
Abstract
Chemotherapy with platinum doublets, including cisplatin plus vinorelbine, is standard of care for non–small-cell lung cancer. Sublethal exposure to certain chemotherapeutic agents has been demonstrated to alter the phenotype or biology of human tumor cells, rendering them more susceptible to cytotoxic T lymphocyte (CTL)–mediated lysis. The effects of cisplatin/vinorelbine on tumor sensitivity to T-cell cytotoxicity and its molecular mechanisms, however, have not been fully elucidated. We examined the effect of this chemotherapy on growth, cell-surface phenotype, and CTL-mediated lysis of five distinct human lung carcinoma cell lines in vitro and examined the molecular mechanisms associated with enhanced CTL sensitivity. These studies demonstrate that sublethal exposure of human lung tumor cells to the platinum doublet modulates tumor cell phenotype and increases sensitivity to major histocompatibility complex–restricted perforin/granzyme–mediated CTL killing. These studies also demonstrate that exposure to chemotherapy markedly decreased the protein secretion ratio of transforming growth factor-β/interleukin (IL)-8. We examined the gene expression profile of two lung tumor cell lines to identify a shared gene signature in response to sublethal cisplatin/vinorelbine and found coordinate expression of only 16 transcripts, including those for cytokine/chemokine expression and apoptosis such as tumor necrosis factor-α, IL8, CXCL5, and B cell lymphoma-2–like genes (BCL-2). Overall, these results suggest that sublethal exposure to cisplatin/vinorelbine increases sensitivity to perforin/granzyme–mediated CTL killing by modulation of (a) tumor phenotype, (b) cytokine/chemokine milieu, and (c) the proapoptotic/antiapoptotic gene ratio. The data presented here propose a complex mechanism that is distinct from and complementary to that of immunogenic cell death. This molecular signature may be useful in predicting responses to immunotherapy as well as provide the rationale for the potential clinical benefit of the combined use of vaccine with cisplatin/vinorelbine regimens.
Introduction
Lung cancer is the leading cause of cancer death worldwide. 1 Standard-of-care therapies for these patients include a combination of surgery, chemotherapy, and/or radiation therapy. The treatment of choice for patients with early stage disease is surgical resection in combination with adjuvant chemotherapy. 2 Cisplatin-based adjuvant chemotherapy, in particular, has been shown to improve 5-year survival and disease-free survival among patients with completely resected non–small-cell lung cancer (NSCLC). 3 Even with modern treatment regimens, however, the 5-year survival rate for all stages combined remains 15%. 4 Thus, there is an urgent need for adjuvant therapies that can reduce the risk of relapse and improve survival.
Immunotherapy is an approach under active investigation in both preclinical and clinical settings for a range of human cancers. 5 –7 Recent approval of the first therapeutic anticancer vaccine for prostate cancer (Provenge,® Dendreon, Seattle, WA) by the Food and Drug Administration, together with other preclinical and clinical findings, indicates that vaccine immunotherapy may have potential therapeutic benefit for the treatment of cancer, including lung carcinoma. 7,8 Moreover, recent evidence suggests that specific chemotherapeutic regimens can reduce the tumor growth rate in cancer patients when combined with certain cancer vaccines. 9
The clinical efficacy of standard-of-care chemotherapy regimens relies mostly on direct cytotoxicity to cancer cells. Mounting evidence, however, suggests that certain chemotherapeutic agents have immunomodulatory properties that can be exploited to enhance vaccine-mediated antitumor effects. 5,8,10 In this study, we demonstrate that sublethal multiagent chemotherapy with cisplatin plus vinorelbine enhances the susceptibility of human lung carcinoma to cytotoxic T lymphocyte (CTL)–mediated lysis. These studies also demonstrate that sublethal exposure to cisplatin plus vinorelbine mediates modulation of (a) tumor phenotype, (b) the cytokine/chemokine milieu, and (c) the proapoptotic/antiapoptotic gene ratio. Most importantly, these findings suggest that current treatment strategies for lung cancer can potentially be complemented by active specific immunotherapy approaches. This work thus serves as a rational basis for clinical trials using the standard of care in lung cancer concurrent with immunotherapy.
Materials and Methods
Tumor cell lines
Human lung carcinoma cell lines (A549, NCI-H1703, SKLU-1, NCI-H23, and H460) and colorectal LS174T cells (control) were used in these studies and cultured in media designated by American Type Culture Collection for propagation and maintenance. Cells were incubated at 37°C in a 5% CO2 environment.
Chemotherapeutic drugs
Cisplatin (1 mg/mL; Abraxis Pharmaceuticals, Schaumburg, IL) and vinorelbine (10 mg/mL; Bedford Laboratories, Bedford, OH) were further diluted with sterile phosphate buffered saline (PBS). For in vitro studies, the drugs were further diluted in complete medium, as described below.
Tumor sensitivity to chemotherapy
To evaluate the effects of chemotherapy on the growth of tumor cells, cell lines were exposed in vitro (37°C/5% CO2) for 6 hours to cisplatin and vinorelbine at a concentration of 0.50 μg/mL and 0.04 μg/mL, respectively, unless stated otherwise. Control cells received PBS. Cells were harvested 1, 2, or 3 days after exposure, and the total number of adherent viable cells was determined by trypan blue exclusion.
Phenotypic analysis
To investigate the effects of in vitro exposure of tumor cells to chemotherapy on the cell-surface expression of immune-relevant proteins, adherent tumor cells were harvested 72 hours after treatment with cisplatin and vinorelbine, as described above. Cell surface staining of tumor cells was performed using the primary labeled monoclonal antibodies CD66-FITC, CD95-FITC, CD54-PE, human leukocyte antigen (HLA)-ABC-PE, and the appropriate isotype-matched controls purchased from BD Biosciences (San Diego, CA). Proteins were scored as upregulated if detection levels increased by ≥50% or if mean fluorescence intensity (MFI) increased by ≥30% after chemotherapy. Conversely, proteins were scored as downregulated if detection levels decreased by 10% or MFI decreased by ≥30% compared with untreated samples. Stained cells were acquired on a FACScan or FacsCalibur flow cytometer using CellQuest software (BD Biosciences). Isotype control staining was <5% for all samples analyzed. Dead cells were excluded from the analysis based on side scatter profile.
Carcinoembryonic antigen (CEA)-specific CTL
The CEA-specific and A2-restricted CD8+ CTL line V8T, which recognizes the epitope YLSGANLNL (CAP-1), was maintained and propagated as described previously. 11
Cytotoxicity assays
V8T cells were used on day 4 of the stimulation cycle after Ficoll purification. Tumor cells were left untreated or exposed to chemotherapy for 6 hours. Forty-eight (48) hours after treatment, tumor cells were used as targets in a standard cytotoxicity assay. 10,12 After being labeled using 111Indium oxine (Amersham Health, Silver Spring, MD), tumor cells (2×103) were incubated with V8T cytotoxic T-cells (6×104) (effector-to-target ratio [E:T]) of 30:1 for 18 hours at 37°C with 5% CO2. Targets and CTLs were suspended in complete medium supplemented with 10% human AB serum in 96-well U-bottom plates (Costar, Cambridge, MA). After incubation, supernatants were collected and assayed on a Cobra Auto-Gamma counter (Packard Instruments, Downers Grove, IL). The percentage of specific lysis was determined by the standard equation: % specific lysis=[(experimental−spontaneous)/(maximum−spontaneous)]×100. For major histocompatibility complex (MHC)-I blocking studies, tumor cells were incubated with anti-HLA-A2 mAb (20 μg/mL, AbD Serotec, Raleigh, NC) or isotype control mAb (IgG2b, 20 μg/mL, AbD Serotec) for 1 hour at 37°C before being used as CTL targets as above. In certain experiments, V8T CTLs were preincubated for 2 hours with 100 nmol/L concanamycin A (CMA), a specific inhibitor of perforin-dependent lysis. CTLs were then used as effectors against tumor targets in a standard 18-hour cytotoxicity assay in the presence of CMA.
Cytokines
To examine the effect of cisplatin plus vinorelbine on the production of cytokines, A549 cells were left untreated or exposed in vitro for 6 hours to cisplatin (0.50 μg/mL) and vinorelbine (0.04 μg/mL). At 3 days after exposure, supernatants were harvested and analyzed for protein levels of interleukin (IL)-1β, IL-2, IL-4, IL-5, IL-10, IL-12p70, interferon (IFN)-γ, and tumor necrosis factor (TNF)-α, by Cytokine Bead Array (BD Biosciences), and IL-8 and transforming growth factor (TGF)-β by ELISA (R&D Systems, Minneapolis, MN), according to the manufacturers' instructions.
Molecular array studies
To investigate the effects of in vitro exposure of tumor cells to chemotherapy on the modulation of genes involved in CTL sensitivity, tumor cells were harvested 48 hours after 3 hours of exposure to cisplatin and vinorelbine. Two arrays were used: a human genome array (U133 GeneChip, Affymetrix, Santa Clara, CA), which monitors >38,500 genes (assays performed by Expression Analysis, Durham, NC), and an apoptosis OligoArray (SuperArray, Bioscience Corp, Frederick, MD), which focuses on 384 genes involved in apoptosis. These arrays were used to analyze gene expression changes pre- and post-treatment with cisplatin/vinorelbine. Post-treatment, genes were considered to be upregulated or downregulated if their normalized intensity ratio was ≥2 or ≤0.5 (twofold cutoff) as previously described. 12
Statistical analysis
Statistical analyses were performed using GraphPad Prism 4.0a for Macintosh (GraphPad Software, San Diego, CA). Differences in cytotoxicity between treatments were evaluated using an unpaired Student t test with a two-tailed distribution at a 30:1 E:T cell ratio. Results of all tests for statistical significance are given as p values calculated using a confidence interval of 95%. In graphic representations of data, y-axis error bars indicate the standard error. In some cases, the variation is such that the plot symbol obscures the error bars. Significant differences in the distribution of flow cytometry analysis data were confirmed by the Kolmogorov-Smirnov test, using CellQuest software (BD Biosciences).
Results
Combination therapy with multiagent chemotherapy (cisplatin and vinorelbine) is sublethal
Adjuvant chemotherapy with cisplatin-based regimens is standard of care for resectable NSCLC. To determine the phenotypic and functional consequences of exposure of carcinoma tumor cell lines to sublethal cisplatin/vinorelbine chemotherapy, five biologically distinct NSCLC (NCI-H1703, A549, NCI-H23, SKLU-1, and H460) tumor cell lines and one colon carcinoma (LS174T) cell line were chosen (Fig. 1). To identify sublethal doses of chemotherapy, dose-response studies were performed using various concentrations of cisplatin and vinorelbine based on reported clinical unbound peak plasma concentrations and relative ratios between peak plasma concentrations of both drugs. 13 Tumor cells were exposed in vitro to cisplatin (0.5 μg/mL) and vinorelbine (0.04 μg/mL) for 6 hours, or were left untreated.
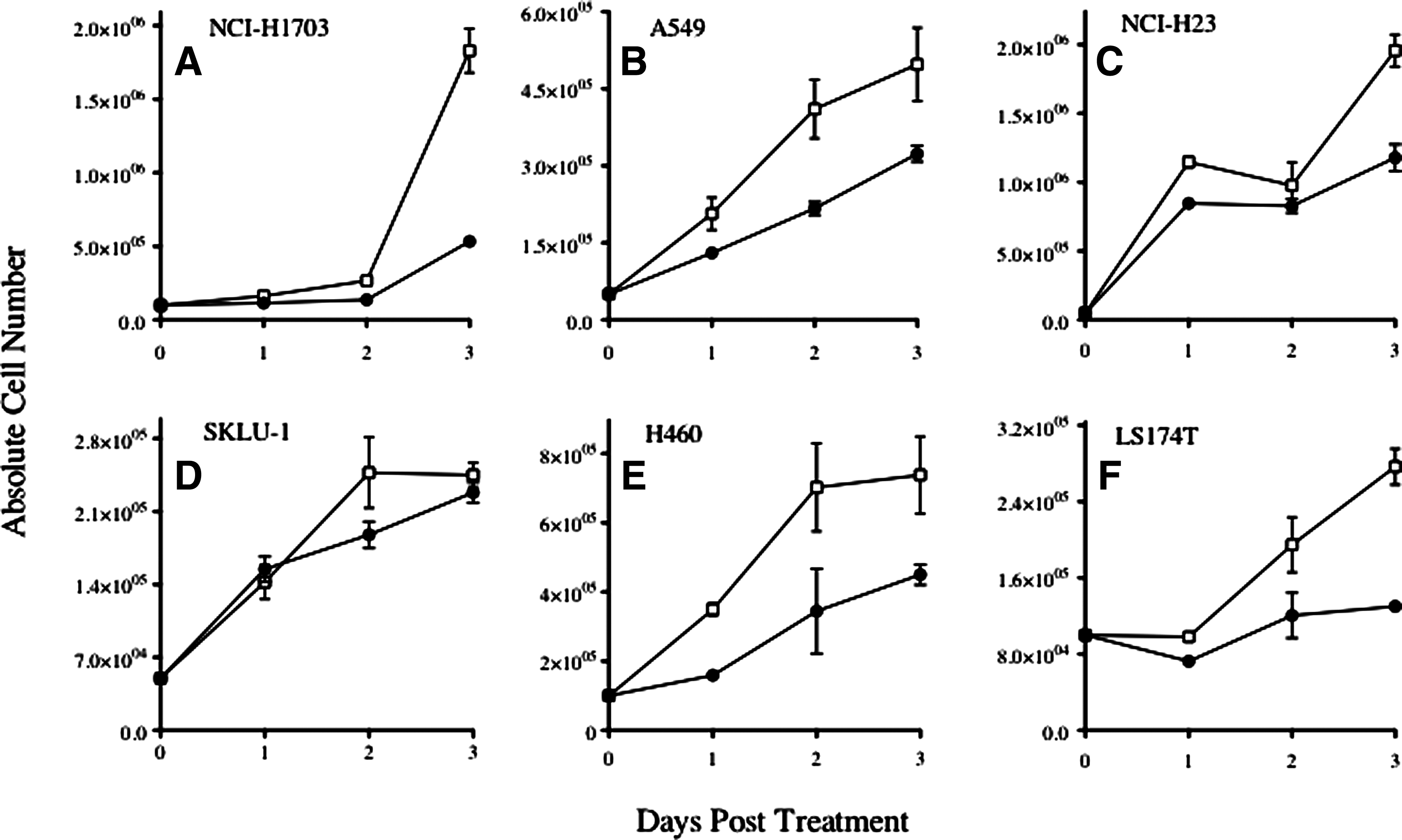
Combination therapy with multiagent chemotherapy (cisplatin and vinorelbine) is sublethal. Lung (NCI-H1703, A549, NCI-H23, SKLU-1, H460) and colorectal (LS174T) carcinoma cells were exposed in vitro to cisplatin (0.5 μg/mL) and vinorelbine (0.04 μg/mL) for 6 hours (closed circles) or left untreated (open squares). At various times after treatment, the total number of viable cells was determined by trypan blue exclusion. Results presented as mean±standard error of the mean from three replicate flasks.
At 1, 2, and 3 days post-treatment, the total number of adherent viable cells was determined by trypan blue exclusion; cells were harvested before confluency. As shown in Figure 1, exposure of all cell lines to cisplatin/vinorelbine had minimal (SKLU1) to moderate (NCI-H1703, A549, NCI-H23, H460, and LS174T) effect on the cellular growth of treated cells relative to untreated controls during the 3 days after chemotherapy. Importantly, all five lung cell lines were proliferating at day 3. No significant decreases of viability were observed, as determined by trypan blue staining for all cell lines (not shown).
Multiple changes in surface protein expression in human lung cancer cell lines after treatment with cisplatin/vinorelbine chemotherapy
Studies with various human cell lines have demonstrated cell surface modulation of immunologically relevant molecules in response to chemotherapy, 14 including with cisplatin-based therapies. 10,15,16 Furthermore, because several cell-surface proteins on tumor target cells have previously been shown to be critical for interactions with CD8+ T-cells, we next examined the effect of cisplatin/vinorelbine exposure on the phenotype of human carcinoma cell lines. Each of the five surface molecules monitored in this study (intercellular adhesion molecule 1 [ICAM-1], Fas, MHC class I, MUC-1, and the tumor-associated antigen [TAA] CEA) has been implicated in enhancing antitumor T-cell responses through diverse mechanisms. 12,17 –20 We sought to quantify the expression of these proteins in each of the five lung carcinoma cell lines pre- and post-treatment.
Human tumor cells were treated in vitro for 6 hours with cisplatin (0.5 μg/mL) and vinorelbine (0.04 μg/mL), or were left untreated. Cell-surface expression of CEA, ICAM-1, Fas, and MHC class I molecules was monitored by flow cytometry 2 days after treatment (Table 1). Whereas the cell lines exhibited different levels of sensitivity to chemotherapy, treatment modulated the expression of the surface proteins analyzed. As an example, the treatment of A549 cells with cisplatin/vinorelbine increased the population of CEA+ cells from 5% to 18%, a 3.6-fold change (Table 1). In addition, MFI of CEA+ cells increased 62% after treatment. Overall, a significant increase in the population of CEA+ cells and/or MFI was observed after treatment in all (five of five) lung cancer cell lines. An augmented effect on Fas expression was observed in H460 cells. In addition, the population of A549 cells expressing ICAM-1 increased 61%, with a concomitant increase in MFI of nearly 170%. Increased ICAM-1 expression was also observed after exposure to cisplatin/vinorelbine in H460. MHC I expression was also augmented in A549 cells, where MFI after treatment was >eight-fold over that of control cells. In total, each of the five tumor cell lines responded to low-dose chemotherapy by upregulating one or more immunologically relevant surface molecules. LS174T is a CEA+ human colon carcinoma cell line with very low (2.1%) HLA-A2 expression 21 and was used as a control. These cells significantly upregulated CEA, Fas, ICAM-1, and MHC-1 after exposure to chemotherapy (Table 1).
Human tumor cells were treated in vitro for 6 hours with 0.5 μg/mL cisplatin/0.04 μg/mL vinorelbine, or left untreated, and then analyzed by flow cytometry after 48 hours. Cells were analyzed for CEA, Fas, ICAM-1, and MHC class I surface expression after each treatment. Numbers indicate the percentage of positive cells. Numbers in parentheses denote MFI. Isotype control antibody staining was <5% in all samples. Bold=marked upregulation (≥50% increase in percent of cells or 30% increase in MFI not observed in isotype control).
Tumor cells treated with chemotherapy demonstrate significantly increased sensitivity to antigen-specific cytotoxic T-cell killing
We have previously shown that treatment of murine lung carcinoma cells with cisplatin/vinorelbine increased tumor sensitivity to CTL-mediated lysis. 8 Further, we have also demonstrated that cisplatin/5-fluorouracil 10 (5-FU) chemotherapy enhanced antigen-specific CTL killing of human tumor cells. To evaluate the functional significance of phenotypic changes in human lung tumor cells after exposure to sublethal doses of cisplatin plus vinorelbine, we examined the sensitivity of five lung carcinoma cell lines to CTL-mediated killing post-treatment.
To address this issue, a human HLA-A2–restricted CEA-specific CD8+ T-cell line that recognizes an HLA-A2–restricted epitope on human CEA-expressing carcinoma cells was selected. 11 It has been previously shown that CEA is overexpressed in several malignancies, including lung cancer. Carcinoma lines positive for HLA-A2 and CEA (A549, NCI-H1703, NCI-H23, H460, and SKLU-1) were subjected to no treatment or chemotherapy with cisplatin/vinorelbine for 6 hours. The CEA+, HLA-A2- carcinoma cell line LS174T was used as a control. After 2 days, the sensitivity of carcinoma cells to CEA-specific killing was determined in a standard 18-hour assay (Fig. 2). All five HLA-A2+, CEA+ tumor cell lines showed substantially enhanced CTL killing after exposure to chemotherapy when compared with their respective untreated controls (Fig. 2A–E). Untreated NCI-H1703 cells were minimally killed; exposure to chemotherapy, however, resulted in a significant enhancement of tumor cell lysis (Fig. 2A; p=0.001, vs no treatment). A549 cells were poorly killed at baseline; treatment with cisplatin/vinorelbine, however, resulted in a significant increase of tumor cell lysis (Fig. 2B; p<0.001, vs no treatment). Similar results were observed with NCI-H23 and H460 tumor cells (Fig. 2C and E; p=0.002 and p=0.04, vs no treatment, respectively). Untreated SKLU-1 also showed minimal lysis at all E:T ratios; however, chemotherapy offered significant enhancement of CTL-mediated killing (Fig. 2D; p=0.003, vs no treatment). SKLU-1 showed the highest level of killing by CTL when exposed to cisplatin/vinorelbine. Thus, in five of five CEA+, HLA-A2+ cell lines tested, the highest level of lysis was seen after cells were exposed to chemotherapy.

Tumor cells treated with chemotherapy demonstrate significantly increased sensitivity to antigen-specific cytotoxic T-cell killing. Human tumor cells received no treatment (open squares), or the combination chemotherapy (closed circles) for 6 hours. Two days after treatment, cells were labeled with 111In, and used as targets in a CTL lysis assay. After CEA-specific CD8+ T-cells were incubated at an E:T ratio of 30:1, 15:1, or 10:1 for 18 hours, supernatants were harvested and specific lysis was determined. Results are presented as mean±standrd error of the mean from three replicate wells.
To confirm that the enhanced sensitivity of lung cancer tumor cells after chemotherapy to CTL-mediated lysis was restricted to MHC class I+ tumors, untreated and treated LS174T cells were used as targets for CEA-specific killing. LS174T is a CEA+ human colon carcinoma cell line with very low (2.1%) HLA-A2 expression. 21 Untreated LS174T cells were not killed by the HLA-A2–restricted, CEA-specific CTL cells used in these experiments (Fig. 2F). Furthermore, chemotherapy did not increase its tumor sensitivity to CTL-mediated lysis (p=0.3448), despite its modulation of phenotype (Table 1).
Exposure of lung tumor cells to cisplatin plus vinorelbine renders them more sensitive to perforin/granzyme–mediated CTL killing
T-cell cytotoxicity on target tumor cells is mediated through two main mechanisms—ie, perforin/granzyme and Fas/FasL death pathways. 22 Thus, to examine the contribution of these distinct mechanisms on tumor cell lysis after chemotherapy, we focused on the sensitivity of A549 and SKLU-1 lung carcinoma cells to killing by CEA-specific CTLs in the presence of CMA in a standard 18-hour assay. CMA inhibits perforin-mediated cytotoxicity without affecting Fas-mediated killing. 23 Tumor cells were exposed to cisplatin plus vinorelbine, or were left untreated. CEA-specific killing of A549 and SKLU-1 carcinoma cells after chemotherapy was significantly increased (p<0.05) relative to that of untreated cells (Fig. 3A). In the presence of CMA, however, lysis of both A549 and SKLU-1 cells exposed to chemotherapy was similar to that of untreated tumor targets in the absence of CMA, suggesting that the increased sensitivity of tumor cells to CTL cytotoxicity as a result of chemotherapy is mediated by perforin/granzyme cell-death pathway.
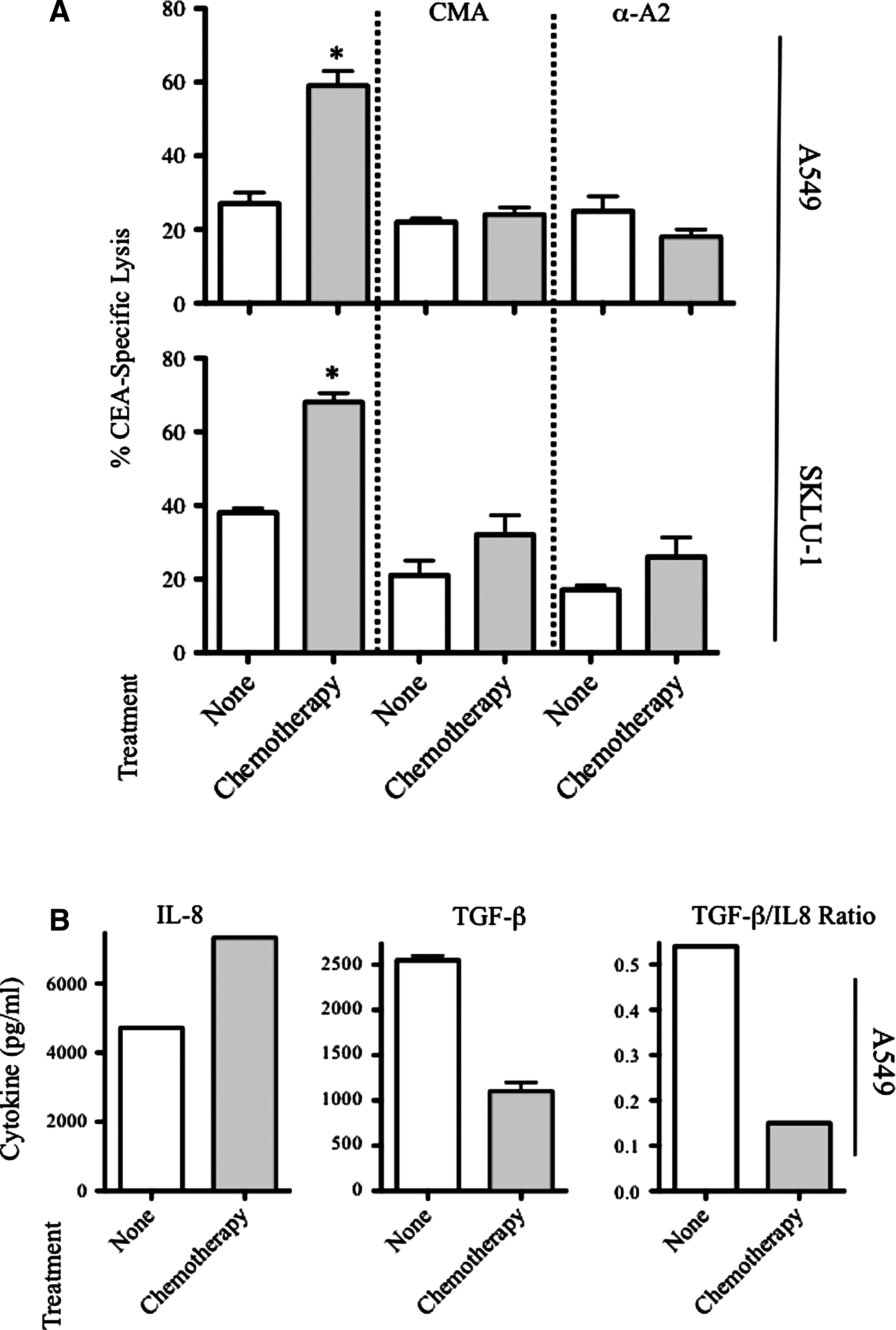
Increased CTL sensitivity mediated by chemotherapy is dependent on MHC and perforin/granzyme.
To further examine the mechanism of T-cell killing after exposure to the platinum doublet, A549 and SKLU-1 cells were treated with cisplatin/vinorelbine before being used as targets in a cytotoxic T-cell assay in the presence of an anti-HLA-A2 blocking antibody or matched isotype control. As shown in Figure 3A, CTL killing of treated A549 or SKLU-1 carcinoma cells was reduced to pretreatment levels in the presence of MHC blocking, confirming MHC-restricted CTL killing. Taken together, these data indicate that exposure of human lung carcinoma cells to cisplatin/vinorelbine increases their sensitivity to MHC-restricted perforin/granzyme CTL-mediated lysis.
Sublethal doses of chemotherapy modulate the production of cytokines in a net CTL-activating fashion
To better understand the mechanisms involved in the enhancement of carcinoma sensitivity to CTL killing after exposure to sublethal chemotherapy, we examined the cytokine secretion profile of A549 cells in response to cisplatin/vinorelbine exposure, including that of IL-8 and TGF-β. IL-8 is a proinflammatory mediator that can be secreted in response to chemotherapy and has been associated with the development of resistance to apoptosis, including when mediated by cisplatin. 24,25 TGF-β has been shown to suppress CTL-mediated killing of malignant cells. 26 Thus, to address the role of cytokines in CTL-mediated lysis of lung carcinoma cells, A549 tumor cells were left untreated or exposed to cisplatin plus vinorelbine, and supernatants were analyzed for protein contents of IL-1β, IL-2, IL-4, IL-5, IL-10, IL-12p70, IFN-γ, TNF-α, IL-8, and TGF-β.
No difference in the secretion of IL-1β, IL-2, IL-4, IL-5, IL-10, IL-12p70, IFN-γ, or TNF-α was observed on exposure to chemotherapy (not shown). A549 cells, however, responded to chemotherapy by increasing IL-8 secretion 1.5-fold. In contrast, tumor cells demonstrated a statistically significant decrease in TGF-β secretion (p=0.006) on exposure to chemotherapy relative to untreated controls (Fig. 3B), suggesting that low exposure to cisplatin plus vinorelbine renders tumor cells more sensitive to CTL lysis given their diminished capacity of TGF-β secretion. Analysis of the ratio between TGF-β and IL-8 concentrations indicated that exposure to the platinum doublet markedly reduced TGF-β/IL-8 secretion per cell. Taken together, these results suggest that exposure to sublethal doses of cisplatin plus vinorelbine modulates tumor phenotype and alters the production of cytokines in a net CTL-activating fashion.
Differentially expressed genes in multiagent chemotherapy-treated lung cancer cells
To investigate the effects of in vitro exposure of tumor cells to chemotherapy on the modulation of genes involved in CTL sensitivity, A549 and SKLU-1 tumor cells were exposed to cisplatin plus vinorelbine, or were left untreated. Forty-eight (48) hours after treatment, differences in gene expression were examined, as described in Materials and Methods. Using a Human Genome U133 gene array, quantitative analysis of the difference in gene expression of more than 38,500 genes, between treated and untreated A549 cells, resulted in a total of 935 genes upregulated after chemotherapy by at least twofold. In contrast, 810 genes had at least twofold decreased mRNA levels in response to treatment (Fig. 4A). Exposure of SKLU-1 tumor cells to the platinum doublet resulted in 105 genes with increased expression; 35 other genes were repressed at least twofold in response to treatment.
Differentially expressed genes in multiagent chemotherapy-treated lung cancer cells. A549 and SKLU-1 cells were treated with cisplatin/vinorelbine for 3 hours. After 48 hours, a human genome microarray analysis was performed. Genes were considered differentially expressed if their levels of expression differed at least twofold compared with matched untreated control cells.

Further analysis focused on those genes that exhibited a shared change in expression of at least twofold in both A549 and SKLU-1 tumor cells, which corresponded to seven genes being upregulated and one gene showing decreased expression at 48 hours postchemotherapy. These genes were then grouped in accordance with their presumed function (Fig. 4B). Also shown in Figure 4B is the normalized relative intensity ratio for each individual gene. IL-8, one of the seven genes differentially upregulated after treatment, is a chemoresponse gene that encodes the proinflammatory and angiogenic chemokine IL-8. After exposure to cisplatin/vinorelbine, IL-8 expression was enhanced 3.8- and 6.4-fold in A549 and SKLU-1 cells, respectively. The upregulation of IL-8 gene expression was in accordance with the observations at the protein level previously observed (Fig. 3B).
The gene encoding chemokine ligand CXCL5 was also upregulated in response to chemotherapy. CXCL5 is upregulated in response to inflammation and stimulates the chemotaxis of neutrophils. 27,28 Other inflammation-response genes associated with tumor cell survival were upregulated after chemotherapy. These included PLCXD2, a phospholipase involved in signal transduction that has the ability to release CEA from the surface of tumor cells, and GLS, which encodes for the antioxidant enzyme glutaminase, an active participant in tumor response to cisplatin-induced oxidative stress. 29 It has recently been shown that vinorelbine binds histones with high affinity, which precedes chromatin compaction. 30,31 As would be expected, a set of four genes that participate in the chromatin repair process was also differentially regulated in both cell lines in response to chemotherapy, such as HIST1H4C, which encodes a histone cluster 1 protein involved in nucleosome assembly. 29,32 More importantly, TFPI2, the gene encoding for the tumor suppressor tissue factor pathway inhibitor 2 was upregulated significantly after exposure to cisplatin/vinorelbine. TFPI2 has been identified as a Kunitz-type serine protease inhibitor and has been shown to inhibit invasion, tumor growth, and metastasis. 33
Differentially expressed apoptosis genes in multiagent chemotherapy-treated lung cancer cells
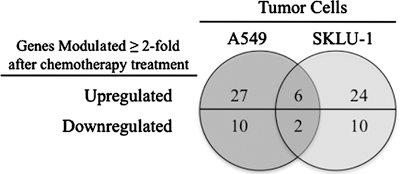
Next, we focused on the molecular basis for the enhanced lytic response after exposure of lung tumor cells to the platinum doublet by examining the expression of proapoptotic and antiapoptotic genes. A549 and SKLU-1 tumor cells were left untreated or were exposed to cisplatin plus vinorelbine as described. At 48 hours postexposure, differences in gene expression pre- and post-treatment were evaluated using an OligoArray, which focuses on 384 genes involved in apoptosis. Post-treatment, genes were considered to be upregulated or downregulated if their normalized intensity ratio was ≥2 or ≤0.5 (twofold cutoff).
Quantitative analysis of the difference in gene expression post-treatment relative to untreated A549 cells resulted in a total of 27 apoptosis-related genes upregulated after chemotherapy (Fig. 5A). In contrast, 10 genes had at least twofold decreased mRNA levels in response to treatment. Exposure of SKLU-1 tumor cells to cisplatin plus vinorelbine resulted in 24 genes with increased expression, while 10 genes were downregulated. Further analysis identified a total of eight genes that shared a significant change in expression in both A549 and SKLU-1 tumor cells, which corresponded to six genes being upregulated at 48 hours postchemotherapy (Fig. 5B). These included BBC3, which encodes PUMA, a p53 transcriptional target and proapoptotic gene involved in cisplatin-induced apoptosis as well as endoplasmic reticulum stress induced by vinka alkaloids, including vinorelbine. 34 Two members of the tumor-necrosis superfamily of genes were also upregulated after treatment in both cell lines—ie, TNF and TNFSF15. TNF, a strong mediator of inflammatory responses, can induce apoptosis via death domain receptors. 35 Although TNF gene expression was strongly upregulated after treatment, it did not translate into increased protein levels (not shown). Both TNFSF15 and TNFSF14 act as autocrine factors to induce cell death. 35 However, whereas TNFSF15 was upregulated 2.23 to 2.56-fold in response to chemotherapy, the expression of TNFSF14 was 3.43 to 4.93-fold less after exposure to the platinum doublet.
Differentially expressed apoptosis genes in multiagent chemotherapy-treated lung cancer cells. A549 and SKLU-1 cells were treated with cisplatin/vinorelbine for 3 hours. After 48 hours, a focused apoptosis OligoArray analysis was performed. Genes were considered differentially expressed if their levels of expression differed at least twofold compared with matched untreated control cells.
Of the four antiapoptotic genes that were differentially regulated after exposure to cisplatin plus vinorelbine, only NOX5 was mildly downregulated. 36 In contrast, the pro-survival genes BCL2L10, IL1A, and TNFRSF10D were significantly upregulated in response to chemotherapy. Overexpression of the tumor suppressor BCL2L10 has been shown to suppress apoptosis in gastric cancer cells. 37 IL1A encodes IL-1α, a proinflammatory cytokine known to stimulate tumor growth and IL-8 expression through NF-kB activation. 38,39 TNFRSF10D encodes the TRAIL decoy receptor 2 (DcR2), also designated as TRAIL-R4.
Discussion
The clinical efficacy of standard-of-care chemotherapy regimens relies mostly on direct cytotoxicity to cancer cells and is often mediated by maximum tolerated doses of antineoplastic agents. Multiple investigations suggest that patients undergoing primary cancer treatment have tumor cell populations that are ultimately exposed to sublethal doses of chemotherapy. Noncytotoxic exposures have been associated with the upregulation of several classes of molecules on tumor cells, including HLA molecules, TAAs, costimulatory molecules, adhesion molecules, components of antigen processing pathways, inflammatory mediators, cytokines, and death receptors. 10,40,41 These alterations have been shown in vitro to enhance immune-mediated killing in several human carcinoma cell lines. 12 Certain agents can also induce immunogenic death of tumor cells, leading to activation of dendritic cells, followed by antigen presentation and cross-presentation to T-cells, resulting in CTLs with greater and more efficient cytotoxic potential. 42,43 Recent evidence also suggests that specific chemotherapeutic regimens can reduce the tumor growth rate in cancer patients when combined with certain cancer vaccines. 9
Platinum alkylating agents such as cisplatin are commonly used to treat a variety of malignancies, including lung cancer. Accumulating evidence suggests that nonlethal concentrations of these agents can induce immune-relevant changes in tumor cells that can be exploited in a combined chemotherapy/vaccine regimen to achieve potent antitumor immunity. 5,8 In Europe and the United States, cisplatin combined with vinorelbine is a standard-of-care treatment for NSCLC that has shown promise in the adjuvant setting. 3 Cisplatin has been shown to modulate both murine and human tumor cell characteristics toward a more immunogenic phenotype. 16,44 To the best of our knowledge, the effects of cisplatin/vinorelbine on human tumor sensitivity to T-cell cytotoxicity and its molecular mechanisms have not been reported. In this study we examined the effect of cisplatin/vinorelbine on growth, cell-surface phenotype, and CTL-mediated lysis of five distinct human lung carcinoma cell lines in vitro and explored its molecular mechanisms.
Our results demonstrate that sublethal doses of cisplatin plus vinorelbine (Fig. 1) enhance MHC-restricted CTL-mediated killing (Fig. 2), modulate tumor phenotype (Table 1) and alter gene expression profile (Figs. 4 and 5) and cytokine secretion (Fig. 3) of human lung carcinoma cells. Each of the five surface molecules monitored in this study (Fas, CEA, MUC-1, ICAM-1 and MHC class I) has been implicated in enhancing antitumor T-cell responses through diverse mechanisms. Binding of Fas to Fas receptor on cytotoxic T-cells is one of the main mechanisms through which CTLs directly induce apoptosis of tumor targets. 17 ICAM-1 has both cell adhesion and costimulatory properties. It has been demonstrated that increased expression of adhesion molecules on tumor cells correlates with increased T-lymphocyte binding and killing of tumor targets. 45 In addition, increased ICAM-1 expression could enhance immune destruction via direct costimulation of T-cells, thus making tumor cells better immunogens. CEA is a tumor-associated antigen differentially expressed in tumors versus normal tissues, 18 and MHC class I molecules are important for antigen presentation of tumor molecule epitopes to circulating T-cells. 20,46
Several reports have demonstrated that the downregulation of these genes is a common mechanism used by tumors to escape immune recognition and elimination. 47 It therefore seems likely that the upregulation of these genes by cisplatin/vinorelbine could make tumor cells more susceptible or amenable to attack by the immune system. Further, increased sensitivity to antigen-specific CTLs was also observed in human colon carcinoma cell lines treated with cisplatin alone, an effect associated with enhanced expression of ICAM-1 and Fas. 48 Similar results have been reported with other cisplatin doublets. In one study, exposure of HNSCC cell lines to cisplatin plus 5-FU resulted in a synergistic increase of ICAM-1. 49
It is notable that all five lung cancer cell lines varied as to their origin, previous therapies, epithelial/mesenchymal balance, p53, and oncogenic Ras status. 50,51 Moreover, each cell line showed a distinct phenotypic signature in response to chemotherapy. They were all lysed to a significantly greater degree after exposure to cisplatin/vinorelbine, however. For all tumor cell lines tested, the only discernible pattern of upregulation of any surface molecule examined that correlated with enhanced sensitivity to CTL (Fig. 2) was to some extent in CEA and, to a much lesser degree, MHC I and ICAM-1 expression (Table 1). While it is possible that the array of molecules examined could contribute to enhanced CTL killing of tumor targets, it is also likely that many other proteins could contribute to these observations, including cytokines/chemokines.
Lung tumor cells responded to treatment with the platinum doublet by markedly decreasing TGF-β secretion as well as the ratio per cell of TGF-β/IL-8 proteins, despite IL-8 gene upregulation (Fig. 4) and increased IL-8 protein secretion (Fig. 3). IL-8 is a proinflammatory mediator and an angiogenic growth factor overexpressed in various malignancies, including carcinomas of the lung. 52 IL-8 can be secreted in response to chemotherapy and has been associated with the development of resistance to apoptosis, including when mediated by cisplatin. 24,25
TGF-β is known to be the most potent inducer of epithelial-to-mesenchymal transition in carcinoma cells. 53 The presence of TGF-β in the tumor microenvironment can have a pronounced effect on the antitumor activity of T-cells; for example, TGF-β has been shown to inhibit the differentiation of effector T-cells and suppress CTL-mediated lysis of carcinoma cells. 26 In addition, TGF-β promotes the differentiation of regulatory T-cells that are known to be an abundant source of TGF-β production. TGF-β secretion modulates the tumor microenvironment by promoting migration and invasion while adjacent immune effectors are prevented from destroying the invasive carcinoma cells in response to TGF-β stimulation. Results presented here (Fig. 3B) indicate that sublethal exposure of lung tumor cells to cisplatin/vinorelbine modulates tumor toward a lower metastatic phenotype and alters the production of cytokines in a net CTL-activating fashion.
It has been established that the ratio of proapoptotic and antiapoptotic proteins play a critical role in deciding the ultimate fate of a cell. 54 Studies have confirmed that central molecular mediators of apoptosis can impair the tumor suppressive activity of antigen-specific CTL. 55 Taken independently, this would seemingly limit the ability of immunotherapy to destroy malignant populations preselected to express high levels of antiapoptotic proteins. Thus, observations in this study were extended to include the analysis of gene expression in response to sublethal chemotherapy, using a human genome array and a focused apoptosis array.
We examined the gene expression profile of two lung tumor cell lines to identify a shared gene signature response to sublethal cisplatin/vinorelbine. Using this broad-based approach, we were able to identify a total of eight molecules modulated by chemotherapy in both A549 and SKLU-1 cells that can be exploited to increase the sensitivity of a tumor target to an immunotherapy-mediated attack (Fig. 4). For instance, the upregulated molecules GLS and PLCXD2 could serve as potential tumor-associated molecules and targets for tumor-specific CTL. IL-8 expression was increased in response to treatment at both gene and protein levels (Fig. 3B). Although A549 and SKLU-1 cells showed different sensitivity to sublethal exposure to cisplatin/vinorelbine (Fig. 1), both cell lines were significantly more sensitive to CTL-mediated lysis, suggesting that IL-8 may modulate cisplatin-induced perforin/granzyme CTL killing of tumor targets. CXCL5, a gene encoding a second proangiogenic chemokine, was also upregulated. CXCL5 stimulates chemotaxis of neutrophils and can be upregulated in response to inflammatory cytokines such as the antiapoptotic IL-1α or TNF, 27,28 both of which were also shown to be upregulated after chemotherapy (Fig. 5).
It has been shown recently that vinorelbine binds histones with high affinity, which precedes chromatin compaction. 30,31 As would be expected, a set of four genes that participate in the chromatin repair process was also differentially regulated in both cell lines in response to chemotherapy, suggesting an active cell repair/survival process. 29,32 This is in agreement with the upregulation of antiapoptotic genes observed after treatment (Fig. 5).
Certain proapoptotic genes were also upregulated in response to cisplatin plus vinorelbine and may be part of the repair response to low-dose nonlethal chemotherapy. These included BBC3, which encodes PUMA, a p53 transcriptional target also involved in the response to endoplasmic reticulum stress induced by vinka alkaloids, including vinorelbine. 34,56,57 Sublethal exposure of both cell lines to cisplatin plus vinorelbine increased the expression of TFPI2, a gene encoding the protein tissue factor pathway inhibitor 2 (Fig. 4). This blood coagulation and serine protease inhibitor was recently identified as a potent tumor suppressor and demonstrated to inhibit invasion, tumor growth, and metastasis. 33,58 –60 In a recent study, PUMA upregulation has also been associated with increased sensitivity to cisplatin as a result of impaired expression of slug, a transcription factor associated with promotion of tumor invasion and metastasis. 61 These results further support the decreased secretion of TGF-β observed after treatment, suggesting a modulation of the tumor toward a less aggressive biology. It remains to be discovered, however, what the implications of these results will be on tumor invasion and metastasis, as well as the potential relationship between of IL-8 and TGF-β with TFPI2 in the context of sublethal exposure to cisplatin plus vinorelbine.
It has been demonstrated that certain chemotherapeutic regimens trigger cancer cell death while stimulating immune responses against the tumor. This immunogenic cell death and subsequent immune responses have been shown to be critical for the clearing of tumor cells that survive therapy. 62,63 There is another possible mechanism, however, by which chemotherapy can be used in combination with active vaccination. The data presented here propose a complex mechanism that is distinct from and complementary to that of immunogenic cell death. One of the hallmarks of immunogenic cell death has been suggested to be calreticulin translocation. In addition to the modulation of tumor phenotype, cytokine/chemokine, and apoptotic gene ratio in tumors treated with cisplatin/vinorelbine, our laboratory has preliminary data of membrane translocation of calreticulin after chemotherapy or radiation, suggesting that chemotherapy and radiation therapy induced a complex continuum of events resulting in increased T-cell sensitivity and induction/amplification of tumor-specific immune responses.
Taken together, these results form the rational basis for clinical trials using the current platinum-based standard of care in lung cancer in combination with T-cell–mediated immunotherapy. We envision a translational path to clinically test these findings in the adjuvant setting by immunizing patients with early-stage NSCLC after surgical resection. Patients would be vaccinated with tumor vaccines to generate tumor-specific T-cells, and then undergo standard-of-care chemotherapy and receive vaccine boosts between chemotherapy cycles. These vaccines, which consist of recombinant poxviruses expressing the transgenes CEA and MUC-1 and three T-cell costimulatory molecules (TRICOM), 64 are currently in Phase II and Phase III clinical trials. It is hoped that such adjuvant therapy will reduce the risk of relapse and improve survival in NSCLC patients.
Footnotes
Acknowledgments
We thank Dr. Jeffrey Schlom, NCI, NIH, for his helpful suggestions in the review of this manuscript. The authors thank Marion Taylor for excellent technical assistance, and Debra Weingarten for her editorial assistance in the preparation of this manuscript. This research was supported by the Intramural Research Program of the Center for Cancer Research, National Cancer Institute, National Institutes of Health.
Disclosure Statement
No competing financial interests exist.