
Abstract
Type III interferon (IFN-λ) is a novel member of the interferon family, which preferentially promotes antiviral responses from epithelial cells and cooperates with type I IFNs in the clearance of viral infections. However, the effect of mIFN-λ2 to the LA795 lung adenocarcinoma cell is largely unknown. In this study, we transfected Ad-mIFN-λ2 vector into LA795 tumor-bearing mice to explore the effect of mIFN-λ2 on the proliferation of LA795 lung adenocarcinoma cell and on the immune response of the mice. Transfected by Ad-mIFN-λ2 vector, a significant decrease in the tumor growth, the subcutaneous tumor necrosis, cystic degeneration, and tumor apoptosis were more evident; at the same time, mIFN-λ2 protein and gene were significantly more expressed. And, flow cytometry analysis suggested that CD3+CD4+, CD3+CD8+, and NK (CD3−CD49+) cells were all significantly increased after transfected by Ad-mIFN-λ2. The study demonstrated that recombinant Ad-mIFN-λ2 transfection effectively inhibited the growth of LA795 lung adenocarcinoma cell, which may work through inducing apoptosis of tumor cell and regulating cell immune response.
Introduction
Lung cancer is now the most common and leading cause of cancer death in both men and women all over the world, with over 1,000,000 deaths occurring yearly. 1 The principles of treatment have changed little over the last 50 years, and surgery, radiotherapy, chemotherapy, or a combination of one or all of these remain the treatments of choice. 2,3 Recently, with the development of the society economy and the recognition of the diseases, lots of new therapies were constantly raised, one of which is interferon therapy.
Interferons (IFNs)-λ1, -λ2, and -λ3, also known as IL-29, IL-28A, and IL-28B, respectively, whereas mice only express IFN-λ2 and -λ3, are novel type III IFNs identified and termed recently by two independent research groups. IFN-λs are a functionally nonredundant component of the mucosal antiviral innate immune system. They exhibited an antiviral effect in HT29, A549, and HaCaT cells infected with vesicular stomatitis virus and in HT29 and HepG2 cells infected with encephalomyocarditis virus (EMCV). 4,5 These IFNs must bind to a distinct heterodimeric receptor (IL28R), consisting of the IL28Rα, also called IFN-λ receptor 1 (IFN-λR1), and the IL10Rβ chains, belonging to the class II cytokine receptor family (CRF2). Functional receptors for IFN-λs are preferentially expressed on epithelial cells. Although using distinct receptor complexes, IFN-λs seem to induce intracellular signals similar to those induced by the type I IFNs to promote apoptosis and inhibit cell proliferation by activating caspase-3, caspase-8, and caspase-9. 6,7 Several studies have demonstrated that IFN-λs regulate a range of immune responses through interactions with their corresponding receptors. 8,9
To observe the effects of the mIFN-λ2, it necessitates a vector. A vector is an important DNA carrier that is composed of plasmids (a DNA molecule that occurs inherently in bacteria), liposome (a packaged plasmid DNA similar to a cell membrane), and most often viruses. Vectors are chosen because they are small enough to enter the cell and replicate easily. Adenoviral vectors are the most widely used vector systems for clinical gene therapy against lung cancers. Due to the pneumotropic characteristic of these vectors, the lung cells could be transformed at any stage of the cell cycle and, therefore, the transformation efficiency in the lung tissue could be relatively high. Importantly, there is no risk that adenoviral DNA could integrate into the chromosome of the host. 10
This study aimed to transfect LA795 lung adenocarcinoma cells with Ad-mIFN-λ2 vector and explores the effects of this vector on proliferation and the immune response of LA795 tumor-bearing mice. Given the essential functions of type IFN-λs in the innate immunity, our finding could support it as a new drug of gene therapy systems for the treatment of the lung cancer. And it may be potentially applied to other cancer systems.
Materials and Methods
The recombinant adenoviruses, Ad-mIFN-λ2 and Ad-Lacz, were constructed and stored at the Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences, People's Republic of China. The LA795 lung adenocarcinoma cell line was obtained from the Cell Culture Center of the Basic Institute of Medical Sciences, Peking Union Medical College. Cell culture reagents were obtained from Gibco. All polymerase chain reaction (PCR) primers were synthesized by the Shanghai Sangon Biological Engineering Technology & Services Co. Ltd. TRIZOL reagent was purchased from Invitrogen. PCR master mix was obtained from Toyobo. TdT-mediated dUTP-biotin nick end-labeling (TUNEL) assay kit was purchased from Kaiji BioTech. Anti-IFN-λ2 antibody was purchased from R&D Systems.
Tumor-bearing mice model
Thirty Kunming mice, 4 weeks old and weighing 18–22 g, were purchased from the Experimental Animal Center of Jiangsu University. The LA795 lung adenocarcinoma cells were cultured in DMEM (Gibco-BRL) supplemented with 10% fetal bovine serum, 2 mM
Animal experiment
The mice were ready for transfection when tumors reached 5–20 mm in diameter. The tumor-bearing mice were preinjected with 50 μL lidocaine hydrochloride in the quadriceps femoris. After 15 minutes, each group of mice was injected in situ with the following: 200 μL PBS in the blank group, 3×108 pfu Ad-Null in the Lacz group, and 2×109 pfu Ad-mIFN-λ2 in the Ad-mIFN-λ2 group. Adenovirus transfection was performed once weekly for 3 weeks. The volume of the tumor was measured at days 0, 7, 14, and 21. After 21 days, all mice were anesthetized and killed and tissues were dissected from the tumor, skeletal muscle, lung, and spleen. Some tissues were fixed in 4% paraformaldehyde and others were stored at −76°C for subsequent analysis.
Tumor growth curve and inhibition rates
Once the size of the tumor had reached 5–20 mm in diameter, the short (a) and long (b) diameters of the tumor were measured using vernier caliper every 7 days. The measurement included the skin thickness. The volumes of the tumors for plotting a growth curve were calculated according to the formula V=a 2 ×b×0.52. The tumor inhibition rates were calculated based on the last measurement of tumor volume according to the following formula: tumor inhibition rate=(average tumor volume in control group−average tumor volume in treatment group)/mean tumor volume in control group×100%.
Morphological analysis
Dissected tissues from the tumor, lung, and spleen were fixed with 4% paraformaldehyde for 24 hours, embedded in paraffin, sliced in 5-μm-thick sections, and stained using a hematoxylin and eosin staining procedure. Pathological changes were examined in the tissue sections under an optical microscope and photographed for documentation.
Apoptosis assay
Apoptosis was analyzed using TUNEL assay kit according to the manufacturer's instructions. In brief, 5-μm-thick frozen sections were prepared and fixed in 4% formaldehyde for 20 minutes at room temperature (15°C–25°C). This was followed by rinse with PBS for 30 minutes and incubation with 3% hydrogen peroxide in methanol for 10 minutes at room temperature. After 25-minute washes with PBS, the samples were incubated with 0.1% Triton X-100 and 0.1% sodium citrate in water for 30 minutes at room temperature. For negative control, TdT was not added to the sample, and for positive control, cells were treated by DNase I. After two washes with PBS, pretreated specimens were incubated with 50 μL TdT labeling reaction buffer at 4°C overnight in dark and then in a humidified atmosphere at 37°C for another 2–3 hours. Subsequently, the slides were incubated with 50 μL streptavidin-HRP for 60 minutes, followed by detection with 50 μL diaminobenzidine reagent for 10 minutes. The sections were observed and photographed under an optical microscope.
Western blotting
Western blotting was performed to determine the mIFN-λ2 expression. Tumor tissue (about 1 g) was cut into pieces on ice and homogenized. After rapid centrifugation, the supernatant was discarded and the pellet was resuspended with 1000 μL precooled RIPA lysis buffer, 3 μL phenylmethyl sulfonylfluoride, 3 μL protease inhibitory cocktail, and 3 μL sodium orthovanadate. The mixture was homogenized and lysed for 45–60 minutes on ice. After centrifugation at 12,000 g at 4°C for 15 minutes, the supernatant was transferred into a new 1.5 mL Eppendorf tube and mixed with equal volumes of 2×loading buffer and 1/20 β-mercaptoethanol in boiling water for 5–10 minutes. The extracted proteins were dispensed and stored at −76°C. Extracted lysates were separated on 15% SDS-PAGE gel and mIFN-λ2 expression was confirmed by the indicated primary antibodies followed by HRP-conjugated secondary IgG antibodies. The protein bands were detected by an image analyzer. GAPDH was used as a negative control and internal reference.
Reverse transcription PCR analysis
Tumor and skeletal muscle tissues (50–100 mg) were cut into pieces on ice and homogenized. After centrifugation at 16,000 g at 4°C for 20 minutes, the supernatant was discarded. The total RNA was isolated from the pellet using TRIZOL reagent. The cDNA was synthesized with Oligo(dT) primers and M-MLV Reverse Transcriptase. The 1/10 cDNA was used for PCR amplification. The PCR protocol was as follows: initial denaturation at 94°C for 5 minutes, followed by 35 cycles at 94°C for 30 seconds, annealing at 61°C (mIFN-λ2) or 58°C (β-actin) for 30 seconds, and extension at 72°C for 45 seconds. The final extension was performed by an incubation step at 72°C for 7 minutes. The PCR products were subjected to electrophoresis in agarose gel and visualized with ethidium bromide. The bands were analyzed with Quantity One software (Bio-Rad). All primers that were used are listed in Table 1.
Flow cytometry
Flow cytometry was applied to examine T lymphocyte subsets in spleen and the number of the NK (also known as CD3−CD49+) cells. After killing the mice, the spleen was carefully removed and prepared into splenocyte suspension. The erythrocytes were lysed by addition of 0.83% Tris-NH4Cl solution. After three rinses with PBS (pH 7.4), the splenocytes were resuspended with staining buffer to obtain 1×106/mL density. Splenocytes (100 μL aliquots) were dispensed into four tubes and each was surface labeled with 5 μL of CD8-FITC, CD4-PE-Cy5, CD3e-PE Hamste, and CD49b-FITC, respectively. All mixtures were incubated at 4°C for 30 minutes in darkness followed by addition of 200 μL precooled staining buffer. After centrifugation at 1500 rpm for 5–10 minutes, the precipitates were resuspended twice and fixed in 200 μL 1% paraformaldehyde. Labeled cells were washed and analyzed with an FACSCalibur flow cytometer using the CellQuest and WinMDI 2.9 software. In each case, the staining was compared with that of the appropriately labeled isotype control antibody.
Statistical analysis
All data were analyzed by SAS 6.12 software and the results were expressed by mean±SD. To compare the differences between multiple groups, statistical significance was analyzed using a one-way analysis of variance followed by post hoc comparisons. Statistical significance was defined as p-values<0.05.
Results
Tumor growth curve and inhibition rates
Tumor volumes continued to increase in the PBS and Lacz groups throughout the study period and significant differences in the volume in both groups were observed on days 14 and 21 compared with the volumes on days 0 and 7 (p<0.01). Although slight increase in the tumor volumes was also present in Ad-mIFN-λ2 group, the volumes were significantly smaller than those observed in the PBS and Lacz groups at days 14 and 21 (p<0.01). The tumor growth curve is displayed in Figure 1. The tumor inhibition rates were calculated based on the last measurement of tumor volume and according to the formula described in the Materials and Methods section.
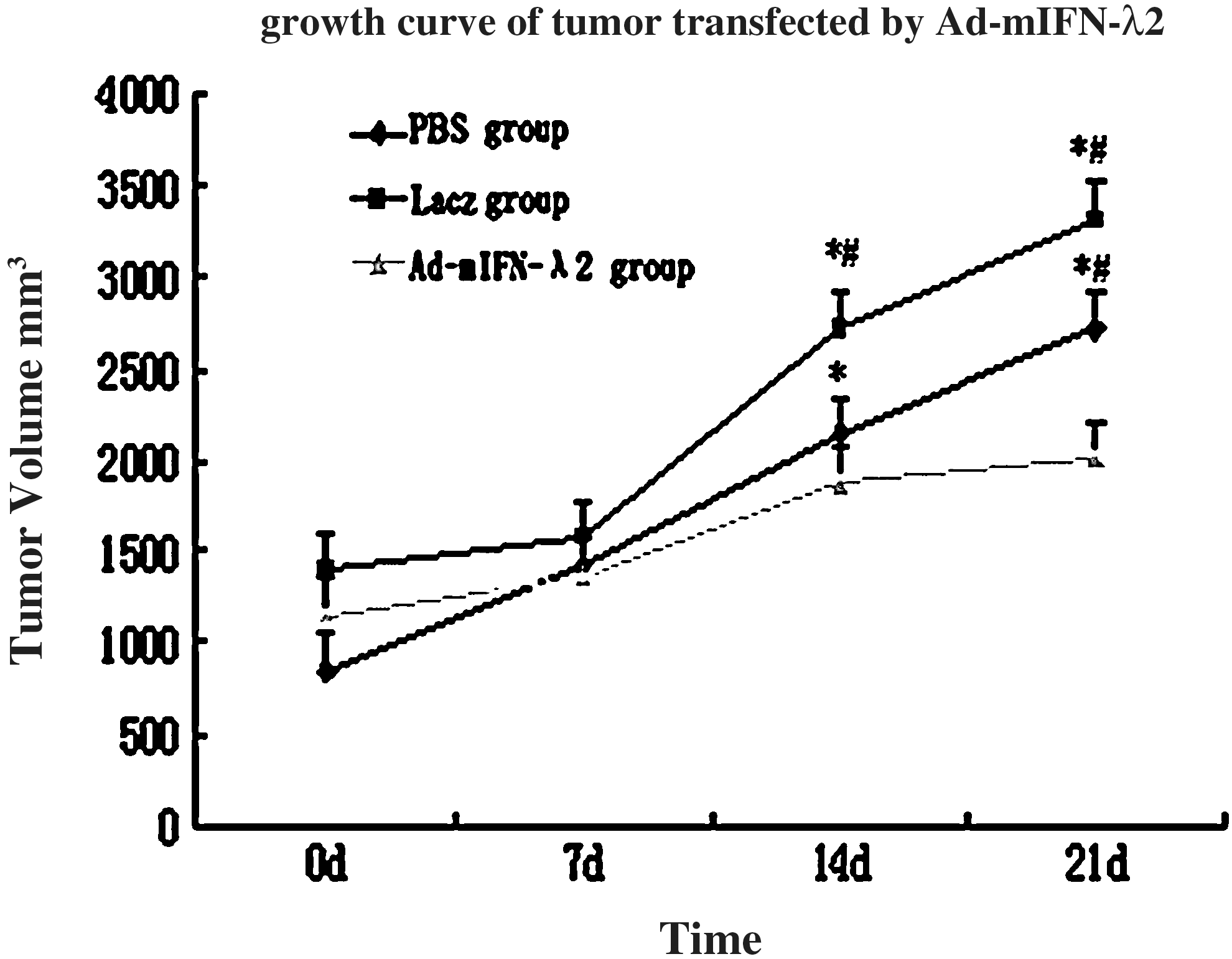
Growth curve of tumor. The symbol * indicates significant increase (p<0.01) in tumor volume in the phosphate-buffered saline (PBS) and Lacz groups on days 14 and 21 compared with days 0 and 7. The symbol # indicates significant decrease (p<0.01) in tumor volume in Ad-mIFN-λ2 group at days 14 and 21 compared with PBS and Lacz groups. Tumor inhibition rate=(2724.8–1411.15)/2724.8×100%=48.2%. Which means that the tumor growth in the Ad-mIFN-λ2 group in comparison with the other two groups was inhibited by 48.2%.
Tumor inhibition rate=(2724.8 – 1411.15)/2724.8×100%=48.2%. Which means that the tumor growth in the Ad-mIFN-λ2 group in comparison with the other two groups was inhibited by 48.2% (see Fig. 1).
Morphological and histopathological analysis
Cystic degenerations were seen in the gross specimens of the tumors from the Ad-mIFN-λ2 group. However, solid tumor degeneration was also present in the control group.
Subcutaneous tumor necrosis was more evident in the tissue sections from the Ad-mIFN-λ2 group compared with the other two groups.
The spleen tissues were significantly increased, some even exhibiting twofold increase, in the Ad-mIFN-λ2 group of mice compared with those in the other two groups. In addition, Ad-mIFN-λ2 transfection induced more multinucleated giant cells in the spleen and widening of the white pulp.
In the lung tissue, severe inflammatory reaction occurred after Ad-mIFN-λ2 transfection, while only moderate inflammation was observed in the Lacz group and no inflammation in the PBS group (see Fig. 2).
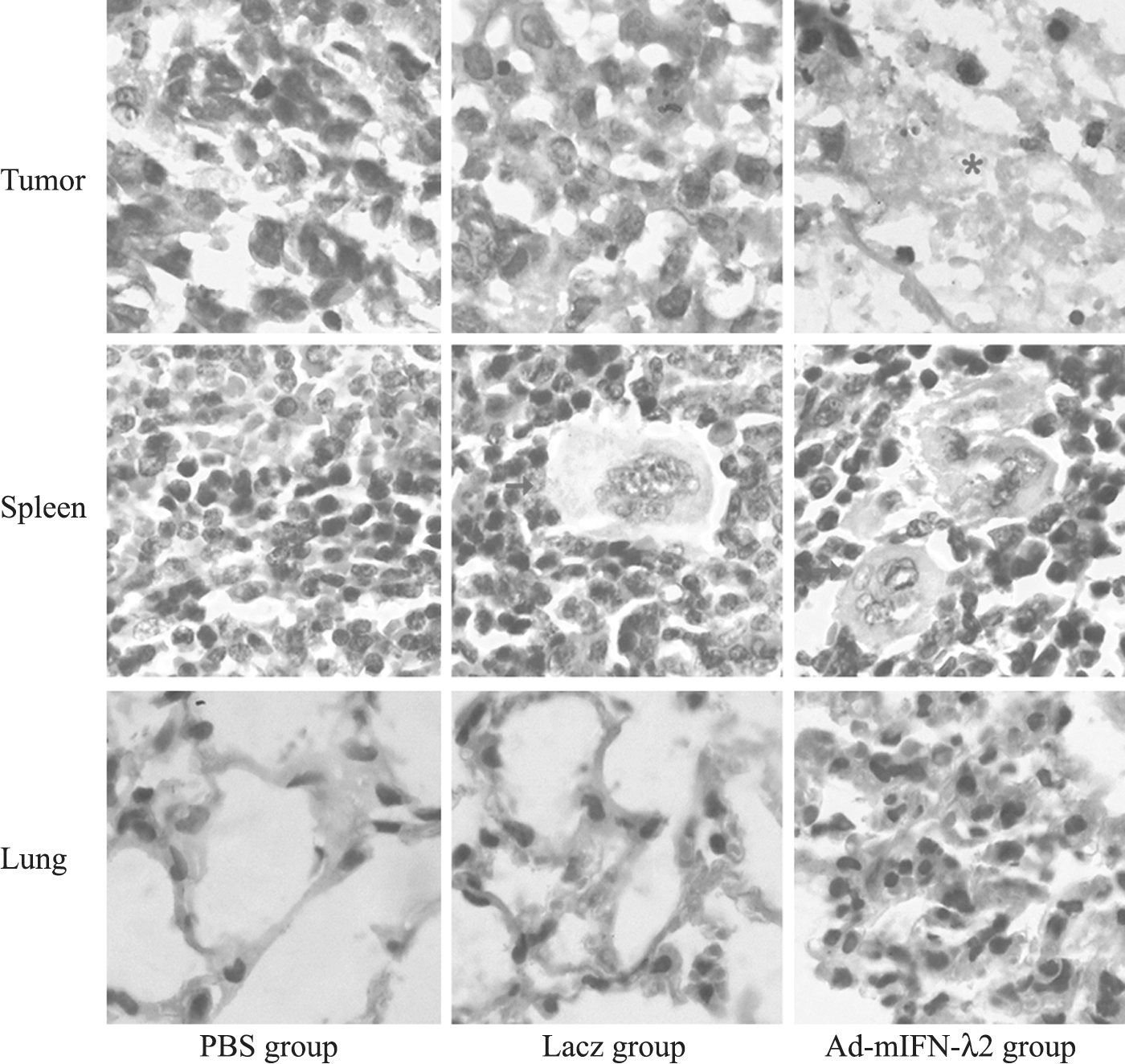
Tissue sections (hematoxylin and eosin staining) from tumor, spleen, and lung showing histopathological changes following transfections. Asterisk indicates subcutaneous tumor necrosis in the tumor. Arrows indicate multinucleated giant cells in the spleen. Much more subcutaneous tumor necrosis in the tumor tissue and multinucleated giant cells in the spleen of the mice transfected by Ad-mIFN-λ2.
Apoptosis of tumor cells
TUNEL method was used to determinate apoptosis in the tumor cells. As expected, apoptotic cells and apoptotic index were significantly higher in Ad-mIFN-λ2 group than in the PBS and Lacz groups (p<0.05). There was no significant difference between the PBS and Lacz groups (p>0.05) (see Fig. 3 and Table 2).
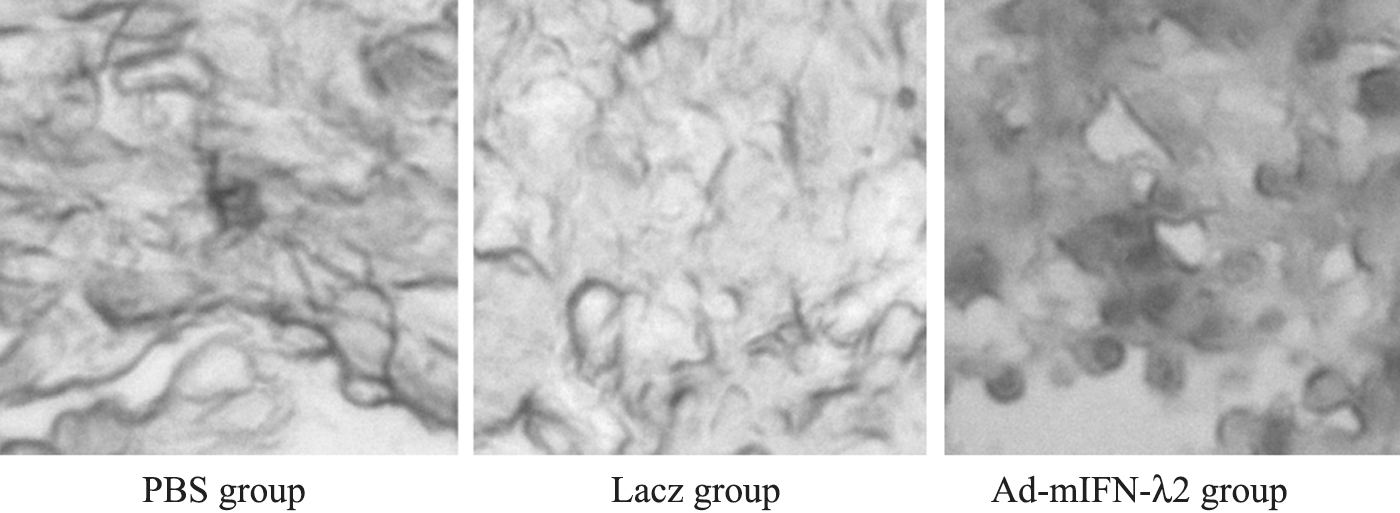
Analysis of apoptosis in tumor cells after the three different transfections. Arrow indicates apoptosis of subcutaneous tumor cells.
mIFN-λ2 versus the other two groups (p<0.05).
Lacz versus PBS (p>0.05).
AI, apoptotic index; PBS, phosphate-buffered saline.
Expression of mIFN-λ2 protein
Western blotting was performed to determine mIFN-λ2 protein expression. Results indicated observable mIFN-λ2 expression in subcutaneous tumor tissue after transfection with Ad-mIFN-λ2. In the PBS and Lacz groups, mIFN-λ2 was rarely expressed (see Fig. 4).
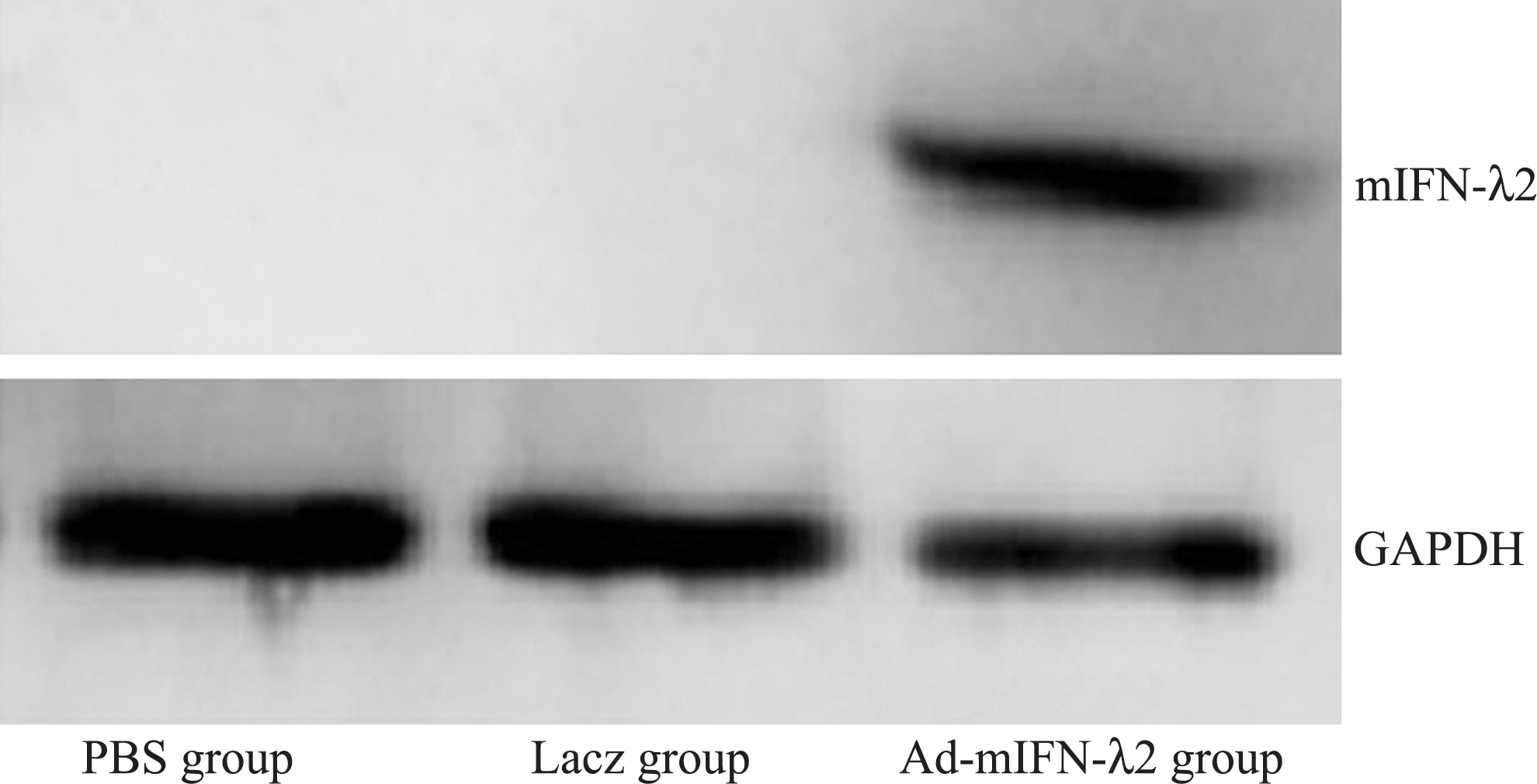
Expression of mIFN-λ2 protein in subcutaneous tumor tissue following transfection. 1: PBS group; 2: Ad-Lacz group; 3: Ad-mIFN-λ2 group. Observable mIFN-λ2 expression in subcutaneous tumor tissue after transfection with Ad-mIFN-λ2.
Expression of mIFN-λ2 gene
RT-PCR was performed to assess mIFN-λ2 gene expression in the fractional skeletal muscle and the tumor tissue. The analysis showed evident mIFN-λ2 gene expression (∼220 bp) after transfection with Ad-mIFN-λ2 in both tissues. In PBS and Lacz groups, mIFN-λ2 gene was rarely expressed (see Fig. 5).
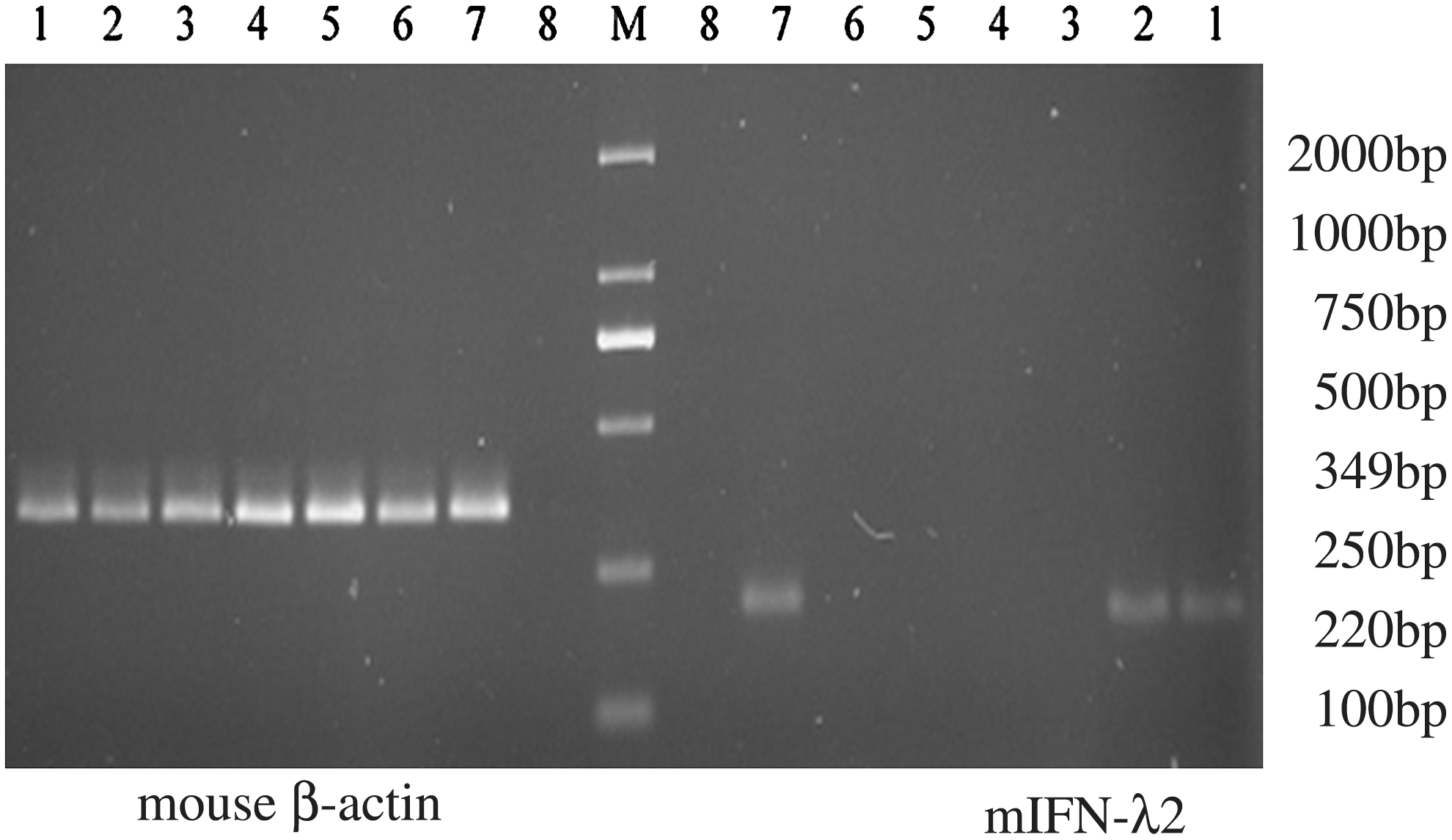
Expression of mIFN-λ2 mRNA in the tissues of the tumor and skeletal muscle following transfection. 1: Ad-mIFN-λ2 group, skeletal muscle tissue; 2: Ad-mIFN-λ2 group, tumor tissue; 3: Ad-Lacz group, skeletal muscle tissue; 4: Ad-Lacz group, tumor tissue; 5: PBS group, skeletal muscle tissue; 6: PBS group, tumor tissue; 7: positive control group; 8: NTC: negative control; M: Marker. The analysis showed evident mIFN-λ2 gene expression (∼220 bp) after transfection with Ad-mIFN-λ2 in both skeletal muscle tissues and tumor tissue.
T lymphocyte subset in spleen and number of NK cells
Flow cytometry was performed to detect the number of CD3+CD4+, CD3+CD8+, and NK (CD3−CD49+) cells. The results showed the number of CD3+CD4+, CD3+CD8+, and NK cells was significantly higher in Ad-mIFN-λ2 group than in the PBS and Lacz groups (p<0.001). No significant differences were found between the PBS and Lacz groups (see Fig. 6 and Table 3).
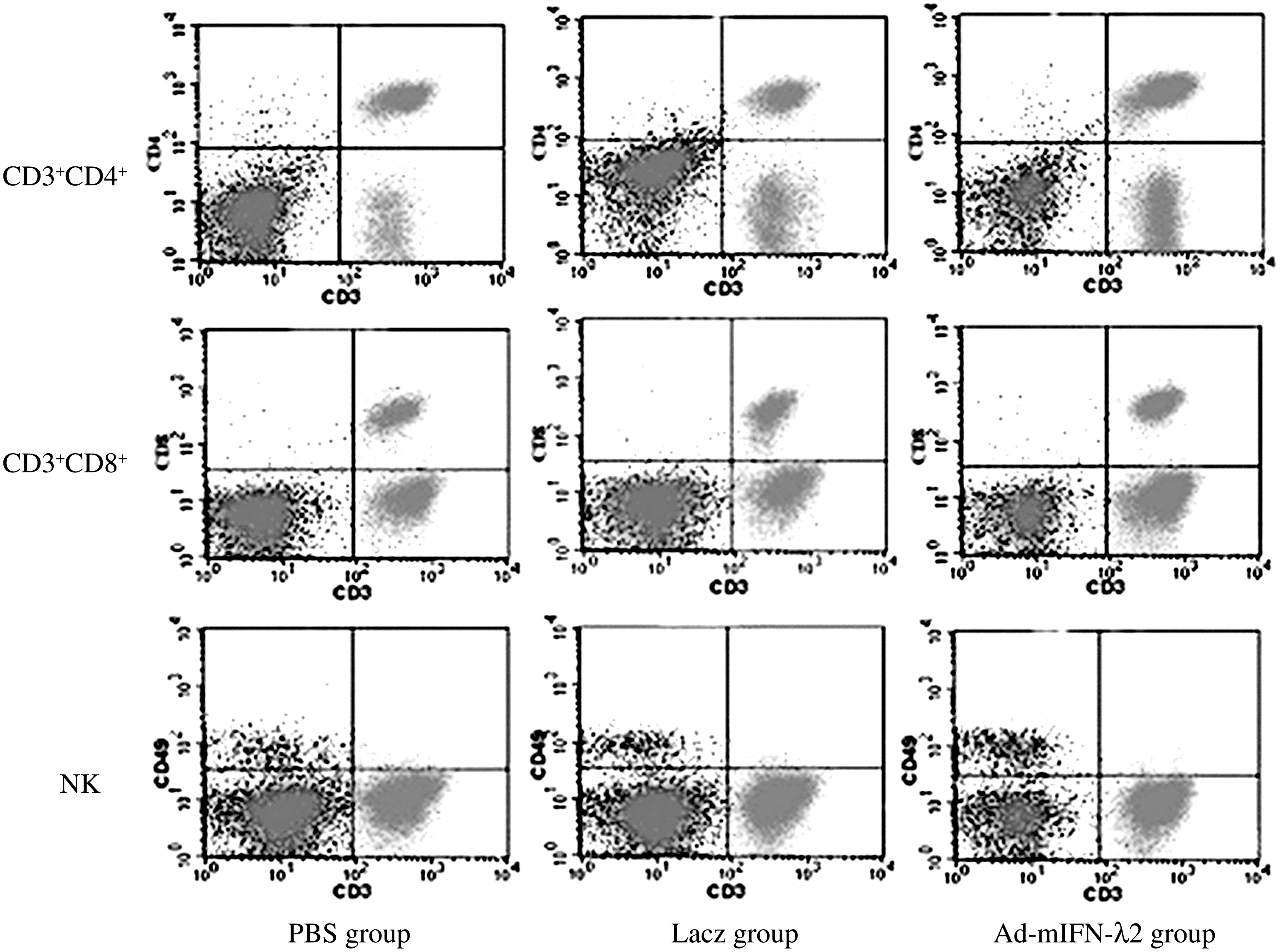
Flow cytometry for detection of spleen T lymphocyte subtype and the number of CD3+CD4+, CD3+CD8+, and NK cells.
Lacz group versus PBS group (p>0.05).
Ad-mIFN-λ2 group versus the other two groups (p<0.01).
Ad-mIFN-λ2 group versus the other two groups (p<0.001).
Discussion and Conclusions
In general, gene therapy is defined as a method of transfer of genes or gene-fusion vectors to somatic cells to induce production of specific therapeutic proteins in order to correct or modulate the disease. To date, adenoviral vectors are the most widely used vector systems for cancer gene therapy due to their advantageous characteristics. 10 Adenoviruses can infect a variety of target cell types with the exception of hematopoietic cells. In human, most tumors are derived from epithelial cells thus rendering the adenoviral vectors, epithelia-specific virus vectors, suitable for either treating tumors directly or for mediating exogenous protein expression of the epithelial cells.
mIFN-λ2 is a new member of the type III IFN family that possesses similar signal pathway and biological activity as type I IFN. 4,5 mIFN-λ2 was induced by Newcastle disease virus infection of mouse embryonic fibroblasts, 11 herpes simplex virus type 2 (HSV-2), EMCV, Sendai virus, and influenza A virus in different types of cells, including A549, Hela, Raji, and U937 cells. 12 The receptor complex for IFN-λ2 includes two subunits IL-28Ra and IL-10Rb. The subunits functionally form a homodimer that mediates the intracellular signaling and activation of biological activities of IFN-λ2. IL-28Ra is more IFN-λ2 specific, whereas IL-10Rb is an accessory chain that is also part of the receptors for the IL-10 family of cytokines, such as IL-10, IL-22, and IL-26. 5 IFN-λ2 induces intracellular signals similar to type I IFNs that promote cell apoptosis and inhibit cell proliferation by activating caspase-3, caspase-8, and caspase-9. 6,7
IL-28Ra-induced signaling might represent a promising therapeutic target to protect the intestinal epithelium from viral infections. Jieliang Li et al. have indicated that the TLR3/IFN-k pathway may play a crucial role in the innate protection against HSV-1 infection of the central nervous system (CNS). 13 In addition, IFN-λs proved effective in treatment of hepatitis C virus and with less side effects than IFN-α. 14,15
Several reports have indicated that IFN-λs play an important role in tumor inhibition through the following mechanisms: inhibition of tumor virus replication; induction of cell apoptosis; suppression of cell proliferation; activation of cell immune response that helps eliminating tumor cells; and regulation of tumor–stromal cell interactions and antineovascularization.
Brand et al. showed recently that IFN-λ2 markedly suppressed the growth of human neuro-endocrine BON1 tumor cells possibly through active cell immune response, including activating polymorphonuclear leucocyte, CD3+ CD8+ cell, and NK cell, and enhancing secretion of other cytokines. 16 However, this conclusion is still controversial. 8,9
There is emerging evidence that IFN-λs significantly inhibit the growth of tumor cells, such as BW5147, BON1, and LN319, in vitro. IFN-λs also significantly inhibit the growth and metastasis of tumor cells in mice melanoma, colorectal cancer, and fibrosarcoma in vivo. 8,9,17 In China, research on IFN-λs is limited and only a few researchers devote themselves to constructing IFN-λ1/IFN-λ2 vectors and carry out transfection studies. However, Tian et al. showed that there was a negative correlation between the expression level of IL-29 and the depth of inflammatory infiltration in esophageal cancer. 18
However, there have been no reports on the antitumor effects of IFN-λs on lung cancer. And, therefore, this study was set up to investigate the inhibitory effect of mIFN-λ2 genes on LA795 lung adenocarcinoma cells in tumor-bearing mice. The analysis showed that the tumor volumes were significantly reduced after Ad-mIFN-λ2 transfection and in some cases the tumors were accompanied by cystic degeneration. The tumor necrosis increased, accompanied by enlarged spleen, multinucleated giant cell formations in the spleen, and wider white pulp. Both the western blotting and RT-PCR suggested that Ad-mIFN-λ2 vector successfully infected the LA795 adenocarcinoma cells in the tumor-bearing mice as seen by the increase in mIFN-λ2 protein and gene expression in Ad-mIFN-λ2-transfected group. TUNEL assay indicated that apoptotic cells and apoptotic index were significantly higher in Ad-mIFN-λ2 group than in the PBS and Lacz groups. Flow cytometry indicated that the number of CD3+CD4+, CD3+CD8+, and NK cells was significantly higher in Ad-mIFN-λ2 group than in the control group.
In conclusion, the results of this study suggest that Ad-mIFN-λ2 inhibits the growth of lung cancer cells and promotes apoptosis to a certain extent. It induces CD3+CD4+, CD3+CD8+, and NK cells thus strengthening the cell immune response and ultimately leading to an antitumor effect. On the other hand, the activation of the innate immunity may also have effects on inflammatory cytokines, such as TNF and IL-6. 19 It will be necessary to study which stage of the tumor cells apoptosis. We anticipate that our study lays an experimental basis for further clinical application of Ad-mIFN-λ2 in lung cancer therapy.
Footnotes
Acknowledgment
This work was supported by the foundation of Ministry of Health of China (No. wkj2006-020206).
Disclosure Statement
No competing financial interests exist.
Conflict of Interest
The authors certify that regarding this article, no actual or potential conflicts of interests exist. The work is original, it has not been accepted for publication nor is concurrently under consideration elsewhere, and it will not be published elsewhere without the permission of the Editor and that all the authors have contributed directly to the planning, execution, or analysis of the work reported or to the writing of the article.