liquid tumors is likely due to the accessibility of the malignant cells as well as the immunosuppressive nature of these diseases, thus limiting the appearance of dose-limiting, antidrug antibodies (ADAs). 10
Careful antigen selection is critical to the success of an immunotoxin intended for systemic use. The overexpression of epithelial cell adhesion molecule (EpCAM) on carcinomas, combined with its limited accessibility on normal tissue, has made it an ideal target for immunotherapy. 11 –14 In spite of this, several antibody-based therapies targeting EpCAM have met with limited success in the clinic. 15 –20 In the case of immunotoxins, their specificity and potency make them attractive therapeutics; however, impediments, such as immunogenicity, toxicity to the liver and kidneys, and vascular leak syndrome (VLS), have confounded their utility in the clinic. 21 –25 The removal of peptide motifs that induce VLS has served to address some of these issues and reduce toxicity; however, a remaining challenge for immunotoxins in the treatment of solid tumors is to reduce the appearance of neutralizing antibodies that often occurs after only one or two cycles of therapy. 3,26–27
Thus, the foreign nature of immunotoxins restricts their use to certain solid cancers, such as nonmuscle-invasive bladder cancer and squamous cell carcinoma of the head and neck. In the case of VB4-845, an EpCAM-targeting scFv linked to Pseudomonas exotoxin A, loco-regional delivery strategies were used to avoid and/or overwhelm neutralizing antibodies. 15,28 –30 Unfortunately, most solid tumors are not amenable to local administration and developing a successful immunotoxin that is suitable for repeat, systemic administration requires a design that harnesses the potency of these proteins while simultaneously addressing the issue of immunogenicity in order to provide a safe therapy with maximal clinical benefit. 26
Therefore, a prerequisite for repeat systemic delivery using a protein toxin is to remove the immunogenic potential of the payload. To address this, our strategy has been the development of VB6-845, an immunotoxin comprised of a humanized anti-EpCAM Fab fragment fused to a T-cell epitope-depleted form of the plant toxin, bouganin. 31 Bouganin is a type I ribosome inactivating protein (RIP) that potently blocks protein synthesis via deadenylation of rRNA. 32 –34 VB6-845 effectively inhibited tumor growth in multiple tumor xenograft models, including ovarian cancer, without any signs of systemic toxicity. 31 Although T-cell proliferation studies indicated that the epitope-depleted variant of bouganin was non-immunogenic compared with wild type, the true immunogenicity of any immunotoxin can only be evaluated clinically. To take VB6-845 to the clinic, a comprehensive preclinical development strategy was undertaken to determine its safety profile.
In this report we describe the preclinical testing conducted for VB6-845. When considering its safety profile and moderate immunogenic potential, together with its specific and potent anticancer properties, VB6-845 represents an important candidate for clinical investigation.
Materials and Methods
Construct
VB6-845 comprises an anti-EpCAM Fab fragment linked to deBouganin, a T-cell epitope-depleted form of the plant toxin bouganin, derived from Bougainvillea spectabilis Willd (Fig. 1). 31 Briefly, the VB6-845 protein is derived from a dicistronic unit where each chain is fused to a PelB 5′ leader sequence from Erwinia carotavora. The two fragments were generated by Splice Overlapping Extension PCR: PelB-(6xHis)-VH-CH-PelB and VL-CL-deBouganin. The PCR fragments were initially assembled in the pSP73 vector (Promega, Madison, WI) to create the VB6-845 construct, in which deBouganin is linked to the light chain of the anti-EpCAM Fab fragment via a furin-cleavable linker. The construct was subsequently cloned into the pING3302 expression vector containing the arabinose-inducible promoter from Salmonella typhimurium (Xoma, Berkeley, CA) using EcoRI and XhoI restriction enzymes. The final construct was expressed as a soluble protein using Escherichia coli cells (Xoma) as previously described. 31 A six-histidine affinity tag was added to the 5′ end of the variable heavy chain to facilitate purification.
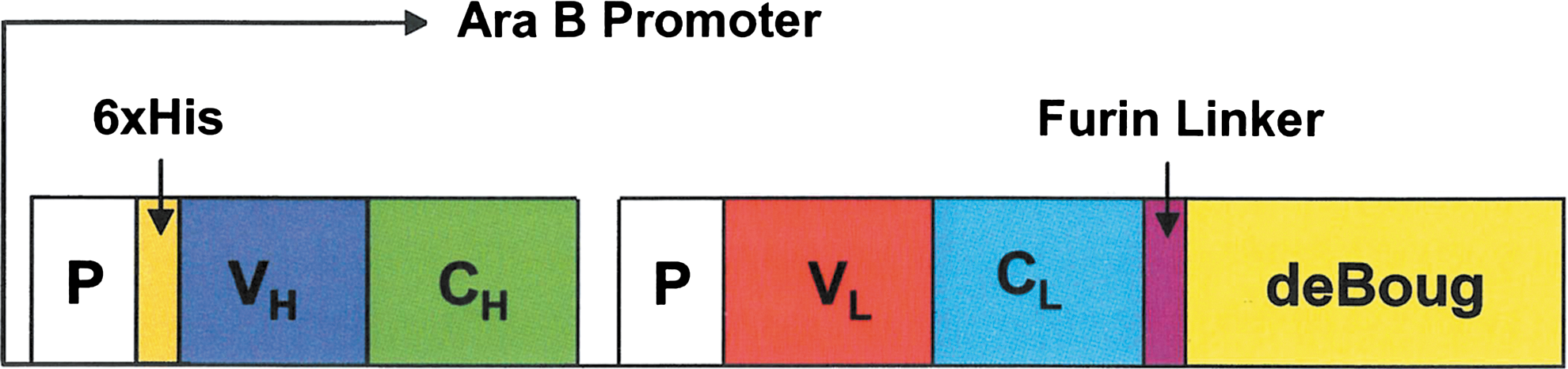
Schematic representation of the VB6-845 construct. VH, VL, CH, and CL correspond to the variable heavy and light chains and constant heavy and kappa chains, respectively; P to the PelB leader sequences; and deBoug to deBouganin. The furin linker and the 6xHis tag, used to facilitate purification, are also indicated.
In vitro specificity and cytotoxicity of VB6-845
Cell lines
All tumor cell lines (American Type Culture Collection, Manassa, VA) and primary normal cell lines (Lonza Walkersville, Inc., Walkersville, MD) were cultured according to specification and maintained at 37°C in a humidified atmosphere containing 5% CO2.
Flow cytometry
The cell surface reactivity of VB6-845 with human normal and tumor epithelial cell lines was assessed by flow cytometry. Briefly, cells (3×105) were incubated with VB6-845 (10 μg/mL) on ice for 2 hours followed by 1 hour on ice with a rabbit polyclonal anti-deBouganin antibody (1:100; Viventia Biotechnologies, Inc., Winnipeg, MB). Binding was detected with fluorescein isothiocyanate-conjugated goat antirabbit IgG (1:100; The Binding Site, San Diego, CA). Cells were resuspended in PBS/FBS buffer containing 0.6 μg/mL propidium iodide (Molecular Probes, Eugene, OR), and binding was analyzed with an FACSCalibur flow cytometer (Becton Dickinson, Franklin Lakes, NJ). The EpCAM-negative melanoma tumor cell line (A-375) was included as a negative control. Binding was considered positive if antibody-treated cells exhibited a positive shift in fluorescence of >30% over a PBS control.
Cell potency assay
The ability of VB6-845 to inhibit cell growth was determined using a standard MTS [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt] assay (Promega, Madison, WI). Briefly, tumor and normal cells (1000 cells/well) in culture medium containing 10% FBS were incubated with VB6-845 over a range of concentrations (0.005–500 nM) for 5 days at 37°C in the presence of 5% CO2. Cell proliferation was determined as specified by the manufacturer and an IC50 value was calculated for each cell line.
Immunohistochemical staining with VB6-845
In accordance with FDA regulations, the reactivity of VB6-845 with normal human tissues was investigated using immunohistochemical (IHC) analysis. 35 A panel of 35 normal human tissues (Genzyme Genetics Analytical Services, Los Angeles, CA) was incubated with 1.25 μg/mL VB6-845 (or negative reagent control) for 30 minutes at room temperature (RT), and blocked and incubated with the secondary antibody rabbit polyclonal anti-deBouganin (Viventia Biotechnologies, Inc.) at 6.25 μg/mL for 30 minutes at RT. Labeled Polymer Horse Radish Peroxidase (HRP) (DakoCytomation, Burlington, ON) was added for 30 minutes at RT and the reaction was visualized by 3,3′-diaminobenzidine tetrahydrochloride (DAB) solution (DakoCytomation). Samples were counterstained with a modified Harris hematoxylin (American Master Tech. Scientific, Inc., Modesto, CA). The degree of immunoreactivity of VB6-845 with normal human tissue was determined by diagnostic pathology. Scoring intensity was based on a semiquantitative system with values ranging from 0 to 3+.
In vivo efficacy
Female HRLN nu/nu mice bearing subcutaneous MCF-7 human tumor xenografts were used to evaluate the in vivo efficacy and tolerability of VB6-845. On day 1 of the study, 1×107 MCF-7 cells were implanted subcutaneously in the right flank of animals 11–12 weeks old and weighing ∼20 g. Once tumor volume had reached the required size, the animals were pair-matched into two groups (n=12), each with a mean tumor volume of 112 mm3. Group 1 mice were untreated tumor growth controls. Group 2 mice received 20 mg/kg of VB6-845 administered by intravenous (i.v.) bolus injection via the lateral tail vein for five consecutive days, followed by 2 days off treatment, for three cycles. Each dose was administered in a volume of 0.2 mL per 20 g body weight (10 mL/kg), scaled to the body weight of the animal.
Selection of an animal model
To establish a pharmacologically relevant species, several animal species (mouse, rat, dog, and monkey) were screened for binding cross-reactivity with VB6-845 through IHC analysis. VB6-845 did not exhibit cross-reactivity against a select panel of tissues from monkey, dog, rat, or mouse. Based on these results, it was determined that VB6-845 would not be pharmacologically active in terms of EpCAM binding in species typically used to conduct toxicology studies. Consequently, pharmacokinetic and toxicologic studies were conducted in two animal models: Sprague–Dawley rats because of their sensitivity to toxins and Cynomolgus monkeys, a nonhuman primate model.
Animal toxicology studies
Sprague–Dawley rats and Cynomolgus monkeys were obtained from Charles River Canada (Montreal, PQ) and MPI Research (Mattawan, MI), respectively. Animals were housed and maintained according to AAALAC International Guide for the Care and Use of Experimental Animals. Rats were fed commercial rodent chow (PMI Certified Rodent #5002; PMI Nutrition International, Inc., Richmond, VA), monkeys with commercial primate food (Lab Diet®, Certified Primate Diet #5048; PMI Nutrition International, Inc.) and other supplements.
Single-dose toxicology in Sprague–Dawley rats
In an initial dose-ranging study, Sprague–Dawley rats (3/sex/dose) were administered VB6-845 at doses of 6.25, 25, or 50 mg/kg, followed by a 2-week observation period. Then, a definitive single-dose study was performed, in which Sprague–Dawley rats (three males/group) were given 100 or 200 mg/kg followed by a 2-week observation period. In both studies, VB6-845 diluted in vehicle (20 mM Tris and 500 mM NaCl [pH 7.4]) or vehicle alone was administered as a single i.v. bolus via the lateral tail vein. Parameters examined were clinical signs, body weight, hematology, and serum chemistry.
Repeated-dose toxicology in Sprague–Dawley rats
A repeated-dose GLP study was conducted in which Sprague–Dawley rats were administered 10, 20, or 40 mg/kg of VB6-845 or vehicle alone, every third day for 2 weeks, followed by a 2-week recovery period. The Main Study group consisted of 20 animals (10/sex/group) with Recovery groups included at the two upper doses (5/sex/group). The Control group received vehicle alone and consisted of 20 animals (10/sex) and a Recovery group of 10 animals (5/sex). The parameters monitored during the study were mortality, clinical signs, body weight, food consumption, ophthalmology, urinalysis, hematology, and organ weights as well as macroscopic and microscopic evaluation.
Pharmacokinetics
The pharmacokinetic profile was evaluated as part of the repeated-dose study using a sandwich enzyme-linked immunosorbent assay (ELISA). Plasma samples were collected at eight sampling time points (0, 0.25, 0.5, 1, 2, 4, 6, and 24 hours) on days 1 and 13 from three animals/sex/dose.
Briefly, ELISA plates were coated with affinity-purified rabbit anti-deBouganin and incubated overnight at 4°C. After a blocking step, plasma samples were incubated for 2 hours at 22°C and biotin-conjugated affinity-purified rabbit anti-deBouganin detector antibody (Viventia Biotechnologies, Inc.) added for 1 hour (22°C) followed by streptavidin-HRP (Pierce, Rockford, IL) for 30 minutes at 22°C. Subsequently, tetramethylbenzidine substrate (KPL, Gaithersburg, MD) was added and the absorbance was read at 450 nm (Vmax ; Molecular Devices Corp., Sunnyvale, CA). The concentration of VB6-845 was interpolated from the standard curve (SOFT max ; Molecular Devices Corp.). Normal rat plasma (Biomeda, Foster City, CA) was used as a negative control and standard curves were prepared by spiking VB6-845 (0–50 ng/mL) into the plasma.
Repeated-dose toxicology in Cynomolgus monkeys
A GLP study was conducted to evaluate the systemic effects of repeated doses of VB6-845 administered via a 3-hour i.v. infusion in Cynomolgus monkeys at a dose volume of 10 mL/kg/hour. In all, four treatment groups of 2/sex/group were administered VB6-845 at 10, 30, 60, or 90 mg/kg on days 1 and 8. In addition, two treatment groups of 1/sex/group were administered 60 or 90 mg/kg of VB6-845 and were maintained for a 20-day recovery period. Parameters monitored during the study included clinical signs, mortality, body weight, and food consumption. Samples for hematology evaluations were collected from all animals pretest and on day 7, and additional samples were collected from Recovery animals on days 14, 21, and 28. Samples for clinical chemistry analysis were collected from all animals pretest and on days 3 and 7 with additional samples being collected from the Recovery animals on days 2, 9, 10, 14, 17, 20, 23, and 26. All animals were necropsied at the end of the study and recovery periods at which time organs were weighed and a histopathological analysis was performed.
Pharmacokinetics
The pharmacokinetic profile of VB6-845, following i.v. administration, was evaluated as part of the repeated-dose study using an ELISA. Plasma samples for pharmacokinetic analysis were obtained from monkeys (2/sex/group) at time 0 and 0.5, 1, 2, 4, and 24 hours after dosing with VB6-845. The analysis was performed as described for the pharmacokinetic analysis in the repeated-dose rat study, except that commercial monkey plasma (Biomeda) was used as a negative control.
Immunogenicity
Monkey plasma samples were taken from the Main Study animals on days 0 and 7 (2/sex/dose cohort), and from the Recovery group animals on days 0, 7, 14, 21, and 28 (1/sex/dose cohort). Samples were analyzed by ELISA to determine the presence of antibodies against the Fab and deBouganin moieties. Briefly, for all samples, ELISA plates were coated with either the Fab or deBouganin, in coating buffer, at equimolar concentrations and incubated overnight at 4°C. After washing and incubation with blocking buffer, serially diluted monkey plasma was added and incubated for 2 hours at 22°C. Rat and monkey pretreatment samples were used as negative controls. HRP-conjugated goat antimonkey IgG, IgM, and IgA (H+L) secondary antibodies (Rockland Immunochemicals, Gilbertsville, PA) were added for 1 hour and incubated at 22°C. After washing, TMB substrate was added, and an absorbance was read at 450 nm (V max ; Molecular Devices Corp.). Titre is defined as the reciprocal of the last dilution with a positive OD450 (>0.1) as determined from 30 independent assays of the diluent alone.
Results
Immunohistochemistry
The pattern of VB6-845 reactivity observed in this study is consistent with the reported expression of EpCAM in normal human tissue. 36 –39 Of the 35 tissues examined, varying degrees of binding were noted in 20 tissue types; binding was mainly membrane associated with some cytoplasmic staining (data not shown). No nuclear staining was observed in any of the tissue specimens. While the majority of tissues showed some degree of cell membrane positivity, the tissues showing the strongest binding (3+) were colon, fallopian tube, gall bladder, pancreas, parathyroid, small intestine, and uterus. Moderate binding (2+) was observed in breast, pituitary, prostate, thyroid, trachea, and ureter, while the lowest intensity staining (1+) was observed in bladder, kidney, lung, ovary, stomach, testis, and thymus. No cell surface binding was detected on white blood cells or nonepithelial tissues: bone marrow, cerebellum, cerebral cortex, eye, heart, liver, lymph node, muscle, placenta, skin, spinal cord, spleen, and tonsil. In addition, no positive staining was observed in stromal components, such as endothelium, smooth muscle, and fibroblast.
In vitro specificity and cytotoxicity of VB6-845
To illustrate the specificity and potency of VB6-845, a panel of epithelial tumor and normal cell lines was tested for cell surface reactivity and cytotoxicity (Table 1). The cell surface reactivity and corresponding potency of VB6-845 varied according to EpCAM expression. In general, high EpCAM expression was associated with high potency. Low EpCAM-expressing or EpCAM-negative cell lines, such as the normal endothelial (HMVEC-bd), normal breast (HMEC), and melanoma (A-375) cell lines, had no measurable cytotoxicity, indicating the specificity of VB6-845 for EpCAM-expressing cells. MCF-7 cells showed high levels of cell surface reactivity and associated cytotoxicity; therefore, an MCF-7 mouse xenograft model was chosen for follow-up efficacy studies.
Mean fold increase in median fluorescence±SEM over PBS control.
Cell lines from American Type Culture Collection (ATCC) (Manassas, VA).
Cell lines from Lonza Walkersville, Inc. (Walkersville, MD).
In vivo efficacy
All MCF-7 tumor-bearing mice (12/12) in the untreated group reached the 1000 mm3 endpoint tumor volume (100% mortality) by day 43 with a median time to endpoint (TTE) of 20.4 days (Fig. 2A, B). In comparison, VB6-845-treated mice exhibited a 111% tumor growth delay (TGD), defined as the percent increase in median TTE of treated versus control mice (p<0.001), with a 70% survival rate over the study period. The median tumor volume in the treatment group was 148 mm3 at the end of the study. Three animals achieved a complete response, defined as complete tumor regression with no measurable mass (<13.5 mm3) for three consecutive measurements during the course of the study; one of these animals had no measurable tumor from day 15 through to the termination of the study. One additional animal was a partial responder, defined as having three consecutive measurements where the tumor size was ≤50% of the size on day 1, but ≥13.5 mm3 for one or more of those measurements. There were two unexpected deaths that could not be attributed to treatment and were therefore excluded from the analysis. No significant weight loss was observed over the course of the treatment, indicating that VB6-845 was well tolerated.

Mean tumor growth curves and survival plots.
Animal toxicology studies
Single-dose toxicology in Sprague–Dawley rats
No treatment-related changes occurred in the initial single-dose range finding study when animals were dosed at 6.25, 25, or 50 mg/kg or during the following 2-week observation period. All clinical signs, body weight, hematology, serum chemistry (after 2 weeks), and macroscopic observations at necropsy were within the normal range. Based on these results, a maximum tolerated dose (MTD) was not determined and a second acute dose range finding study was conducted in which three males per group were dosed with vehicle alone, 100, or 200 mg/kg VB6-845.
No unexpected deaths occurred and there were no adverse clinical signs in either the untreated control group or animals dosed up to and including 100 mg/kg. However, rats administered 200 mg/kg demonstrated adverse clinical signs, including edema of the forepaws and a slight decrease in activity level. Rats in both the 100 and 200 mg/kg dose groups showed less weight gain when compared with the control group. Dose-dependent increases in alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels followed the administration of VB6-845 (Fig. 3A, B). On day 3, AST levels increased 5- and 16-fold whereas ALT increased 6- and 38-fold over controls at 100 and 200 mg/kg, respectively. In both treatment groups, ALT returned to control levels by day 8, and AST by day 15. After 2 weeks of recovery, all clinical signs, hematology, clinical chemistry, and urinalysis were within normal range. Immunohistochemistry conducted on livers at the end of the study indicated no signs of toxicity. The elevated AST and ALT levels, in conjunction with the paw edema, determined an MTD of 200 mg/kg. The no-observable adverse effect level (NOAEL) for VB6-845 was concluded to be 100 mg/kg.
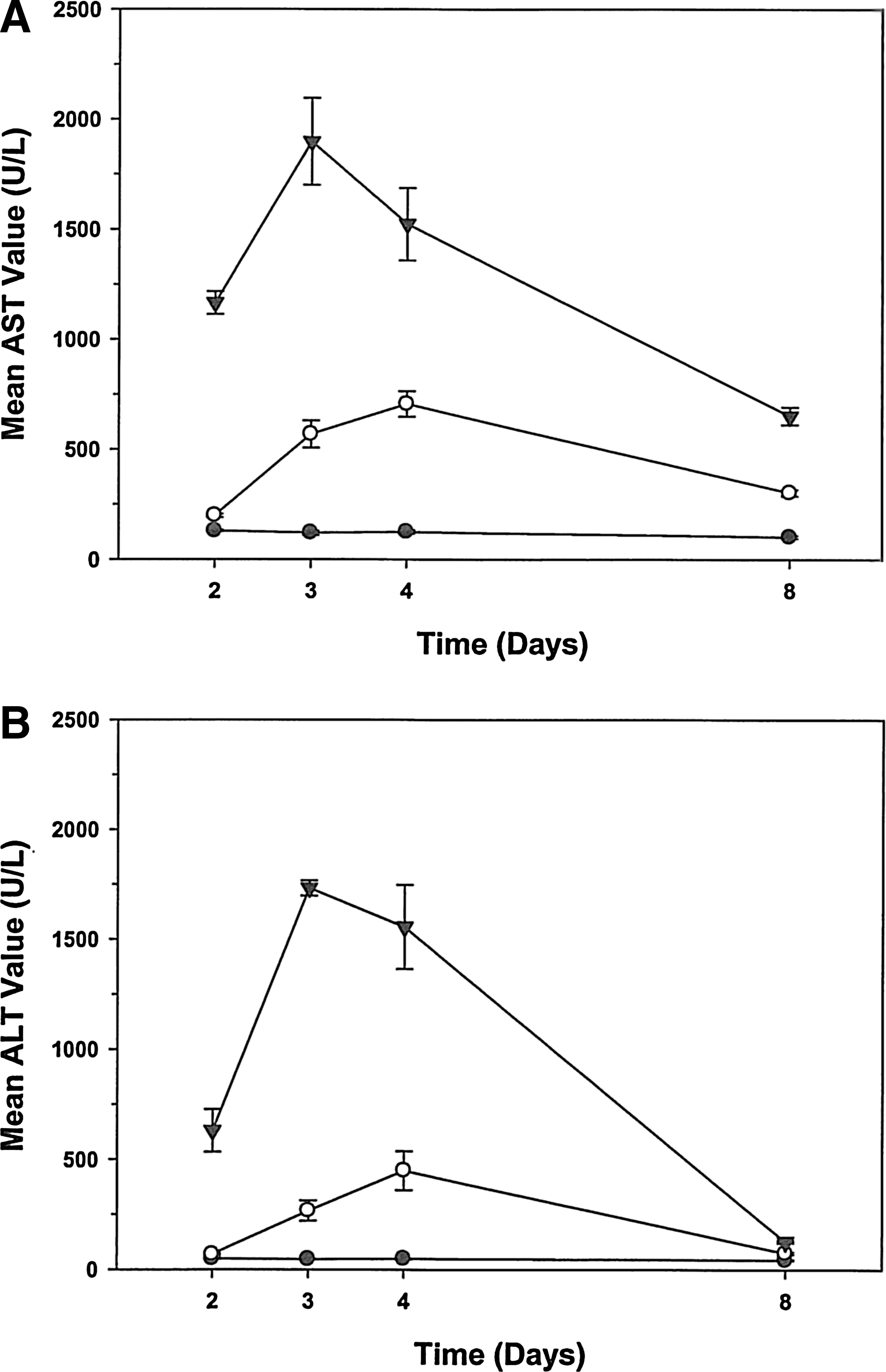
Comparison of liver enzyme concentrations in plasma
Repeated-dose toxicology in Sprague–Dawley rats
All animals tolerated three doses of VB6-845 at 10, 20, and 40 mg/kg. Clinical signs appeared after the fourth or fifth dose that were dose dependent and included cold to touch, decreased activity, labored breathing, and cyanosis ranging from mild to severe. There was a dose-dependent decrease in body weight, body weight gains, and food consumption during the treatment period (data not shown). Food consumption returned to normal by the third week and body weight gains were similar to the control groups by the fourth week.
Some clinical signs, such as the appearance of cyanosis after the fourth or fifth dose, were indicative of an allergic response to VB6-845. These observations are consistent with anaphylaxis experienced in rats. 40 Of the 20 animals dosed at 10 mg/kg, four developed cyanosis after the fifth dose with no incidence of mortality. Thirty animals each received doses of either 20 or 40 mg/kg. Twenty-one rats experienced cyanosis after the fourth or fifth dose at 20 mg/kg (11/21 survived). Twenty-four rats dosed at 40 mg/kg experienced cyanosis after the fourth dose (11/24 survived). Due to toxicity, animals dosed at 40 mg/kg did not receive the fifth dose. In all, thirty animals were found dead or were euthanized preterminally at 20 (12/30) and 40 (18/30) mg/(kg·day−1). Minimal-to-slight heart lesions were noted in all animals that were found dead and all had macroscopic evidence of gastrointestinal (GI) tract alterations, such as hemorrhaging and congestion, again consistent with an anaphylactic response in rats. 40,41 The most common organ to be affected was the heart that showed minimal-to-marked myocardial degeneration with concurrent minimal-to-slight fibrosis. Examination of animals in the Recovery group, dosed at 20 and 40 mg/kg, indicated minimal-to-moderate heart fibrosis, a low incidence of alveolar histiocytosis, and perivascular mixed cell inflammatory infiltrate, indicative of an ongoing recovery process. All other treatment-related occurrences noted in the Main Study animals were absent from the Recovery animals, indicating side-effects were reversible and resolved on their own. A NOAEL was not determined as toxicity was observed at all dose levels.
Hematology and clinical chemistry
Increases in group mean platelet count (1.5- to 1.7-fold over controls) were noted in animals dosed at 40 mg/kg (Supplementary Data, Table S1; Supplementary Data are available online at
Pharmacokinetics
Plasma concentrations of VB6-845 were similar among animals of each group at most blood collection time points on day 1; however, there was a higher inter-individual variability within groups on day 13. VB6-845 was not quantifiable in any samples collected predose on day 1 or 13. There was no apparent difference in concentration levels between male and female rats. The estimated composite terminal half-lives ranged from 3.74 to 4.36 hours on day 1 and 1.14 to 1.30 hours on day 13 (Supplementary Data, Table S2). The volume of distribution (Vd ) was higher than the blood volume in rats (64 mL/kg), suggesting a quick initial equilibration in rat tissue; Cmax values increased as a function of dose on both occasions. Peak plasma concentrations of VB6-845 ranged from 84.0 to 408 μg/mL on day 1 and from 68.0 to 176 μg/mL on day 13.
Repeated-dose toxicology in Cynomolgus monkeys
Hematology and clinical chemistry
There were no mortalities or changes in body weight that could be attributed to the administration of VB6-845. Treatment-related clinical signs were limited to decreased activity and hunched posture following the second treatment at all dose levels; there was no indication of a dose relationship and all clinical signs generally resolved the following day. On day 7, red cell parameters were comparably decreased in all groups, with the exception of reticulocytes that were elevated, although remained within normal range (Table 2). These decreases were maintained in the Recovery animals on day 14, and were suggestive of a test article-related effect. Increased reticulocytes on day 14 were compatible with a delayed bone marrow regenerative response.
Baseline clinical hematology and serum chemistry data were established based upon a historical database collected by MPI Research.
Values reported are mean±SD.
Outside of normal expected range.
Data for males and females are combined. All animals were examined twice daily for mortality, morbidity, and the availability of food and water. Clinical observations were conducted daily beginning on day 2.
Total parameters examined: erythrocyte indices (MCV, MCH, MCHC, and RDW), hematocrit, hemoglobin, mean platelet volume, platelet count, red blood cell count, reticulocyte count (absolute and percent), and white blood cell count (total, absolute, and percent differential). Samples for hematology evaluations were collected from all animals pretest and on day 7. Additional samples were collected from Recovery animals on day 14.
Total parameters measured were as follows: A/G ratio (calculated), ALT, albumin, alkaline phosphatase, AST, blood urea nitrogen, calcium, chloride, cholesterol, creatinine, gamma-glutamyl transferase, globulin, glucose, phosphorus, potassium, SDH, sodium, total bilirubin, and total protein. Samples for clinical chemistry evaluations were collected from all animals pretest and on day 7. Additional samples were collected from Recovery animals on days 2, 9, 10, 14, 17, 20, 23, and 26.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; SDH, sorbitol dehydrogenase; NS, not sampled.
Transient changes in liver enzymes were observed throughout the treatment (AST, ALT, and sorbitol dehydrogenase [SDH]) and recovery periods (AST and ALT) (Table 2). These liver enzyme changes did not appear to be dose dependent; however, the frequency of microscopic changes observed in the liver did tend to be dose dependent (Table 3). At the end of the treatment period (i.e., day 9) dose-dependent gross necropsy changes, including pale discoloration of the kidneys at 90 mg/kg (males only), mild enlargement of the spleen at 60 to 90 mg/kg (females only), and pale discoloration of the liver at 30 to 60 mg/kg (males and females, respectively), were observed (Table 3). All changes in liver enzymes had resolved by day 20 in the Recovery group and none of the gross necropsy changes observed on day 9 were present.
Treatment groups consisted of two males and two females per group. Recovery group consisted of one male and one female per group. Values reported as M/F.
NA, not applicable.
Overall, the changes associated with the administration of VB6-845 during the treatment period were resolved by the end of the recovery period, except for the tubular regeneration in the kidneys, which was still ongoing in the male at 60 mg/kg and in the female at 90 mg/kg. Based on the histopathological changes seen at 60 and 90 mg/kg, the NOAEL was determined to be 30 mg/kg and the NOEL was determined to be 10 mg/kg.
Pharmacokinetics in Cynomolgus monkeys
The pharmacokinetic parameters of VB6-845 were evaluated as part of the repeated-dose toxicology study. Plasma sample analysis indicated a proportional relationship between dose and pharmacokinetic parameters (mean peak exposure [Cmax ] and the extent of exposure [AUC(0-Tlast)] in samples collected on both days 1 and 8) (Fig. 4; Table 4). The mean half-life (T 1/2) values of VB6-845 on days 1 and 8 were 2.5±0.1 and 2.4±0.4 hours, respectively. Some variability was observed between males and females; however, a statistical difference could not be calculated due to the limited number of monkeys in each dosing group. The Cmax on day 8 was consistently lower than on day 1, with concomitant decreases in clearance and volume distribution (Vd ) (Table 4).
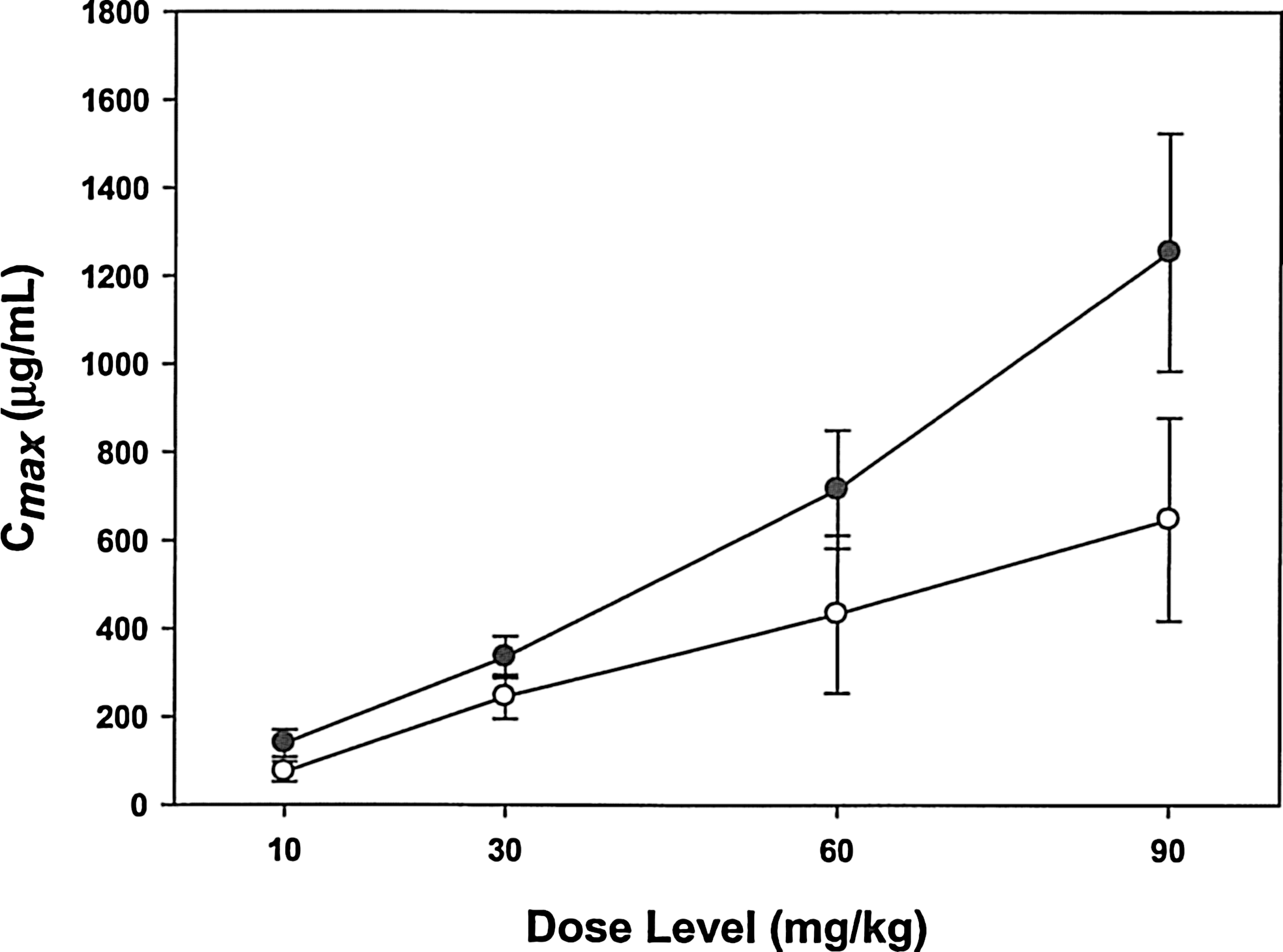
Comparison of mean Cmax values and VB6-845 dose levels for days 1 (●) and 8 (○) in Cynomolgus monkeys.
Data for males and females are combined.
Values are mean±SD, n=4.
Cmax , maximum observed drug concentration in plasma; AUC (0-Tlast) , area under the drug concentration–time curve from time 0 to time t, where t is the time of the last measurable plasma concentration; T 1/2, elimination half-life; Cl, apparent plasma clearance; Vd , apparent volume of distribution.
Immunogenicity in Cynomolgus monkeys
No immune response was observed against either the humanized Fab fragment or the toxin moiety in the prebleed samples taken prior to dosing or in any animals on day 7; however, antibody responses were detected by day 14 in the Recovery group animals. The reactivity to deBouganin was 3-fold lower than to the Fab fragment (Fig. 5).
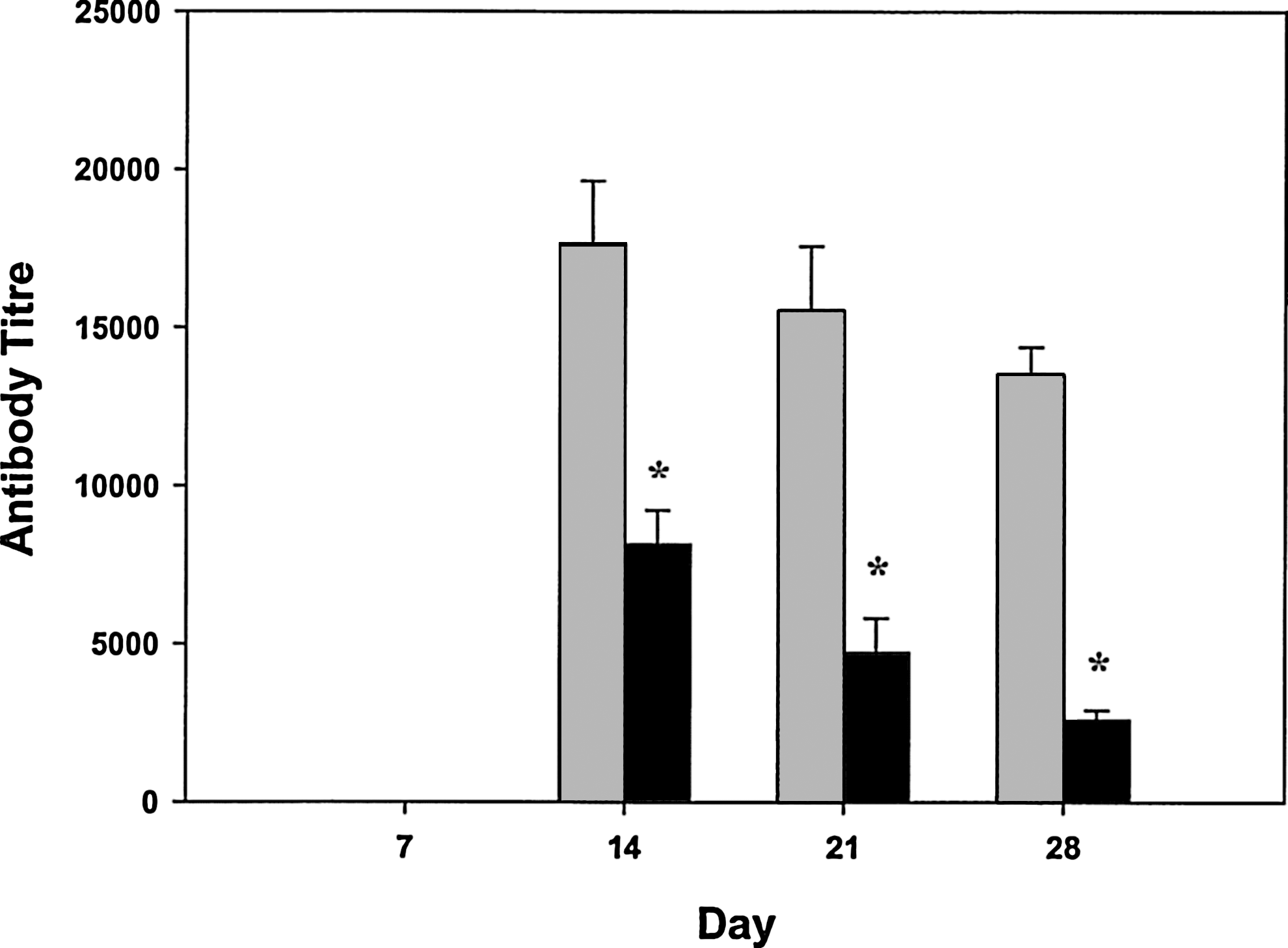
Antibody response to Fab and deBouganin in Cynomolgus monkeys (Recovery group) following i.v. injection of 90 mg/kg on day 1 and day 8. Values for both genders were combined (n=2, 1M and 1F). Fab, gray; deBouganin, black. Mean antibody titres±SD, n=2. *Analysis by Student's t-test indicates that there is a statistically significant difference between the means (p=0.007).
Discussion
The specificity and potency of immunotoxins offer the potential for safer and more efficacious options for the treatment of solid tumors. However, their limitations in the clinic have included toxicity to non-target cells, VLS, and the appearance of ADAs, elicited primarily against the toxin moiety. Careful choice of target antigen to ensure minimal normal tissue reactivity, as well as genetic manipulations to remove VLS-associated domains and motifs have reduced many of the toxicities. 26,42,43 However, as most solid tumors require multiple treatment cycles, it is critical that ADA responses be minimized in order to provide prolonged clinical benefit. In this regard, multiple strategies, including PEGylation and co-administration of immunosuppressant drugs as well as genetic mutation to remove B and T-cell epitopes, have been developed to suppress the patient immune response resulting in varying degrees of success. 44 –49
In this study we performed a comprehensive preclinical assessment of VB6-845, an immunotoxin intended for the systemic treatment of EpCAM-positive tumors of epithelial origin. EpCAM represents an excellent therapeutic target as it is overexpressed on the surface of a wide range of epithelial tumors but has limited expression, mostly confined to the basolateral surface, on normal epithelia. 12 –14 IHC analysis of VB6-845 demonstrated reactivity with 20 tissue types; the binding pattern was membrane associated and comparable to that seen with other EpCAM-reactive antibodies. 12,36,37,39,50,51
To satisfy regulatory requirements for a first-in-man study, the preclinical strategy for VB6-845 followed ICH S6 guidelines. 52 This comprised a stepwise evaluation that first determined the specificity and potency of VB6-845 against tumor cells as well as its immunoreactivity with critical normal human tissue to establish its potential for cross-reactivity. Following the demonstration of in vivo efficacy in a murine model, a comprehensive safety evaluation was performed in the Sprague–Dawley rat and Cynomolgus monkey. To select an animal model, IHC reactivity was examined in several animal species typically used for toxicology studies; however, no cross-reactivity was observed. Given the absence of a pharmacologically relevant animal model, the preclinical safety evaluation was performed in both Sprague–Dawley rats and Cynomolgus monkeys. The effects of immunotoxins in Sprague–Dawley rats are well-documented and it was therefore chosen as the initial model for toxicological examination. 53 In the Cynomolgus monkey, the route of administration and the treatment regimen used during the toxicological study mimicked those to be used in the clinic. A compilation of the in vitro and in vivo data was used to establish the specificity, potency, tolerability, and safety of VB6-845 and determine whether it was a suitable candidate for a Phase 1 trial.
In efficacy studies, VB6-845 administered at 20 mg/kg proved to be efficacious and well-tolerated in mice bearing MCF-7 human tumor xenografts. The excellent tolerability of deBouganin is in agreement with previous studies in which 32 mg/kg was not lethal for any animals, making bouganin the least toxic of the type I RIPs. 32 In comparison, pokeweed antiviral proteins, saporin and gelonin, have LD50 values of 0.9–2.6, 4.0–8.1, and 40 mg/kg, respectively. 54 Further, immunoconjugates comprised of anti-CD80 and anti-CD86 IgGs linked to bouganin, saporin, or gelonin all demonstrated specificity and potency to target cells; however, a considerably lower toxicity was observed for the bouganin conjugates, making this type I RIP a preferred toxin for clinical applications. 34
No mortality or serious toxic effects were observed following single i.v. doses of VB6-845 to Sprague–Dawley rats and clinical signs were limited to slight reddening of the skin and paw edema, as well as elevated AST and ALT levels that returned to normal range post-treatment. In the repeated-dose study, animals tolerated at least three administrations; however, dose-dependent clinical signs were observed following the fourth or fifth treatment that included decreased activity, labored breathing, and cyanosis. Symptoms of anaphylaxis were noted at all dose levels and included intestinal lesions, platelet aggregates, and paw edema. 40,55,56 The occurrence of these symptoms, together with the hypoalbuminemia and increased platelet counts, is suggestive of a systemic inflammatory response. 27 Previous studies with Sprague–Dawley rats have described the sensitivity of this model to anaphylaxis and VLS-like symptoms following the administration of immunotoxins and therefore these results were not unexpected. 15,41,53,57 However, as observed in this study, the VLS-related symptoms were generally limited to the rat and not observed in other species. 41,58 In surviving animals, changes in liver enzymes and albumin/globulin ratios were transient and returned to normal physiological levels in Recovery group animals. Similarly, microscopic changes in the heart and GI tract were resolved in the Recovery group with only minimal levels of fibrosis remaining in the heart.
Given the toxicity observed in Sprague–Dawley rats, a non-human primate model was chosen for a definitive toxicological study. In Cynomolgus monkeys, VB6-845 was administered as a 3-hour i.v. infusion, mimicking the intended dosing regimen in the clinic, with two doses given 1 week apart. There were no mortalities reported at any of the dose levels, and clinical signs were reversible and limited to decreased activity and hunched posture with no apparent dose relationship. Changes in liver enzymes (AST, ALT, and SDH) were observed during the treatment period that did not appear to be dose dependent. In addition, necrotic foci and sub-acute inflammation were noted that appeared to be dose dependent; however, these microscopic variations and all liver enzyme changes had resolved by the end of the recovery period. The anaphylactic response was limited to the rat and was not seen in Cynomolgus monkeys, even at higher dose levels.
The immunogenicity of VB6-845 was investigated in the repeated-dose study in monkeys, and as VB6-845 was a foreign protein an immune response was expected. Antibody titres were observed by day 14 and the immune response was likely reflected in the decrease in Cmax values observed between day 1 and day 8 at all dose levels. Such a response would result in VB6-845 being partially cleared from the circulation resulting in lower plasma concentrations, and given that the T 1/2 values on day 1 and day 8 were under 3 hours, a full clearance of VB6-845 would be expected between weekly doses. Although larger immunotoxins may prolong drug exposure, the reduced size of the Fab-toxin format is preferred for facilitating tumor penetration as well as reducing the potential for off target toxicities. 59
Interestingly, the immune response to deBouganin was very minimal with the major response being directed against the Fab moiety. The low immunogenic potential of deBouganin in Cynomolgus monkeys is in direct contrast to the high antibody titre generated against another type I RIP with similar sequence homology, gelonin, linked to a CD-2-specific antibody. 60 Although the results reported here suggest that deBouganin is minimally immunogenic compared with other type I toxins, a true evaluation of immunogenic potential can only be obtained from patient samples following repeat administration.
The specificity, potency, and safety profile of VB6-845 suggest that it has the potential to be an effective and safe therapy for the treatment of EpCAM-positive tumors. The high dose level achieved in Cynomolgus monkeys implies that VB6-845 has a significant therapeutic window as toxicological effects were only observed at a 1500-fold greater value than the IC50 value for tumor cells. Further, the toxicological data in monkeys indicated an excellent safety profile with a NOAEL value of 30 mg/kg, which is a six-fold higher dose than that required for efficacy in a murine model. 61 The immunogenicity data suggest a minimal response to the toxin moiety that is a key parameter for clinical success using an immunotoxin for repeated systemic therapy.
Taken together, the preclinical results support the clinical development of VB6-845 for the systemic treatment of EpCAM-positive tumors. A NOAEL of 30 mg/kg in Cynomolgus monkeys approximates to a 10 mg/kg dose in humans and applying the generally accepted 1 log lower safety margin would result in the safe starting dose for humans being set at 1 mg/kg. 61,62 Subsequent to this study, a Phase 1 exploratory trial was initiated with a starting dose of 1 mg/kg, administered as a 3-hour i.v. monotherapy infusion, given once weekly.
Footnotes
Disclosure Statement
Joycelyn Entwistle, Shilpa Chooniedass, Jeannick Cizeau, and Glen C. MacDonald are employees of Viventia Biotechnologies, Inc. which has direct financial interest in the subject matter discussed in this manuscript.
Acknowledgments
The authors gratefully acknowledge Shauna Stevenson for contributing to the preparation and review of the manuscript.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.