
Abstract
The aberrant activation of the JAK2/STAT3 signaling in immature myeloid dendritic cells (DCs) is associated with immune tolerance and poor antitumor immunity. The objective of this study was to test the hypothesis that Cucurbitacin B (CuB), a selective inhibitor of JAK2/STAT3 signaling, could promote DC differentiation and improve antitumor immunity. Twelve patients with advanced lung cancers were treated orally with CuB daily for 7 consecutive days. The frequency of peripheral blood myeloid DCs and immature myeloid cells (imCs) in those patients and healthy controls was characterized longitudinally by flow cytometry. The effect of CuB on the differentiation of DCs and p53-specific T responses was evaluated in vitro. The percentages of Lin−DR−CD33+ imCs and Lin−DR+CD33+ DCs were significantly different between patients with lung cancers and healthy controls (1.55% vs. 0.82%, p=0.002; 0.60% vs. 1.90%, p=0.000). Treatment with CuB significantly increased the frequency of Lin−DR+CD33+, but reduced the frequency of Lin−DR−CD33+ in patients with lung cancers (p<0.05). Treatment with CuB induced the differentiation of DCs cocultured with tumor cells 16HBE/BPDE and enhanced the sensitivity of 16HBE/BPDE cells to p53-specific CTL by inhibiting the JAK2/STAT3 activation, but also enhancing the interferon-γ-related STAT1 activation in 16HBE/BPDE cells. CuB significantly reduced the frequency of imCs in patients with lung cancers and enhanced the effect of p53-specific CTL on tumor 16HBE/BPDE cells.
Introduction
Lung cancer is the leading cause of cancer-related mortality worldwide. 1 Clinically, surgery, radiation, and chemotherapy have been used for treating patients with lung cancer. However, 50% of patients have a cancer relapse after surgery, and less than a quarter of patients respond to systemic chemotherapy. 2 Consequently, the long-term survival for patients with lung cancer remains low. Therefore, the development of therapeutic reagents is of great significance.
The p53 gene has been shown to be frequently mutated in a wide range of human cancers, including lung cancer, 3 and has long been viewed as an ideal target for cancer vaccines and immunotherapy. 4 Nikitina et al. 5 engineered dendritic cells (DCs) expressing human wild-type p53 to stimulate T lymphocytes from cancer patients and demonstrated a specific CTL response to cancer cells expressing mutant p53. In addition, injection of DCs that have been transduced with full-length wild-type p53 not only inhibits the growth of xenograft tumor but also minimizes the established tumor in mice. 6 However, the efficacy of the p53-based cancer vaccines in humans is quite limited. Although vaccination with p53 peptides or a DNA vaccine containing the wild-type p53 gene induces a specific immune response in some patients, no objective clinical responses have been observed. 7 –9 A recent report showed that about half of the patients with advanced small-cell lung cancer did not respond to the p53-based DC vaccine, 10 which might be associated with the high frequency of immature myeloid cells (imCs).
Effective immune response depends on the adequate function of bone marrow-derived antigen-presenting cells. Defective differentiation of DCs and accumulation of imCs are hallmarks of lung cancers. 11 –14 The decreased number of mature DCs in tissues impairs the immune system's ability to induce and maintain an effective antitumor immune response. On the other hand, the accumulation of immature DCs (imDCs) and their immunosuppressive nature are responsible for immune tolerance and tumor-associated immune suppression. 14 Hence, these have been thought to be important mechanisms that compromise the effectiveness of cancer vaccines.
The Janus family of tyrosine kinases (JAKs) and the STAT family of transcription factors are critical components of diverse signal transduction pathways that are actively involved in cellular survival, proliferation, differentiation, and apoptosis. JAKs are constitutively associated with many cytokine and growth factor receptors. 15 STAT3 is found constitutively tyrosine phosphorylated and activated in many human cancers. 16 –18 The JAK/STAT pathway is important for normal cell differentiation. Hyperactivation of JAK2/STAT3 induced by tumor-derived factors is responsible for abnormal myeloid cell differentiation in cancer. 19,20 Recently, Nefedova et al. 21 show that inhibition of JAK2/STAT3 signaling dramatically improves DC differentiation and eliminates immunosuppressive myeloid cells in cancer.
Notably, Cucurbitacin B (CuB), an extract of the stem-end of Cucumis melo L, can inhibit the activation of JAK2 and STAT3. 22 In this study, we tested the hypothesis that CuB could promote DC differentiation and improve antitumor immunity. We found that treatment with CuB significantly reduced the frequency of imCs in patients with lung cancer and increased the sensitivity of HBE/BPDE cells to cytotoxic T cells, which was associated with inhibition of JAK2/STAT3 activation, but enhancing interferon (IFN)-γ-mediated STAT1 activation and DC differentiation. We discussed the implications of our findings.
Materials and Methods
Patient characteristics and treatment summary
The experimental protocol was reviewed and approved by the Local Medical Ethics Committee. The inclusion criteria included individual patients at 18 years of age or older with a histologic diagnosis of lung cancer and adequate organ function (WBC, >3000/mm3; ANC, >1500/mm3; platelets, >100,000/mm3; hematocrit, >25%; bilirubin, <2.0 mg/dL; and creatinine, <2.0 mg/dL). Patients with a pre-existing autoimmune disorder, an immunodeficiency condition, an ongoing infection, or a use of immunosuppressive drugs, such as steroids, were not eligible. Patients were excluded if they received any anticancer therapy concomitant with CuB. Written informed consent was obtained from individual patients and healthy subjects.
A total of 27 patients with lung cancers and 24 healthy subjects were recruited. Individual patients with lung squamous cell carcinomas or adenocarcinoma were diagnosed histologically. Their demographic and clinicopathological characteristics are summarized in Table 1. Among the patients, 12 patients who had not been treated with radio- and/or chemotherapy were treated orally with 120 μg CuB (Chongqing Bei-Lin Pharmaceutical) three times per day for consecutive 7 days. The patients were evaluated, and their blood samples were collected before and after treatment. One part of the blood sample was prepared for sera, and the remaining blood samples were characterized for the frequency of DCs and imDCs by a flow cytometry analysis.
Cell lines and cultures
Human bronchial epithelial SV40-transformed cells (16HBE) were kindly provided by Prof. Steven Holgate from the Medical School of University of Southampton, United Kingdom. The tumor cell line 16HBE/BPDE was provided by Prof. Yiguo Jiang from the Institute for Chemical Carcinogenesis of Guangzhou Medical College. 16HBE/BPDE cells are transformed by benzo(a)pyrene metabolite—dihydroxyepoxy benzopyrene (BPDE)—and can form solid squamous carcinomas in nude mice. Those cells were maintained in an MEM supplemented with 10% fetal bovine serum (FBS; Gibco).
Cell isolation
Human peripheral blood mononuclear cells (PBMCs) were isolated from peripheral blood samples of healthy volunteers by Ficoll-Hypaque (d=1.077 g/mL) density-gradient centrifugation. The collected PBMCs at 5×106/mL were cultured in an RPMI 1640 medium supplemented with 10% heat-inactivated FBS for 2 hours at 37°C in 5% CO2. The adherent cells were stimulated with GM-CSF (1000 units/mL) and interleukin (IL)-4 (500 units/mL; Peprotech) for 6 days and used as imDCs. To obtain mature DCs, the cells were treated with PGE2 (1 μg/mL; Sigma) and TNF-α (20 ng/mL; Peprotech) for an additional 48 hours. The nonadherent cells were harvested and incubated with anti-CD22 and anti-CD56 microbeads, followed by magnetic separation. The purity of unbound T cells was determined by the flow cytometry analysis. The obtained T cells with a purity of >94% were used for further experiments.
Flow cytometry
Cells were labeled with FITC-conjugated Lineage cocktail 1 (lin 1) against CD3, CD14, CD16, CD19, CD20, and CD56 (Becton Dickinson). In addition, the cells were stained with FITC-labeled anti-CD80, anti-CD83, Pe-Cy5-labeled anti-HLA-DR, PE-labeled anti-CD33, anti-CD86, anti-CD40, or isotype controls (Becton Dickinson). The cells were fixed with 1% (w/v) paraformaldehyde in phosphate-buffered saline (PBS) for 30 minutes at room temperature and analyzed by flow cytometry on an FACSCalibur flow cytometer (Becton Dickson) using Cell Quest software.
Tumor cell–DC cocultures
The isolated imDCs from healthy subjects were stimulated with GM-CSF (1000 units/mL) and IL-4 (500 units/mL) for 2 days. The cells at 2×105 cells/well were loaded on the top well of a transwell plate (0.2-μm-diameter pore; Corning/Costar) in the presence of CuB (0.03 μg/mL) or the same volume of vehicle dimethyl sulfoxide (DMSO), and 16HBE/BPDE or 16HBE cells at 5×104 cells/well were loaded on the bottom well of the transwell plate in a complete culture medium (CCM) for 4 days. The suspended imDCs were harvested and subjected to a flow cytometry analysis.
Generation of CTLs
imDCs were infected with adenovirus containing the wild-type p53 gene (Ad-p53; SiBiono GeneTech) or the replication-defective control Ad-c at multiplicities of infection of 20,000, stimulated with GM-CSF and IL-4 for 2 hours at 37°C, and exposed to a fresh CCM supplemented with GM-CSF, IL-4, PGE2, and TNF-α. The infected DCs (1.5×106 cells/well) were cultured in the CCM in six-well plates for an additional 48 hours and mixed with 7.5×106 T lymphocytes in the CCM in the presence of 30 units/mL of IL-2 for 5 days. The T cells were collected and re-stimulated with infected DCs for another 5 days. The responder T cells were harvested and assessed for IFN-γ secretion by the ELISPOT assay and cytolytic activity. The supernatants were harvested for testing the concentrations of TNF-α, IFN-γ, IL-4, and IL-10 by Luminex Assays.
IFN-γ ELISPOT assay
The responder DC-p53-T cells, DC-T cells, and freshly isolated control T cells at 1×105 cells/well were stimulated in duplicate with p53-DCs (1×104 cells) at 37°C, 5% CO2 for 24 hours in 96-well plates that had been coated with an anti-IFN-γ antibody using an IFN-γ ELISPOT kit, according to the manufacturer's instructions (Becton Dickinson). The numbers of IFN-γ-secreting T cells were evaluated using an automated ELISPOT reader (BioReader 4000 Pro-X) in a blinded manner.
Western blotting
The relative levels of signaling events in 16HBE/BPDE cells were determined by the Western blot analysis, as previously described. 23 Briefly, 16HBE/BPDE cells were harvested and lyzed in a lysis buffer. After quantification, the protein lysates (30 μg/lane) were separated by SDS-PAGE electrophoresis and transferred onto PVF membranes. After being blocked with 5% fat-free milk, the membranes were incubated with a 1:1000 diluted rabbit monoclonal antibody against human Phospho-STAT1 (tyr701), Phospho-STAT3 (tyr705), Phospho-JAK2 (tyr1007), STAT3, STAT1, or with a mouse monoclonal antibody against human GAPDH (Cell Signaling Technology) at 4°C overnight, respectively. The bound antibodies were detected with 1:1000 diluted HRP-conjugated goat anti-mouse IgG or goat anti-rabbit IgG, and visualized using an ECL chemiluminescent substrate (Amersham), followed by digital scanning in Image Station 2000 (Kodak).
Cytotoxicity assays
Cytotoxicity of the generated CTLs against 16HBE/BPDE was evaluated using a colorimetric cell-counting kit (CCK-8; Dojindo Laboratories), according to the manufacturer's instructions. Briefly, 16HBE/BPDE cells at 1×104 cells/well were cocultured in triplicate with CTLs at the ratio of 1:10, 1:20, and 1:40 for 14 hours at 37°C in 5% CO2 and exposed to 20 μL of CCK-8 solution for 2 hours at 37°C. The resulting formazan was dissolved in DMSO, and the absorbance at 450 nm was measured on a microplate reader. The 16HBE/BPDE cells or CTLs alone were used as controls. The percentage of specific cytotoxicity was calculated as [(effect cell control+target cell control−experimental)/target cell control]×100%. In addition, 16HBE/BPDE cells were first treated with CuB for 12 hours and used as the target cells to examine the effect of CuB on the sensitivity of 16HBE/BPDE cells to CTLs.
Statistical analysis
Data are expressed as mean±standard error of the mean. The difference between two groups was analyzed by unpaired or paired two-tailed Student's t-tests using SPSS for Windows. Statistical significance was defined as a value of p<0.05.
Results
Treatment with CuB modulates high frequency of peripheral blood imCs in patients with lung cancer
To determine the role of myeloid DCs in the development of lung cancers, a total 27 patients with lung cancers and 24 healthy subjects were studied for the frequency of DCs. Their PBMCs were prepared and stained with lineage-specific antibodies, DR-, and CD33-specific antibodies, followed by the flow cytometry analysis. As shown in Figure 1, the Lin− cells were gated in R2, and the Lin−DR+CD33+ DCs and Lin−DR−CD33+ imCs were gated in R5 and R6, respectively. Quantitative analysis revealed that the percentage of Lin−DR+CD33+ DCs in patients with lung cancer was significantly lower than those in controls, whereas the frequency of Lin−DR−CD33+ imCs in patients with lung cancer was significantly higher than those in controls (Fig. 1B). Apparently, a high frequency of imCs was present in patients with lung cancer. Furthermore, treatment with CuB significantly elevated the frequency of Lin−DR+CD33+ DCs, but decreased the percentage of Lin−DR−CD33+ imCs in patients with lung cancer (Fig. 1C). Therefore, treatment with CuB modulated the high frequency of imCs in patients with lung cancer.
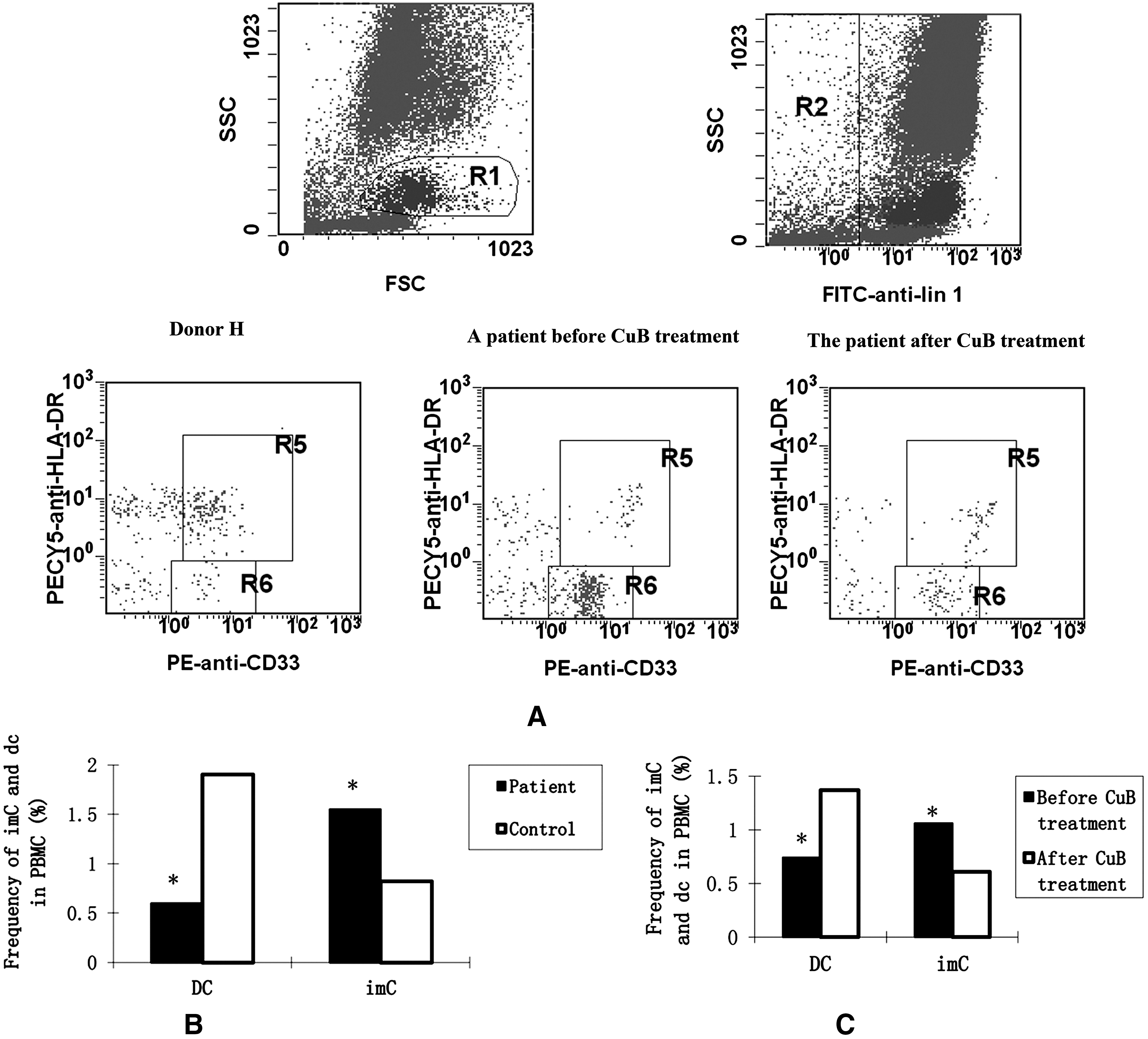
Characterization of lin−DR−CD33+ and lin−DR+CD33+ cells. Peripheral blood samples were obtained from individual patients with advanced lung cancers and healthy controls, and PBMCs were prepared. The PBMCs were stained with a lineage cocktail of FITC-conjugated antibodies against CD3, CD14, CD16, CD19, CD20, and CD56, PEcy5-conjugated anti-HLA-DR, and PE-conjugated anti-CD33 and analyzed by flow cytometry.
Treatment with CuB modulates the sensitivity of 16HBE/BPDE cells to p53-specific CTL
The p53 gene is frequently mutated or inactivated in many types of human lung cancers. 3 Mutations in the p53 gene often result in the overexpression of the p53 protein in tumor cells, although accumulation of p53 may also occur in tumors without mutations in the p53 gene. 24 The 16HBE/BPDE cells have a mutation in the p53 gene, which results in high levels of p53 expression (data not shown). To optimize in vitro induction of CTLs specific for the p53 antigen, imDCs were isolated from healthy donors and infected with Ad-p53 or control Ad-c to generate p53-DCs or control DCs, respectively. Subsequently, T cells from the same donors were stimulated with p53-DCs or control DCs in vitro. The frequency of IFN-γ-secreting activated T cells was analyzed by ELISPOT assays (Fig. 2). Clearly, the numbers of IFN-γ-secreting T cells from cocultured T with p53-DCs were significantly higher than those from cocultured T with control DCs or T cells alone. Analysis of cytokines indicated that there were significantly higher levels of IFN-γ and TNF-α, but not IL-4 and IL-10 in the supernatants of cocultured T with p53-DCs, compared with that in cocultured T with control DCs (Table 2). These two lines of data indicated that p53-DCs induced activation of p53-specific T cells in vitro.
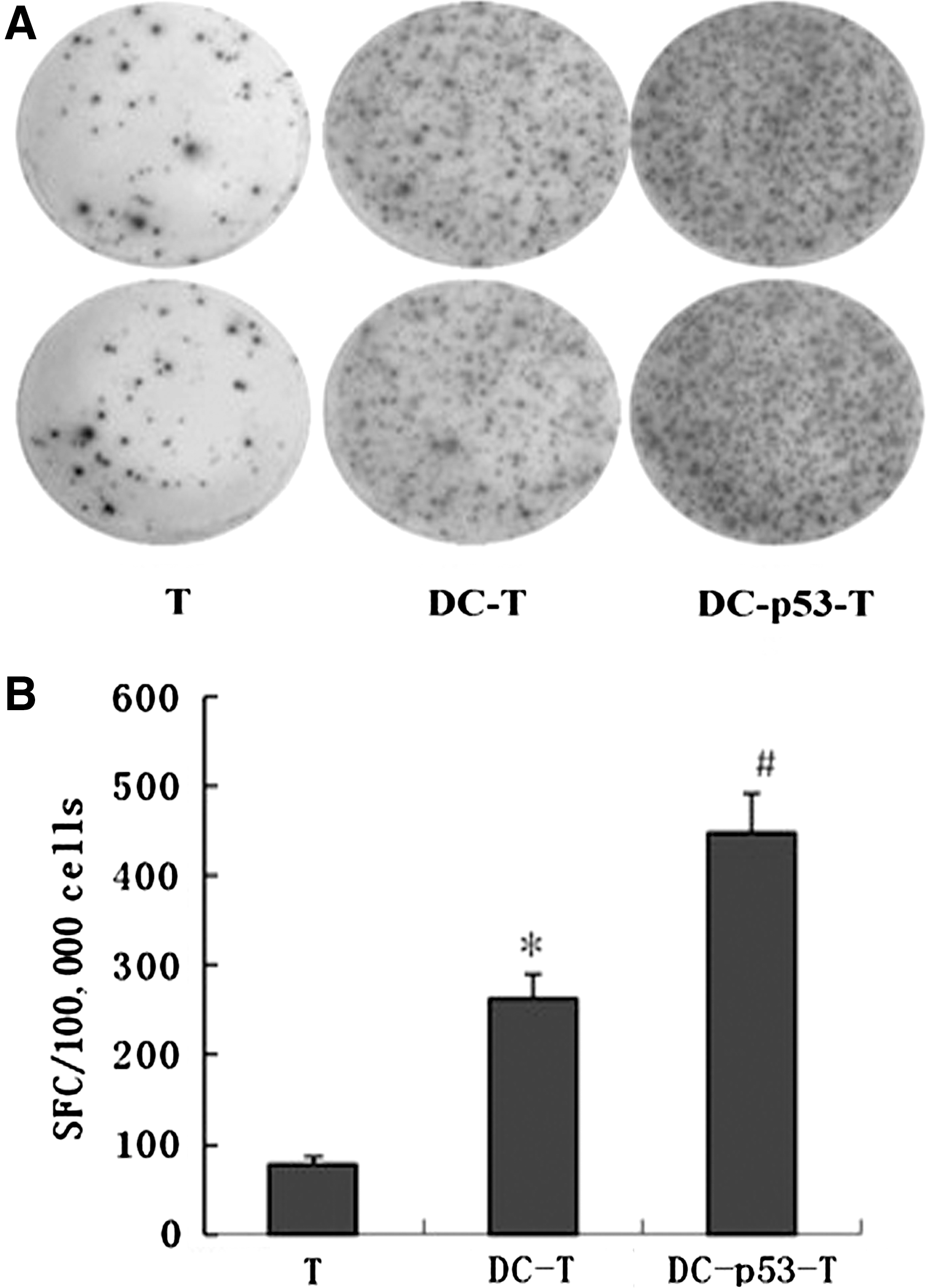
Characterization of IFN-γ-secreting T cells. T cells were purified form healthy subjects and cocultured with Ad-p53 or control Ad-C-infected and cytokine-activated DCs from the same subject in vitro for 10 days. The responder DC-p53-T cells, DC-T cells, and freshly isolated control T cells were stimulated with p53-DCs for 24 hours, and the numbers of IFN-γ-secreting T cells were determined by ELISPOT assays.
Data are expressed as mean±standard error of the mean of each group.
p<0.01 versus the values of c-Dc-T control, determined by the Student's t-test.
DCs, dendritic cells; IFN, interferon; IL, interleukin.
We next examined the cytotoxicity of p53-specific T cells against 16HBE/BPDE cells in vitro by the CCK-8 assay. Unexpectedly, there was no significant difference in the cytolysis between p53-DC-induced T and DC-induced T cells in our experimental system (Fig. 3A). Apparently, 16HBE/BPDE cells were poor targets for p53-specific T cells. Interestingly, the cytotoxic rates of p53-DC-induced T cells against CuB-treated 16HBE/BPDE cells were significantly higher than those against untreated cells (Fig. 3B). Hence, pretreatment of 16HBE/BPDE cells with CuB significantly enhanced the sensitivity of 16HBE/BPDE to p53-DC-induced T cells.

Effect of CuB on the sensitivity of 16HBE/BPDE cells to p53-specific CTLs. T cells were purified from healthy subjects and stimulated twice with Ad-p53-DCs or with Ad-c-DCs for 10 days, respectively. The generated p53-specific activated T cells were mixed with 16HBE/BPDE cells or CuB-pretreated 16HBE/BPDE cells at the indicated ratios, and the cytotoxicity of activated T cells against those tumor cells was determined.
Treatment with CuB modulates the JAK/STAT pathway in 16HBE/BPDE cells
The constitutive activation of the JAK2/STAT3 pathway is a major contributor to oncogenesis in many types of human cancers, which functions as a critical mediator of oncogenic signaling through transcriptional activation of genes encoding apoptosis inhibitors (e.g., Bcl-xL, Mcl-1 and survivin). 25 We next characterized the relative levels of JAK2 and STAT3 expression and phosphorylation in 16HBE/BPDE cells. We found that 16HBE/BPDE cells displayed a high level of JAK2 and STAT3 phosphorylation (Fig. 4A). Notably, CuB has been found to inhibit the JAK2 and STAT3 activation in the human nonsmall-cell lung carcinoma line, 26 and inhibition of the JAK2 and SAT3 pathway should enhance the sensitivity of tumor cells to apoptotic triggers. Indeed, we found that treatment with CuB dramatically reduced the relative levels of p-STAT3 and p-JAK2 in 16HBE/BPDE cells (Fig. 4A), and that the effects of CuB on inhibition of STAT3 phosphorylation were time dependent (Fig. 4B).
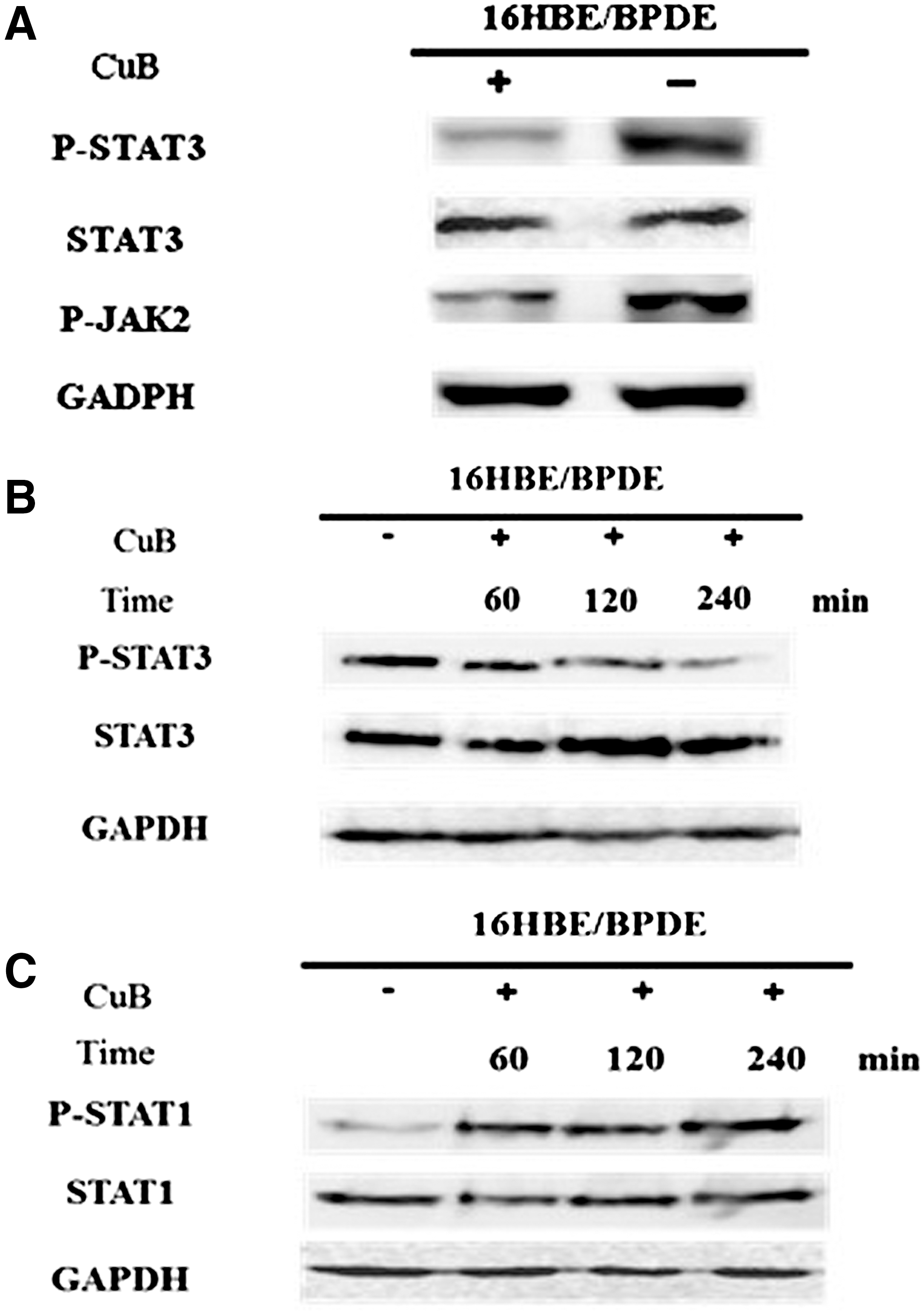
CuB modulates the JAK/STAT signaling in 16HBE/BPDE cells. The 16HBE/BPDE cells were treated with, or without, 0.03 μg/mL of CuB for varying periods, respectively, and the relative levels of JAK2, STAT3, phosphorylated JAK2, phosphorylated STAT3, STAT1, and phosphorylated STAT1 were characterized by Western blot assays using specific antibodies.
IFN-γ is one of the important cytokines secreted by CTL, and STAT1 is the major effector of the IFN-γ signaling, 27,28 contributing to the cytotoxicity of CTLs. Next, we examined whether treatment with CuB could modulate the IFN-γ-induced STAT1 activation in 16HBE/BPDE cells. We found that, while treatment with IFN-γ alone slightly increased the relative levels of phospho-STAT1, treatment with both IFN-γ and CuB dramatically elevated the relative levels of phospho-STAT1 in 16HBE/BPDE cells in vitro (Fig. 4C). Therefore, CuB enhanced the sensitivity of 16HBE/BPDE cells to CTL-mediated cytotoxicity by downregulating the JAK2/STAT3 signaling, but enhancing the IFN-γ-induced STAT1 activation in 16HBE/BPDE cells.
CuB induces the differentiation of DCs cocultured with tumor cells
To evaluate the possible effects of CuB on the differentiation of DCs, the PBMC-derived DCs were cocultured with tumor cells 16HBE/BPDE in a transwell system for 4 days in the presence or absence of CuB, and the DCs were analyzed by the flow cytometry analysis (Fig. 5). In comparison with the DCs cocultured with 16HBE/BPDE in the absence of CuB, the DCs cocultured with 16HBE/BPDE in the presence of CuB displayed a significant increase in the relative levels of CD80, CD83, CD40, and CD86 expression (Fig. 5), suggesting that CuB promoted the differentiation of DCs in vitro by reducing inhibitory factors produced by 16HBDE/BPDE. Therefore, CuB antagonizes the inhibition of tumor cells to promote DC differentiation in our experimental system.
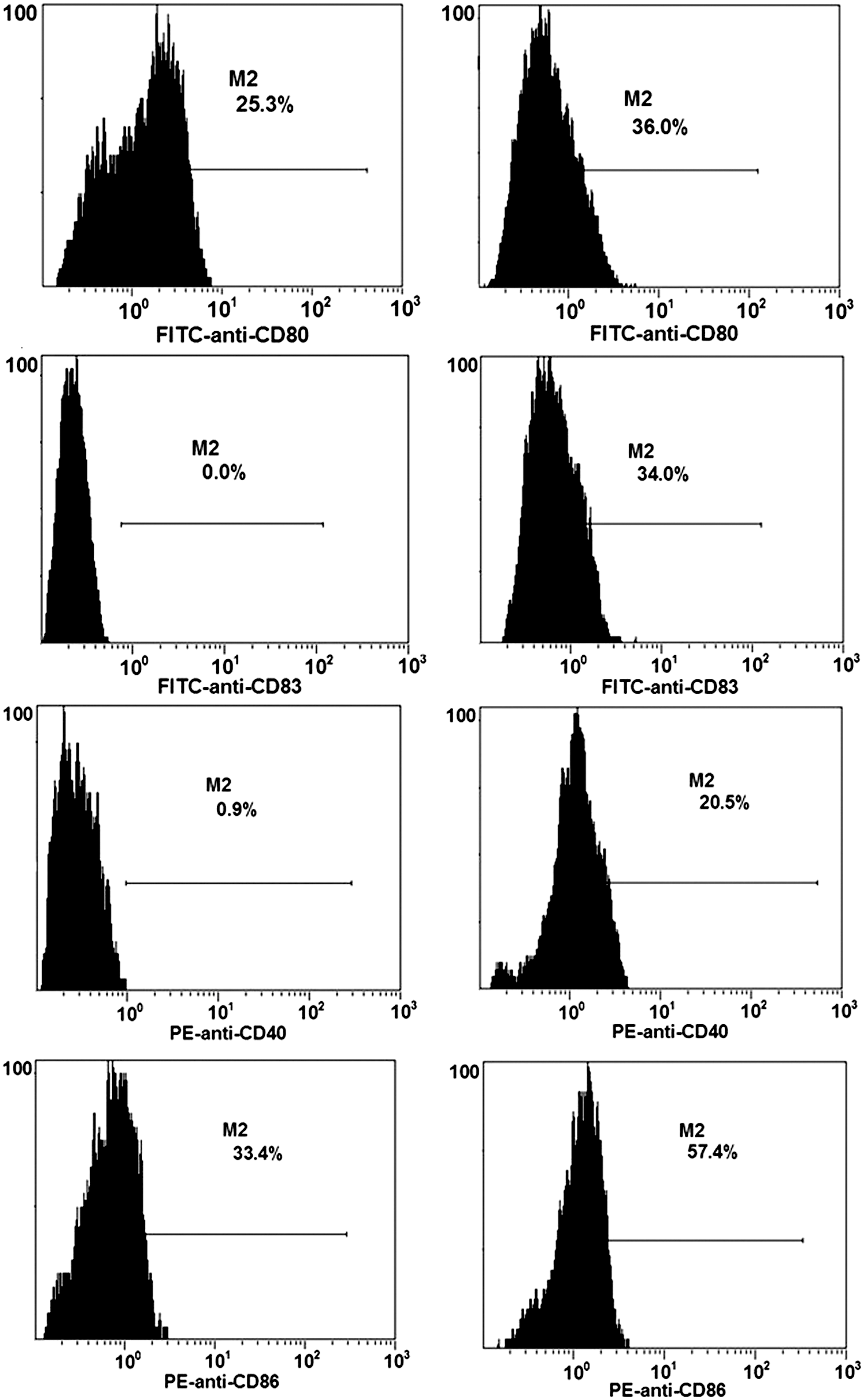
CuB modulates the 16HBE/BPDE cell-induced inhibition of DC differentiation in vitro. PBMCs were isolated from healthy controls and stimulated with GM-CSF (1000 units/mL) and interleukin-4 (500 units/mL) for 6 days. Subsequently, the suspended cells were cocultured with 16HBE/BPDE in the presence (right) or absence (left) of CuB (0.03 μg/mL) for 4 days, respectively. The levels of CD80, CD83, CD40, and CD86 expression on DCs were characterized by a flow cytometry analysis. Data shown are representative histograms of each group from three separate experiments.
Discussion
Defective differentiation and maturation of DCs are hallmarks of lung cancers. 11 –13 In this study, we determined the frequency of peripheral blood immature and mature DCs and found that there was a higher frequency of peripheral blood Lin−DR−CD33+ imCs and lower percentages of Lin−DR+CD33+ DCs in patients with advanced lung cancers. Our data were consistent with a previous observation. 29 Apparently, low frequency of mature DCs may contribute to the defective antitumor immunity in patients with advanced lung cancers.
Aberration of the JAK2/STAT3 pathway promotes tumor cell proliferation, and a previous study has suggested that inhibitors for the JAK2/STAT3 signaling may be used for promoting the efficacy of cancer immunotherapy. 21 CuB is a selective inhibitor of JAK2/STAT3 signaling. In this study, we investigated whether treatment with CuB could modulate the tumor-associated immune suppression. We found that oral treatment with CuB significantly increased the frequency of Lin−DR+CD33+ DCs, but decreased the percentage of Lin−DR−CD33+ imDCs in patients with advanced lung cancers. Furthermore, CuB appeared to modulate inhibitory factors produced by tumor cells to promote the differentiation and maturation of DCs in vitro. Evidentially, while DCs cocultured with 16HBE/BPDE tumor cells displayed lower levels of co-stimulator molecules, CD80, CD83, and the MHC class II molecule HLA-DR, than those cocultured with control 16HBE cells, addition of CuB significantly increased the levels of CD80, CD83, CD86, and HLA-DR expression in DCs cocultured with 16HBE/BPDE tumor cells in a transwell system. It is possible that CuB modulated tumor cells and reduced inhibitory factors' production to promote the differentiation and maturation of DCs, as human tumor cells can secrete inhibitory factors that suppress immune responses. 30 Alternatively, CuB may directly affect DCs to promote their differentiation and maturation. We are interested in further investigating the mechanisms by which CuB modulates the differentiation and maturation of DCs.
A previous study has shown that DCs transduced with wild-type p53 can induce cytotoxic T cells specifically against p53-overexpresing tumors. 5 We found that although higher frequencies of IFN-γ-secreting T cells were induced by p53-DCs in vitro, they failed to recognize the p53-mutated 16HBE/BPDE tumor cells. Instead, the induced p53-specific CTL had potent cytotoxicity against other p53-mutated H460/cDDP tumor cells (data not shown). More interestingly, treatment with CuB significantly increased the cytotoxicity of p53-specific CTLs against 16HBE/BPDE cells in vitro. These data suggest that aberration of JAK2/STAT3 signaling in 16HBE/BPDE cells may result in the insensitivity of 16HBE/BPDE cells to p53-specific CTLs or cytotoxic cytokines, such as IFN-γ and TNF-α. Indeed, IFN-γ is one of the most abundant cytokines secreted by CTL, and IFN-γ can induce tumor cell apoptosis through the STAT1 pathway. 31 Consistent with the notion, we found that treatment with CuB increased the STAT1 activation in 16HBE/BPDE cells. Hence, it is possible that CuB may upregulate the STAT1 expression and increase the sensitivity of 16HBE/BPDE cells to the cytotoxicity of p53-specific CTLs.
The natural CuB constitutes a group of diverse triterpenoid substances, which are well known for their bitterness and toxicity. Structurally, they are characterized by the tetracyclic cucurbitane nucleus skeleton, namely 19-(10→9b)-abeo-10alanost-5-ene (also known as 9b-methyl-19-nor lanosta-5-ene), with a variety of oxygenation functionalities at different positions. 32 CuB has been used traditionally to treat liver cancer and hepatitis for more than 20 years in China. Our data indicated that CuB promoted the differentiation of imDCs and enhanced the cytotoxicity of p53-specific CTLs against tumor cells in vitro. More importantly, treatment with CuB significantly increased the frequency of mature DCs in patients with advanced lung cancer. Therefore, our findings suggest that pharmacological inhibition of the JAK2/STAT3 pathway may be a useful complementary tool in cancer immunotherapy, and that CuB may be valuable for both modulating the sensitivity of tumor cells to CTLs and enhancing antitumor immunity in the clinic.
Footnotes
Disclosure Statement
The authors declare that they have no conflicts of interest.