
Abstract
Transitional cell carcinoma of bladder (TCCB) is a common malignancy worldwide, and outcomes for patients with advanced bladder cancer remain poor. To study the pathogenesis of TCCB, we investigated roles of Phospholipase C (PLC)ɛ, an effector of Ras and Rap small GTPases. RNA interference was used to knockdown PLCɛ expression in human bladder cancer cell lines (BIU-87 and T24). The expression levels of PLCɛ mRNA and protein were detected by reverse transcriptase–polymerase chain reaction and Western blot, respectively. Flow cytometry (FCM) was used to detect distribution of cell cycle. Cellular apoptosis was reflected by transmission electron microscopy and the expression of bcl-2 and bax. We found that PLCɛ could be efficiently knocked down by shRNA. FCM assay showed that the pGenesil-PLCɛ-transfected cells were arrested at the G0/G1 phase. Silence of PLCɛ might induce apoptosis via modulation of bcl-2 and bax. In conclusion, our results suggest that PLCɛ plays an important role in the pathogenesis of human bladder cancer cells. PLCɛ may be used as a potential target of gene therapy for bladder cancer in future.
Introduction
Bladder cancer is the most common cancer in the urinary system, and it is estimated that there were about 357,000 new cases and 145,000 deaths worldwide 1 year. 1 Despite much research on this topic, it is still difficult to predict tumor progression, optimal therapy, and clinical outcome. Therefore, ongoing effects must be performed to clarify the molecular mechanism of tumor development and carcinogenesis in bladder cancer.
The most frequently detected alterations in oncogenes in animal and tumor models of human cancer are mutations in the RAS family of oncogenes. Several studies have provided evidence that Ras mutation is a major risk for bladder cancer. 2,3 The RAS gene family codes for 21-kDa (p21RAS) proteins that are found in the cytoplasm, and it is associated with the inner surface of the plasma membrane; the normal function of p21RAS proteins is to interact with tyrosine kinase receptors to activate a signal transduction pathway. 4 Most of the mutations are point mutations in codons 12, 13, or 61, which convert the ras gene to a transforming oncogene. 5 The mutated p21RAS has a structure that obstructs its ability to bind to a GTPase-activating protein, thus keeping p21 in a guanosine triphosphate-bound activated state.
Phospholipase C (PLC)ɛ is one of the members of the phosphoinositide-specific PLC family that catalyzes the hydrolysis of a membrane phospholipid, phosphatidylinositol 4,5-bisphosphate, to generate two important second messengers, diacylglycerol and inositol 1,4,5-trisphosphate. Diacylglycerol and inositol 1,4,5-trisphosphate induce the activation of protein kinase C (PKC) and mobilization of calcium from intracellular stores, respectively. 6,7 PLCɛ is characterized by the presence of a Ras-associating domain at its C-terminus and a CDC25-like domain at its N-terminus. Studies focusing on PLCɛ interaction with Ras family small GTPases demonstrated that PLCɛ served as a direct downstream effector of Ras small GTPases, Rap1 and Rap2. 8 –10 Given the importance of Ras- and Rho-family GTPases in the generation and progression of tumors, several studies have aimed to establish a possible role for PLCɛ in the context of tumor cells and cancer development. 11 –14 In our previous study, we found that PLCɛ mRNA expression in transitional cell carcinoma of bladder (TCCB) tissues was significantly higher than that in normal bladder tissues, and semiquantitative polymerase chain reaction (PCR) confirmed that the PLCɛ mRNA level was significantly corrective with the stages of TCCB, but not the grades. 15 PLCɛ also appears to play a crucial role in bladder-cancer cancer cell invasion. 16 However, the questions of whether PLCɛ is involved in regulating bladder cancer apoptosis still remain to be answered.
Apoptosis is a genetically regulated process of cell suicide that is modulated by a variety of cellular signaling pathways. 17 Cell homeostasis is maintained through a balance between cell proliferation and cell death. Cancer maybe occurred when the balance is broken. Caspases and Bcl-2 are critical in regulation of apoptosis. Bcl-2 is an apoptosis-inhibiting gene, plays a central role in the execution of apoptosis, and was first identified as an oncogene from follicular B-cell lymphoma. 18 Members of the Bcl-2 family include the proapoptotic protein Bax and the antiapototic protein Bcl-2. Bcl-2 and Bax form homodimer with itself or heterodimer with each other, though sharing the highly conserved BH1 and BH2 domains, and regulate programmed cell death or antiapoptosis.
The recent discovery of RNA interference (RNAi), a natural process through which the expression of a targeted gene can be knocked-down with high specificity and selectivity, presents an invaluable tool for personalized cancer therapy. 19 The applications of RNAi can be mediated through two types of molecules: chemically synthesized double-stranded small-interfering RNA (siRNA) or vector-based short-hairpin RNA (shRNA). In current study, endogenous PLCɛ expression was inhibited by short-hairpin RNA (shRNA) to determine the role of PLCɛ in bladder cancer apoptosis. Results showed that knockdown of endogenous PLCɛ expression induced bladder cancer cell apoptosis. The apoptosis was further characterized by expression of proapoptotic protein Bax and the antiapototic protein Bcl-2. The results further indicate that knockdown of PLCɛ-induced cell apoptosis may associate with shifting the Bcl-2/Bax ratio.
Materials and Methods
Cell lines and culture conditions
The bladder cancer cell lines BIU-87 and T24 were obtained from the China Center For Type Culture Collection (Wuhan University) and the Research Institute of Ultrasound, Chongqing University of Medical Science, respectively, and were cultured in RPMI-1640 (Gibco) containing 10% bovine calf serum (Gibco), 100 units/mL penicillin, and 100 units/mL streptomycin at 37°C under 5% CO2 and 95% humidified air.
shRNA preparation
Based on the PLCɛ cDNA sequence (Gene Bank ID: NM016341), specific shRNAs targeting PLCɛ were designed. The shRNA transcription template synthesis was proceeded by Wuhan Genesil Biotechnology Company. The sequences used were F: 5′-GATCCGCACATACTGTCAGAVGAAGTTTCAAGACGATTCGTCTGACAGTATGTGCTTTTTTGTCGACA-3′; R: 3′-GCGTGTATGACAGTCTGCTTCAAAGTTCTGCTGAAGCAGACTGTCATACCGAAAAAACAGCTGTTCGA-5′ (pGenesil- PLCɛ). Meanwhile, we designed a pair of nonspecificity sequences for a negative blank control (pGenesil-NP), F: 5′-GATCCACTACCGTTGTTATAGGTGTTCAAGACGCACCTATAACAACGGTAGTTTTTTTGTCGACA-3′; R: 3′-GTGATGGCAACAATATCCACAAGTTCTGCGTGGATATTGTTGCCATCAAAAAAACAGCTGTTCGA-5′. Finally, the construction of plasmid was verified by sequencing and identified by restriction endonuclease digestion.
Transfection of shRNA
One day before transfection, BIU-87 and T24 cells were seeded in a growth medium without antibodies so that the cells will be 90%–95% confluence at the time of transfection. Compounds of plasmids and liposome were transfected with the Lipofectamine 2000 (Invitrogen) transfection reagent at the ratio of 1:2. Vector (4 μg) and 8 μL of Lipofectamine 2000 were diluted into a serum-free medium, mixed, and then incubated for 20 minutes at room temperature. The mixture was then added to cells; after 6 hours of incubation, the mixture was replaced with a full medium. Cells were harvested after 24 hours of transfection. Transfection efficiency was observed using a fluorescence microscope.
Total RNA extraction and RT-PCR
BIU-87 and T24 cells were lysed directly in the culture wells, and total RNA was extracted using Trizol reagent (Invitrogen) according to the manufacturer's instructions. RNA was eluted into RNase-free water; yield and quality of the RNA were assessed by measuring absorbance at 260 and 280 nm, followed by electrophoresis on 1% agarose gels, and stored at −80°C until use. Total RNA aliquots (0.5 μg) were used for reaction. RT-PCR was preformed with the TaKaRa RNA PCR Kit (AMV) Ver.3.0 (Takara Bio). Sequence-specific primers for PLCɛ, Bcl-2, Bax, and the housekeeping gene β-Actin, and GAPDH were used for cDNA amplification (see Table 1). RT-PCR was performed as follows: cDNA amplification was initiated after heat denaturation at 95°C for 5 minutes. Thermal cycling conditions were as follows: 95°C for 30 seconds; various annealing temperatures for each gene for 30 seconds (Table 1); and 72°C for 30 seconds, with a final extension step at 72°C for a 7-minute PCR. Products were visualized on 1.5% agarose gels.
Cell cycle assay
Cell cycle was analyzed by flow cytometry (FCM). After 48-hour transfection with pGenesil-PLCɛ, cells were digested to centrifuge tubes, and then washed with a phosphate buffer solution (PBS) for three times and fixed in 70% ethanol overnight at 4°C. Cells were washed with PBS again and then resuspended in the PBS containing 0.2 mg/mL RNase A, 1%v/v Triton X-100, and 20 mg/mL PI at 37°C for 15 minutes. Cells were analyzed for flow cytometry by an FACScan flow cytometer (Becton Dickinson).
Cell apoptosis assay
Cell apoptosis was reflected by flow cytometry and transmission electron microscopy. Cells were digested in centrifuge tubes after 48-hour transfection, and then centrifuged at 2000 rpm for 20 minutes at 4°C; the supernatant was removed; and the cells were fixed in 4% glutaraldehyde for 2 hours. Then, the cells were washed with PBS, and fixed using 1% osmic acid 1 hour, and dehydrated in acetone. Specimens were embedded in Epon, and a thin section was cut using ultramicrotome and double stained with uranyl acetate and lead citrate. Transmission electron microscopy (Hitachi) was performed. Meanwhile, cells were cultured in six-well plates; 48 hours after transfection, cells were removed from the plate using a trypsin digestion solution, collected, and resuspended in 400 μL 1×binding buffer. After the addition of 5 μL Annexin V-FITC to each well, cells were incubated in the dark for 5 minutes. An aliquot of 10 μL PI was added to each well, followed by an additional incubation in the dark for 5 minutes, and finally flow cytometry was performed.
Western blot analysis
To detect protein expression levels, cells were detached using cell scrapers and washed thrice with ice-cold PBS, and then lysed in a lysis buffer (0.5% deoxycholate, 0.1% sodium dodecyl sulfate (SDS), 1% Triton X-100, 10% glycerol, 50 mM Tris, pH 7.5, 150 mM NaCl, 2 mM EDTA, with 1 mM phenylmethyl-sulfonyl fluoride, and 10 μg/mL aprotinin) for 30 minutes in an ice bath. The supernatant was obtained by centrifugation at 13,000 g for 30 minutes at 4°C. Protein samples were electrophoresed on 8%–12% SDS–polyacrylamide gel electrophoresis gels and transferred to polyvinylidene fluoride membranes (Amersham Pharmacia Biotech). Nonspecific reactivity was blocked in 5% nonfat dry milk in TBST for 3 hours at room temperature. The membrane was then immunoblotted with a blocking solution, containing anti-PLCɛ (1:200 dilution; Santa Cruz Biotechnology), anti-Bax (1:5000; Abcam), and anti-Bcl2 (1:5000; Abcam) antibody. Then, the membranes were incubated with a blocking solution containing a 1:5000 dilution of horseradish peroxidase-conjugated secondary antibodies (Santa Cruz Biotechnology). Each sample was also probed with a β-actin antibody (Santa Cruz Biotechnology) as a loading control.
Statistical analysis
All data are presented as mean±standard deviation (SD); statistical significance was evaluated using t-test and one-way completely randomized variance analysis (ANOVA) using SPSS13.0 software. Independence/dependence between characteristics was tested at a significance level of p<0.05.
Results
Inhibition of PLCɛ expression by pGenesil-PLCɛ
The RT-PCR analysis of BIU-87 and T24 RNA detected expression of PLCɛ mRNAs after transfection with the pGenesil-PLCɛ plasmid (Fig. 1A). Under analysis with the Bio-Rad image system, expression of PLCɛ mRNAs in positive plasmids was lower than in the blank control and NP groups, and the difference was significant (p<0.01). Relative to the NP control group, the inhibition ratio was 76.0% for T24 and 78.8% for BIU-87. Expression of the internal standard β-actin did not differ significantly between two groups (p>0.05). As for protein levels (Fig. 1B), the optical density value for the bands for each of the groups was compared with that of β-actin. The ratios of pGenesil-NP groups were apparently lower than for the other two groups, and the difference was significant (p<0.01). The inhibition ratio was 65.4% for the T24 cells and 64.8% for the BIU-87 cells. These data show that the recombinant plasmid specifically inhibited the expression of PLCɛ in T24 and BIU-87 cells.
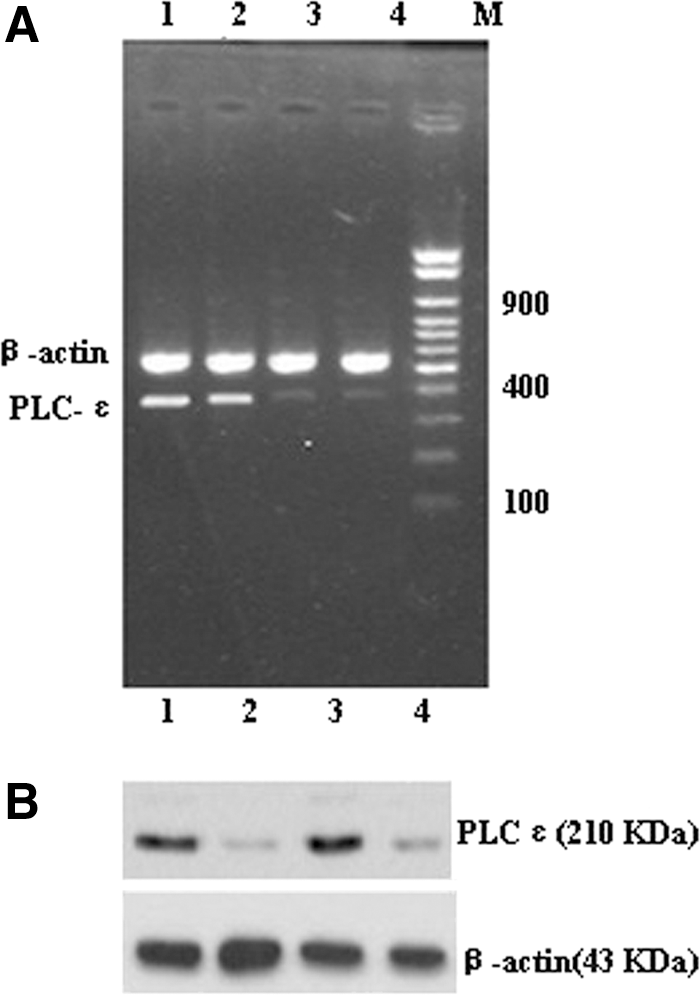
Effect of pGenesil-PLCɛ on the expression of PLCɛ. mRNA and protein were extracted after transfected with the pGenesil-PLCɛ and negative plasmids for 24 hours.
Cell cycle was arrested at the G0/G1 phase
Cell cycle was analyzed using FCM. The representative histograms of the cell cycle distribution are shown in Figure 2A and B; the cell percentage in each phase is summarized in Figure 2C. Cells transfected with pGenesil-PLCɛ resulted in a statistically significant increase in the G0/G1 phase, and accompanied a decrease in the G2/M phase and the S phase in contrast with pGenesil-NP cells. Furthermore, cells were found to contain a sub-G1 amount of DNA after pGenesil-PLCɛ transfection, which indicate a pGenesil-PLCɛ competent to induce apoptosis.

Effect of pGenesil-PLCɛ on cell cycle analysis in BIU-87 cells. Cells were collected after transfected with the pGenesil-PLCɛ and negative plasmids for 48 hours, and then analyzed by flow cytometry.
Effects of pGenesil-PLCɛ on Bcl-2 and Bax expression
The expression of Bcl-2 and Bax was determined by RT-PCR and Western blot. As shown in Figure 3, the Bcl-2 mRNA level was significantly downregulated by pGenesil-PLCɛ transfection (Fig. 3A); in contrast, the Bax mRNA level was upregulated (Fig. 3B). We also found that the ODBcl-2/ODBax ratio tends to decrease (Fig. 3D). As is expected, the protein level of Bcl-2 and Bax changed in concert with changes in mRNA levels (Fig. 3C). Therefore, we believe that the regulation of Bax and Bcl-2 expression after transfection of pGenesil-PLCɛ is partly involved in the induction of apoptosis; *p<0.05, as compared to the pGenesil-NP groups.
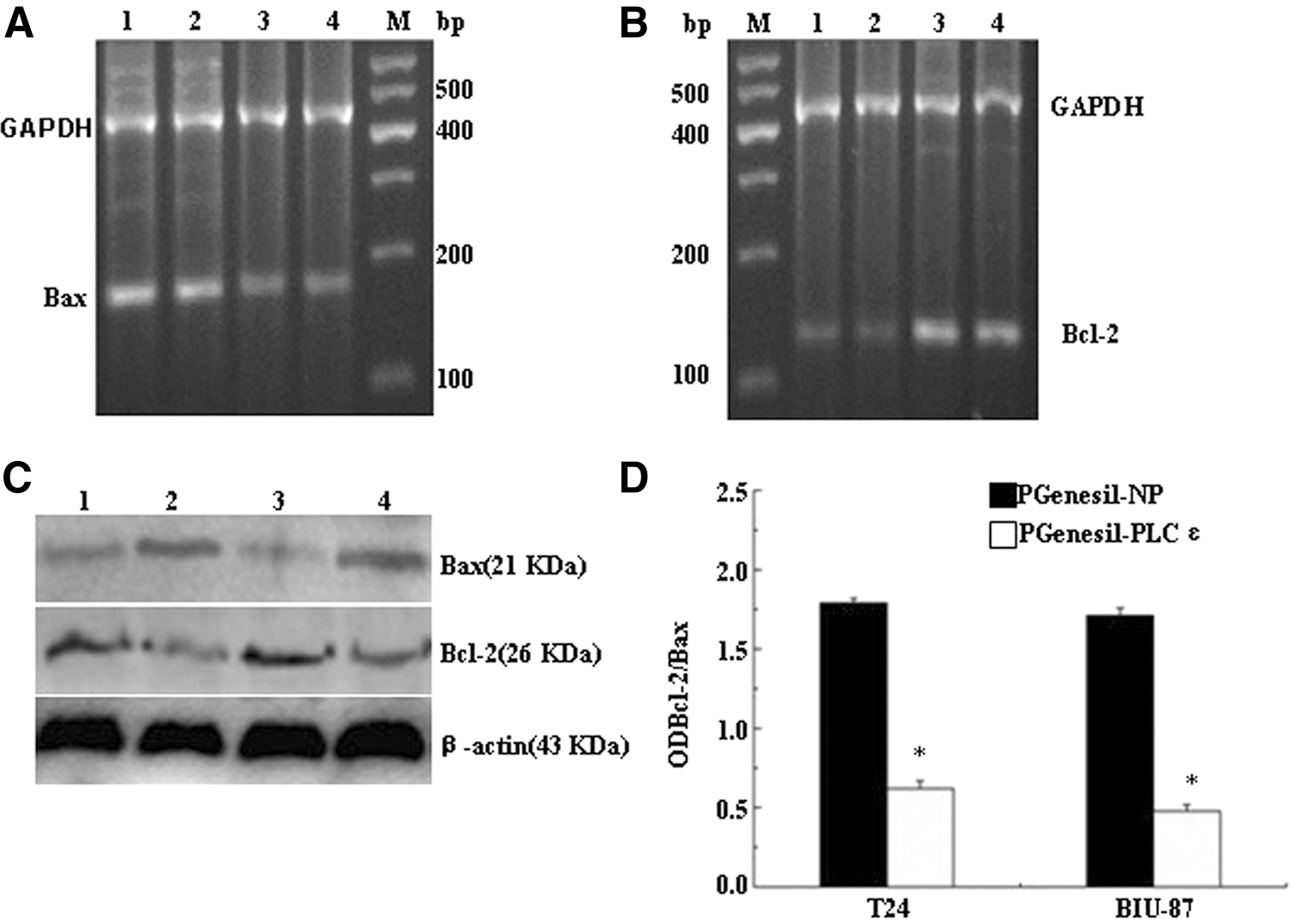
Effect of pGenesil-PLCɛ on expression of Bcl-2 and Bax.
pGenesil-PLCɛ transfection induces apoptosis in bladder cancer cells
Cell apoptosis was examined by the PI method and transmission electron microscopy. Flow cytometry demonstrates that the apoptosis levels of bladder cancer were significantly increased by pGenesil-PLCɛ transfection at a 48-hour time point in both T24 (Fig. 4A, B) and BIU-87 (Fig. 4C, D) cells. TEM is considered the gold standard to confirm apoptosis, as categorization of an apoptotic cell is irrefutable if the cell contains certain ultrastructural morphological characteristics. As shown in Figure 5A, the control cells showed an irregular shape. The cells contained normal-looking nuclei and chromatin. However, in the pGenesil-PLCɛ group (Fig. 5B, C), typical morphological changes can be observed. These results confirmed that PLCɛ plays an important role in inducing apoptosis of T24 and BIU-87 cells.
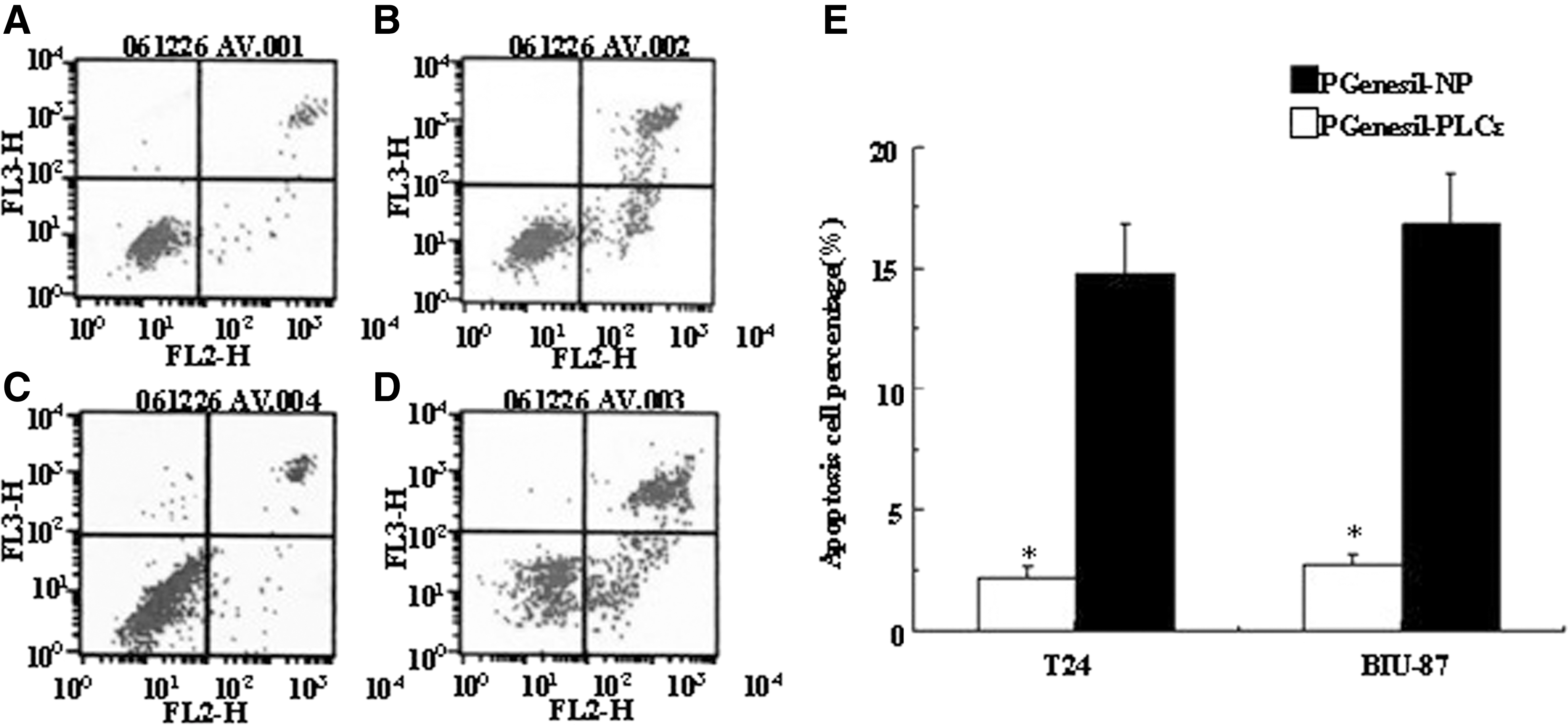
Effect of pGenesil-PLCɛ on apoptosis in bladder cancer cell. Cells were transfected with pGenesil-PLCɛ for 48 hours, and the negative plasmid-transfected cells were used as control. Then, quantitation of apoptosis was analyzed by flow cytometry.

Effect of pGenesil-PLCɛ transfection on cell ultrastructure change.
Discussion
The association between mutation in the RAS gene family (KRAS, HRAS, and NRAS) and 30% of all human cancers, including bladder cancer, suggests that an aberrant RAS function plays an important part in cancer development. Some studies proved the concept that activation of the HRAS oncogene by point mutation and the activation of all RAS genes (mainly NRAS and KRAS) by overexpression are frequent events in bladder cancer cases.
PLCɛ was revealed to be downstream of Ras oncogenes. It has been reported that PLCɛ involved in skin tumor 20 and head and neck squamous cell carcinoma development. 11 Furthermore, PLCɛ could inhibit epidermal growth factor receptor (EGFR) downregulation, 21 and this process may induce an abnormal increase of EGFR, which may lead to oncogenesis. On the basis of such findings, we propose that there exists a previously unrecognized role for PLCɛ as an important factor involved in regulating bladder cancer progression. In our previous study, we demonstrate that PLCɛ plays a crucial role in bladder cancer, 22 and this result was confirmed in this study.
Apoptosis is cell suicide that is induced by various stimuli, and it is important in physiological functions of aging cell clearance and homeostasis maintenance. Furthermore, it is closely related with immune response, regulation of the hematopoietic system, and embryonic development. These lead it to play an important role in tumor development. Generally, it is assumed that tumor is induced by multiple gene changes, leading to continuous cell proliferation or apoptosis inhibition. Apoptosis and proliferation are a pair of coexistence of contradictions; when cell proliferation and apoptosis are imbalanced, it will increase the likelihood of cancer.
B-cell lymphoma-/leukemia-2 (Bcl-2) is a member of Bcl-2 family, 18 which includes apoptosis suppressor genes such as Bcl-2, Bcl-xl, Bcl-w, and Mcl-1 and apoptosis-enhancing genes such as Bax, Bcl-xs, and Bak. Most of the Bcl-2 members contain two homologous domains (BH1 and BH2), through which they form homodimer and heterodimer to mediate cell apoptosis. Bcl-2 is considered to be a cell apoptosis-inhibiting gene. Bcl-2 inhibits cell apoptosis, though suppresses caspase activity. 23 Bax is another member of the Bcl-2 family, is a proapoptosis protein of the Bcl-2 family. Bax homodimerizes and forms heterodimers with Bcl-2 in vivo; overexpressed Bax accelerates apoptotic death induced by cytokine deprivation in an IL-3-dependent cell line. Overexpressed Bax also counters the death repressor activity of Bcl-2, and the ratio of Bcl-2 to Bax determines survival or death after an apoptotic stimulus. 24
Our objective in this study was to examine the effects on apoptosis of specific inhibition of the PLCɛ expression in tumor cells, and to determine if PLCɛ is a valid therapeutic target. Treatment of human bladder cancer cell lines with PLCɛ-shRNA resulted in morphologic and biochemical changes. We detected an apoptotic peak in the pGenesil-PLCɛ transfection group by cell cycle assay. Therefore, we have speculated that cells grow slowly might be partly due to cell apoptosis; consistent with this idea, it was shown that the apoptosis level was increased after pGenesil-PLCɛ transfection.
Our data also showed that Bcl-2 was downregulated after pGenesil-PLCɛ transfection. On the contrary, Bax was upregulated, and the ratio of Bcl-2 to Bax was decreased, which lead to the decrease in the formation of heterodimer of Bcl-2/Bax and increase in the formation of homodimer of Bax/Bax. Previous studies demonstrated that a low ratio of Bcl-2/Bax promotes sensitivity of chemotherapy in cancer therapy. 25,26
In summary, overexpression of PLCɛ promotes cell proliferation and inhibits cell apoptosis, and this biofunction promotes cancer formation and development. Thus, we suspect that it can inhibit or postpone bladder cancer development, though suppresses the overexpression of the PLCɛ gene. Our findings provide novel insights for the development of the gene therapy technology to treat patients with bladder cancer.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (No. 81072086).
Disclosure Statement
No competing financial interests exist.