
Abstract
Ferulic acid (FA) is a monophenolic phenylpropanoid occurring in plant products such as rice bran, green tea, and coffee beans. It has been shown to have significant antioxidant effects in many studies. In the present study, we show that intraperitoneal administration of FA at a dose of 50 mg/kg body weight 1 hour prior to or immediately after whole-body γ-irradiation of mice with 4 Gy results in considerable reduction in the micronuclei formation in peripheral blood reticulocytes. Administration of the same amount of FA immediately after 4 Gy γ-irradiation showed significant decrease in the amount of DNA strand breaks in murine peripheral blood leukocytes and bone marrow cells as examined by comet assay. Further, immunostaining of mouse splenic lymphocytes for phspho-γH2AX was carried out, and it was observed that FA inhibits the γH2AX foci formation. Finally, the survival of mice upon 6, 8, and 10 Gy γ-ray exposure was monitored. FA enhances the survival of mice by a factor of 2.5 at a dose of 6 Gy γ-radiation but not at higher doses. In conclusion, FA has protective potential in both pre- and postirradiation exposure scenarios and enhances the survival of mice possibly by decreasing DNA damage as examined by γH2AX foci, micronuclei formation, and comet assay.
Introduction
Ionizing radiations interact with cells by inducing a wide range of damaging effects, with the most vital alterations occurring in the cellular DNA. The generation of reactive oxygen species (ROS) through radiolysis of water brings about oxidation of critical cellular machinery leading to DNA strand breaks, peroxidation of lipids in cellular membranes, and protein oxidation. 1,2 Ionizing radiation is a well-known mutagen and the DNA of the exposed cells undergoes single-strand breaks (SSBs), double-strand breaks (DSBs), and damage to the bases and sugars, DNA-protein cross-links, ultimately leading to chromosomal aberrations. Depending on the severity of the damage, the cell may incorporate the damage (mutagenesis) and proliferate, leading to carcinogenesis or may die and get eliminated. Therefore, prevention of cytogenetic damage in the exposed cell may lead to a reduction in chromosomal instability, mutagenesis, and carcinogenesis.
Radiation has been considered an enigma to the general public and the use of radiation for therapeutic and other uses has always been associated with skepticism. With the advent of the nuclear age there came the realization that people would require protection against the toxic effects of radiation that could occur from nuclear accidents, medical exposures, and nuclear war, with the latter risk being the primary driving force for government research and development of radioprotectors during the Cold War. 3 Many natural and synthetic chemicals have been investigated in the recent past for their efficacy to protect against radiation-induced damages in biological systems. 4 –9 However, due to inherent toxicity of some of the synthetic agents, the practical applicability of the majority of these synthetic compounds remained limited at the radioprotective concentration. Therefore, this need has warranted further search for nontoxic and effective alternatives to the synthetic compounds. In fact, no radioprotective agents are now available, either alone or in combination to meet all the requisites of an ideal radioprotector. 10 Amifostine is the only one that is currently in use having good radioprotection to the normal tissues during radiotherapy, even though there are reports about contraindications in some cases. 11 Radiation-induced biological damage mainly arises from the interaction of the radiation-induced free radicals or ROS with biomolecules. Agents that can prevent the formation of free radicals or destroy free radicals by reacting with them, thereby inhibiting their reaction with biomolecules might be considered as radioprotectors. Since free radicals are short-lived, it is necessary for such radioprotective molecules to be present in the cellular milieu in sufficient concentration at the time of radiation exposure.
Ferulic acid (FA; Scheme 1) is a monophenolic phenylpropanoid occurring in plant products such as rice bran, green tea, and coffee beans. It has the ability to act as an antioxidant against peroxyl radical-induced oxidation in neuronal culture and synaptosomal membranes. 12 It scavenges the ROS such as hydroxyl radical (·OH), hypochlorous acid (HOCl), and peroxyl radical (RO2·) 13 and stable free radical 1, 1-diphenyl-2-picrylhydrazyl (DPPH). 14 It significantly reduces the NO·production by lipopolysaccharide-stimulated mouse macrophage-like cells (RAW 264.7 cells) and is useful for preventing cell damage caused by superoxide (O2·−), OH, and NO·in living systems. 15 It has an inhibitory effect on 4-nitroquinoline 1-oxide (4-QO)-induced rat tongue carcinogenesis 16 and depresses 12-O-tetradecanoyl-phorbol-13-acetate (TPA)-promotion of skin tumorogenesis. 17 No in vivo data are available, but in vitro studies on cytotoxicity indicate that in rat hepatocytes the LD50 is 25 mM. 18 Zhao and Moghadasian (1998) has estimated that the sum of FA intake through consumption of cereals, vegetables, fruits, coffee, and juices may reach 150–250 mg/day. 19 Pharmacokinetics of FA in mouse serum is having t1/2 (ka)=1.249±0.365 hours. 20 Short-term feeding study showed that FA-sulfoglucuronide is the main metabolite in the plasma of rat administered FA or its sugar esters. 21,22

Structure of ferulic acid.
Our previous study showed that FA has good radioprotection to plasmid DNA under in vitro conditions and peripheral blood leukocytes and bone marrow cells under in vivo conditions when drug was administered 1 hour prior to 4 Gy γ-radiation exposure. 23 It enhances the DNA repair process in the peripheral blood leucocytes of mice in vivo. 23 It also shows preferential radioprotection to normal tissues compared with tumor cells both under ex vivo and in vivo situations of radiation exposure. 24 Prasad et al. 25 have explored the radioprotective effect of FA on γ-radiation-induced dicentric aberrations and lipid peroxidation with reference to alterations in cellular antioxidant status in cultured lymphocytes. Recently, Ma et al. 26 have suggested that the radioprotective efficacy by FA may be a result of early recovery of hematopoietic cells due to enhanced production of granulocyte colony-stimulating factor and erythropoietin. They have also reported that FA had a radioprotective effect mediated through the extracellular signal-regulated kinases pathway to inhibit apoptosis and oxidation. 27 The present study is concerned with the effect of FA on γ-radiation-induced DNA strand breaks as monitored by micronuclei assay, γH2AX foci, and comet assay. Finally, to confirm the efficacy of radioprotection in real-life situations, the survival of mice was also monitored at lethal doses.
Materials and Methods
Materials
FA, high melting point agarose, low melting point agarose, acridine orange (AO), Na2-EDTA, triton X-100, dimethyl sulfoxide, tris-base, and propidium iodide (PI) were obtained from Sigma-Aldrich Chemicals, Inc. All other chemicals used were of analytical grade procured locally. Antibodies such as phospho-γH2AX and Alexa fluor 488 conjugates were purchased from Cell Signaling Technology).
Animals
Male Swiss mice, 8–10 weeks old and weighing 20–25 g, were selected from an inbred group maintained under standard conditions of temperature (25°C±2°C) and humidity. Animals were provided with food and water ad libitum. Usually, four animals were housed in each sterile polypropylene cage containing sterile paddy husk as bedding. The guidelines issued by the Institutional Animal Ethics Committee of Bhabha Atomic Research Centre, Government of India, regarding the maintenance and dissections of small animals were strictly followed.
Drug administration
For all in vitro experiments 10 mM FA stock was made in 75 mM potassium phosphate buffer (PPB), pH=7.4. Animals were administered FA (50 mg/kg body weight) dissolved in 75 mM PPB and each mouse received 200 μL of FA per injection intraperitoneally.
Gamma-irradiation
A 60Co-Junior Theratron Teletherapy unit (AECL; dose rate 0.39 Gy/min at 38 cm distance) was used for micronuclei and DNA damage study and Gamma Cell 220 for survival study (dose rate 5 Gy/min).
Micronuclei assay in mouse blood reticulocytes
For micronuclei assay, animals were divided into the following groups; (1) PPB+sham-irradiation; (2) PPB+4 Gy irradiation; (3) FA (50 mg/kg body weight)+sham-irradiation, and (4) FA (50 mg/kg body weight)+4 Gy irradiation.
Experiments were done in two sets that is, in the first set animals were administered PPB or FA 1 hour prior to 0 Gy (sham-irradiation) or 4 Gy whole-body γ-irradiation, whereas in the other set PPB or FA was administered immediately after completion of irradiation. For micronuclei study, first AO-coated glass slides were made according to Hayashi et al. 28 In brief, AO was dissolved in distilled water at a concentration of 1 mg/mL. Ten microlitres of this solution was placed on a preheated (about 70°C) cleaned glass slide. The solution was spread by moving a glass rod back and forth over it and air-dried. The AO-coated glass slides were stored at room temperature under dry conditions.
To study micronuclei in peripheral blood of mice, first blood was collected by tail vein puncture at 24 and 48 hours time points and 5 μL aliquots of the blood were kept on AO-coated slides and covered with coverslips. The reticulocytes (RETs) and micronucleated reticulocytes (MN-RETs) were monitored using a Carl Zeiss Fluorescent microscope equipped with Axioskop and blue filter. A suitable region of the slides was selected under low magnification and ∼1000 RETs were scored for micronuclei.
Effect of FA on radiation-induced DNA strand breaks in murine tissues
The animals were divided in the same way as in case of micronuclei assay. Immediately after whole-body exposure to 0 Gy (sham-irradiation) or 4 Gy γ-radiation animals were administered PPB or FA (50 mg/kg body weight) and kept in polycarbonated cages. Animals were sacrificed by cervical dislocation 1 hour postirradiation; blood was withdrawn from the heart using a heparinized hypodermic syringe and collected in eppendorf tubes while bone marrow cells were collected by flushing the femur bone of each animal with PBS. All the samples were stored on ice. For studying the DNA damage in single cell suspensions of blood and bone marrow, alkaline single cell gel electrophoresis (comet assay) was carried out. 23,24,29 For capturing the image of comet, the slides were stained with PI by layering 50 μL of PI (stock, 20 μg/mL) on the top and visualized using a Carl Zeiss Axioskop microscope with bright field phase-contrast and epifluorescence facility. The images (50–60 cells/slide) were captured with high-performance JVG TK 1280E color video camera. The integral frame grabber used in this system (Cvfbo1p) is a PC based card and it accepts color composite video output of the camera. The quantification of the DNA strand breaks of the stored images was done using the CASP software by which tail length and Olive tail moment could be obtained directly. 30
Effect of FA on γH2AX foci formation in splenic lymphocytes
Splenic lymphocytes were obtained by gently squeezing the Swiss mouse spleen through a nylon mesh in a petriplate containing RPMI-1640 medium supplemented with 100 units/mL penicillin and 100 μg/mL streptomycin. The RBCs were lysed by brief hypotonic shock. Lymphocytes were suspended in RPMI 1640-medium. Cells were incubated with FA for 30 minutes before 2 Gy γ-radiation exposure. Fifteen minutes postirradiation, cells were kept on ice and attached on the coverslip coated with poly-
Survival study
To study the survival of mice against different doses of γ-radiation animals were divided into the following groups; (1) PPB control; (2) FA control; (3) 6 Gy+PPB; (4) 6 Gy+FA; (5) 8 Gy+PPB; (6) 8 Gy+FA; (7) 10 Gy+PPB, and (8) 10 Gy+FA.
Immediately after exposure to different doses of γ-radiation that is, 0 (sham-irradiation), 6, 8, and 10 Gy animals were administered a single dose of PPB or FA (50 mg/kg body weight) for three consecutive days. After that animals were kept under observation for 31 days and their death was monitored.
Statistical analysis
The graphs were plotted using Mean±standard error of mean). One-way analysis of variance with Tukey–Kramer Multiple Comparisons as post-test for p<0.05 was used to study the significance level. For mouse survival analysis, Kaplan–Meier's estimate of life time analysis was done using GraphPad Prism 5 software (GraphPad software, Inc.). A dead mouse was counted as censored 0 whereas a live mouse was defined as censored 1. The difference between the estimated survival times of the two groups was evaluated by the log-rank test. The statistical significance of the log-rank test was considered if p<0.05 according to chi-square distribution.
Results
FA inhibits the formation of micronuclei in mouse peripheral blood RETs
It can be seen in Table 1 that when animals were exposed to 4 Gy γ-radiation there was increase in the micronuclei in blood RETs at 24 and 48 hours, but the administration of FA (50 mg/kg body weight) significantly (p<0.001) reduced the number of micronuclei formation. Preirradiation administration of FA is more effective compared with that at postirradiation.
p<0.001.
Animals were administered FA 1 hour prior to radiation exposure (Table A) or immediately after (Table B). Formation of micronuclei was monitored at 24 or 48 hours postradiation exposure.
FA, ferulic acid; MN-RETs, micronucleated reticulocytes; PPB, potassium phosphate buffer; SEM, standard error of mean.
FA inhibits the formation of γH2AX foci in mouse splenic lymphocytes
Our previous experiments indicated that FA has potential to inhibit the formation of micronuclei in blood RETs. It is also well known that the micronuclei formation takes place by double-strand breaks. Since γH2AX has been shown to be a sensitive indicator of DSB, the effect of FA on γH2AX foci formation in splenic lymphocytes was monitored by confocal microscopy and flow cytometry, and it was found that FA has a concentration-dependent inhibition of γH2AX foci formation (Fig. 1A, B).
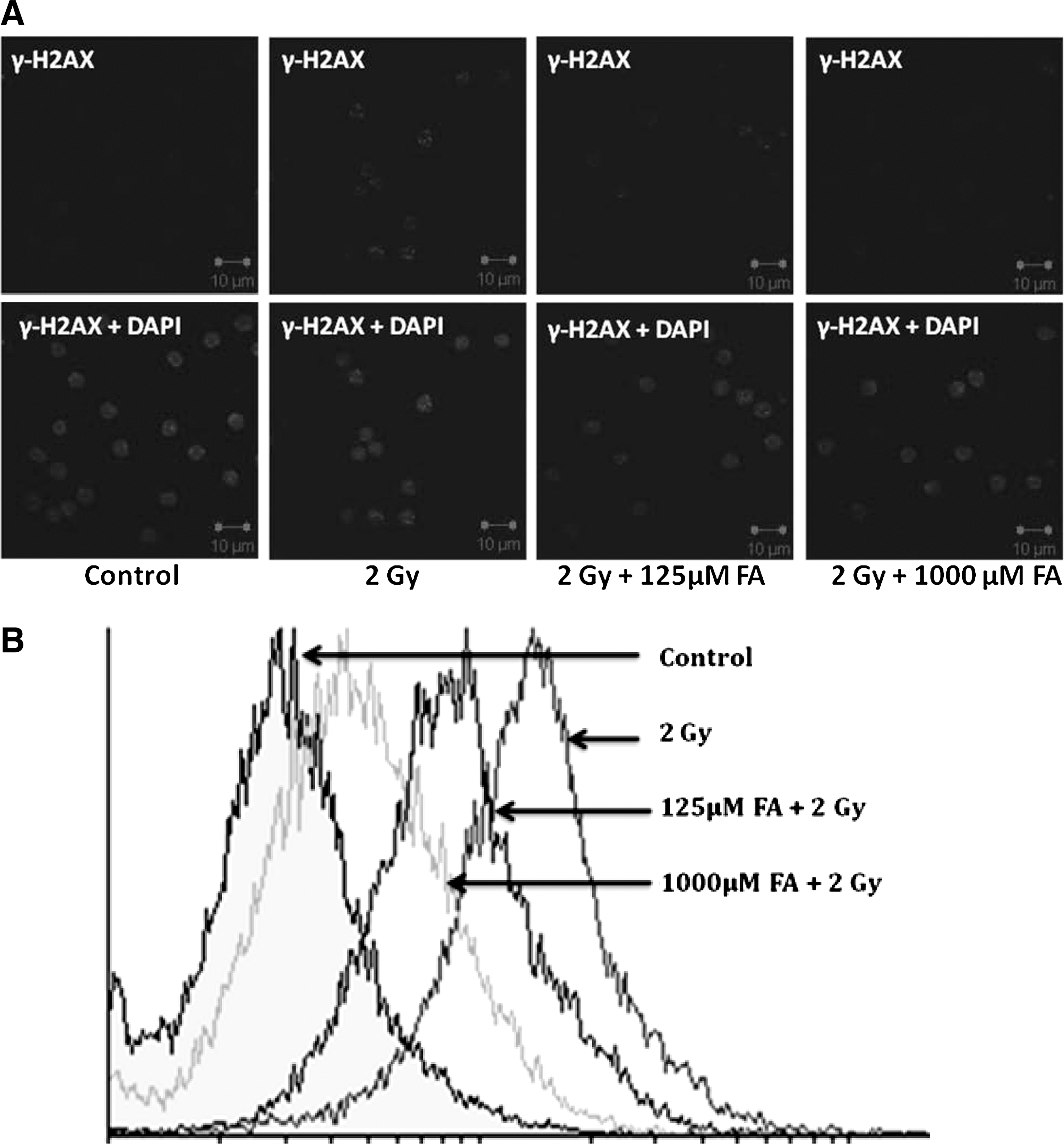
Effect of ferulic acid (FA) on γH2AX foci formation after 2 Gy γ-radiation exposure in splenic lymphocytes. Cells were incubated with 125 and 1000 μM FA and later exposed to 2 Gy γ-radiation. Cells were processed for immunostaining as described in the Materials and Methods section.
FA reduces γ-radiation-induced DNA damage in murine blood leukocytes and bone marrow cells
Damage to cellular DNA in blood leukocytes and bone marrow cells upon whole-body exposure of mice to γ-radiation was studied after 1 hour postirradiation. Whole-body exposure of mice to 4 Gy γ-radiation resulted in increase in comet parameters, as can be evidenced in Figures 2 and 3. Administration of FA (50 mg/kg body weight) i.p. immediately after γ-radiation exposure resulted in significant reduction of the comet parameters (Figs. 2 and 3).
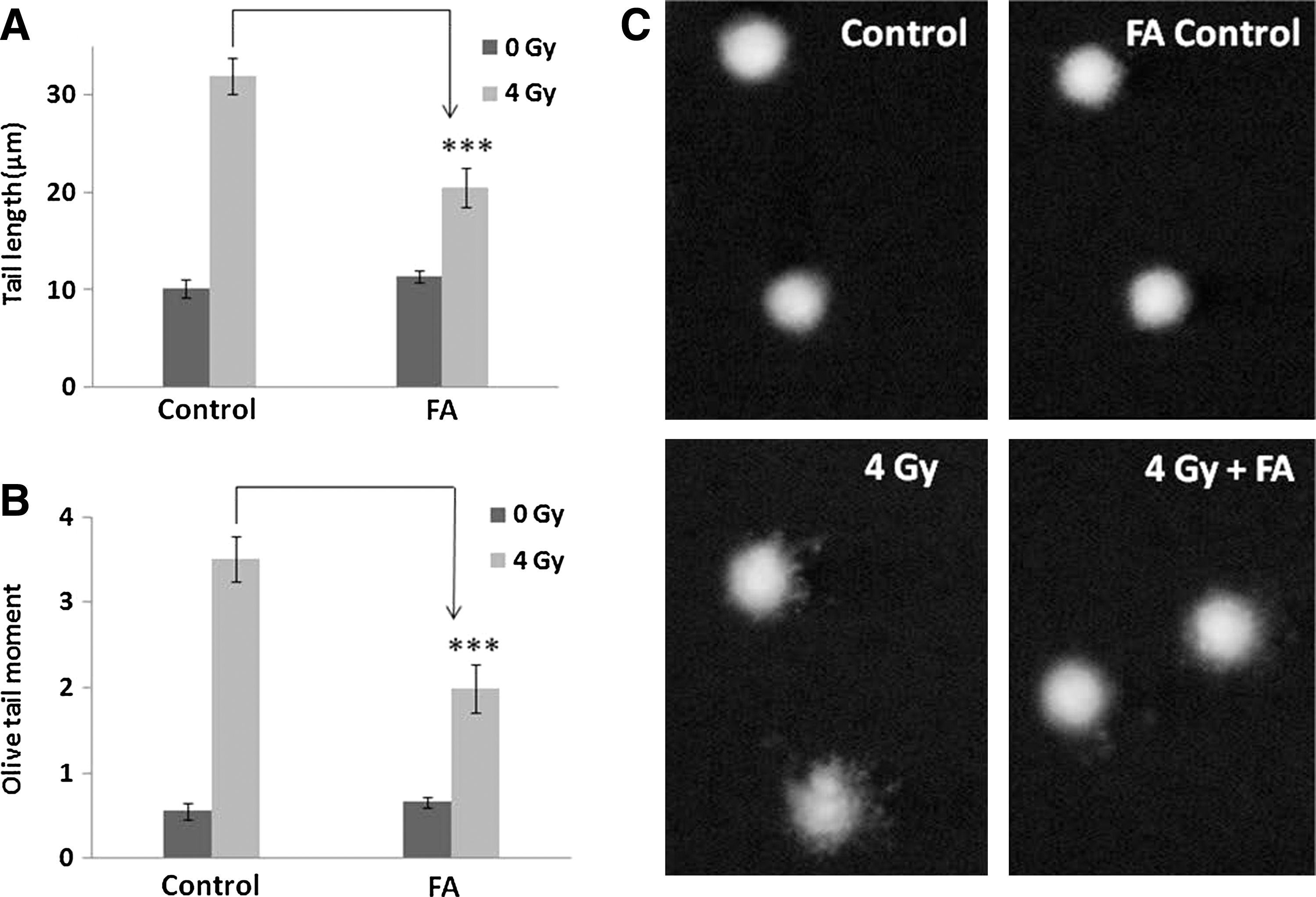
Effect of FA on DNA damage in murine blood leucocytes as assayed by comet assay. Blood leukocytes collected at the end of experiment were subjected to alkaline single cell gel electrophoresis as described in the Materials and Methods section. Tail length
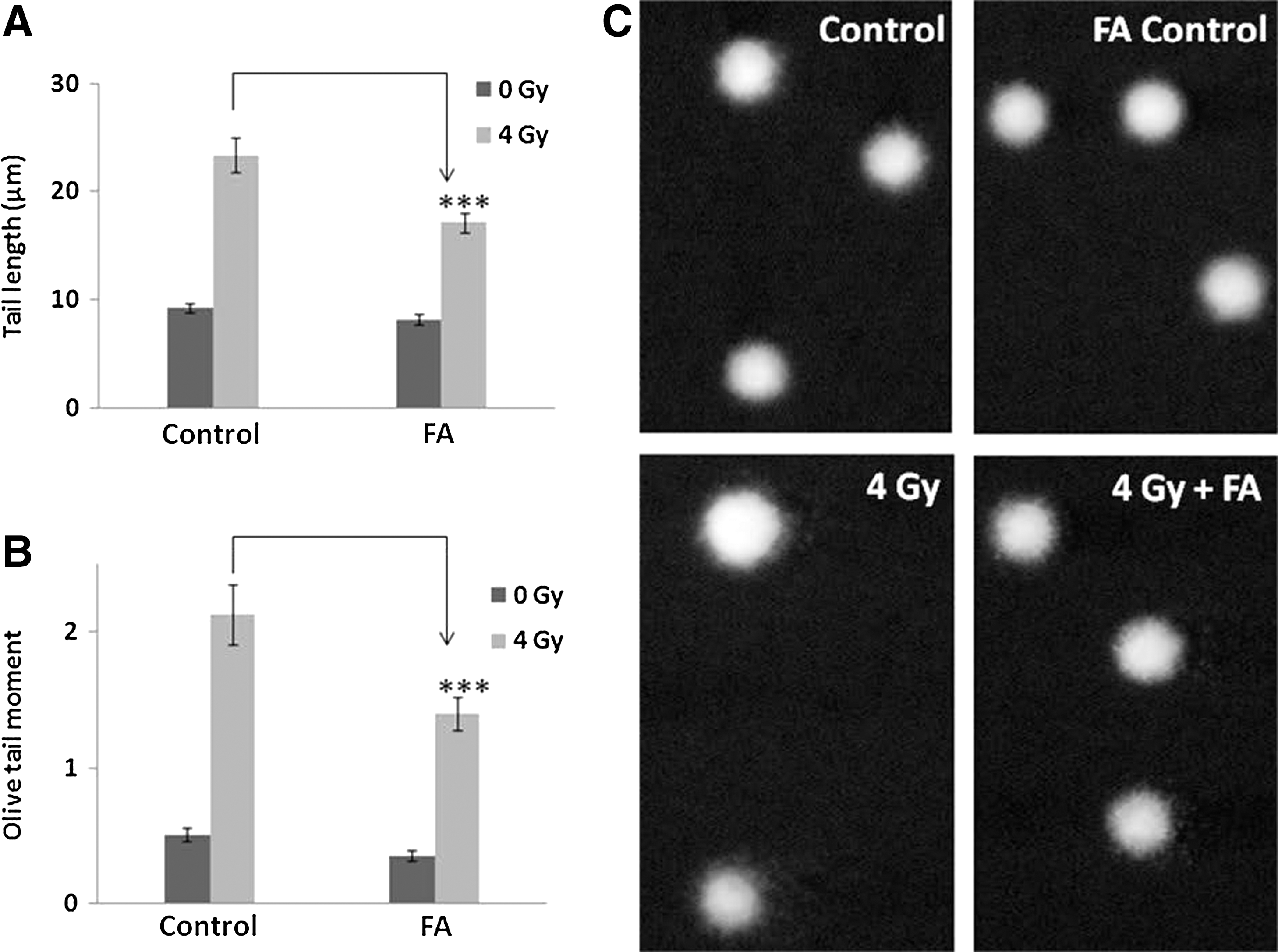
Effect of FA on DNA damage in murine bone marrow cells as assayed by comet assay. Single cell suspensions of bone marrow were collected at the end of experiment and subjected to alkaline single cell gel electrophoresis as described in the Materials and Methods section. Tail length
FA enhances the survival of Swiss mice
As shown in Figure 4, in survival study, at every radiation dose where FA was administered there was enhancement in the survival of mice. There was no adverse effect in the FA control group compared with PPB control group. To represent our data in relative terms, we calculated Survival Enhancement Ratio (SER) using the following formula;
Our finding showed that FA (50 mg/kg body weight) has an SER 2.5 at a radiation dose of 6 Gy.
Discussion
The exposure of human cells or tissues to ionizing radiation causes genomic instability leading to mutagenesis and carcinogenesis; therefore, there is a need to protect the genome against such effects of ionizing radiation. Ionizing radiation-induced damage to cellular DNA is mainly due to strand breaks and may lead to loss of viability of cells. Protecting cellular DNA from radiation damage might result in prevention of the cancers induced by the radiation. Nontoxic compounds that can protect DNA against ionizing radiation have considerable potential as radioprotectors and could be of use in preventing diseases like cancer and degenerative diseases arising from gene mutations. Some of the nontoxic natural radioprotective agents studied include cysteine, cysteamine, 5-hydroxytryptophan, melatonin, curcumin, glycyrrhizic acid, 31 gallic acid, 32 rutin, vitamins like A, C, and E, etc. Amifostine (WR-2721), disulfiram, 33 diethyldithiocarbamate, 34 troxerutin, 35 tocopherol monoglucoside 36,37 etc. comprises some of the recently investigated synthetic and semi-synthetic radioprotectors.
The present study shows that FA has potential to inhibit γ-radiation-induced DNA DSBs as observed in the micronuclei study, which was further confirmed by the γH2AX foci study. Our in vivo results with murine system indicate that, administration of FA 1 hour prior or immediately after 4 Gy γ-radiation exposures significantly inhibit micronuclei formation in mouse peripheral blood RETs and γH2AX foci, a sensitive indicator of DSBs produced by ionizing radiation. 38 Phosphorylation of histone H2AX on serine 139, γH2AX, occurs at sites flanking DNA DSBs and can provide a measure of the number of DSBs within a cell. Our γH2AX foci study showed that treatment of splenocytes with FA inhibits the formation of γH2AX foci, which is an indicator of DSBs. Protection of DNA strand breaks by FA was further studied by alkaline comet assay, which measures both single- and double-strand breaks. Our previous study indicated that administration of FA 1 hour prior to radiation exposure protect DNA in peripheral blood and bone marrow cells. 23 As shown in Figures 2 and 3, administration of FA immediately after 4 Gy γ-radiation exposure protects cellular DNA in peripheral blood leukocytes and bone marrow cells. There are many radioprotectors that have their radioprotective property when administered before radiation exposure but very limited have postirradiation radioprotective property. FA has radioprotective property when administered before or after radiation exposure. Thus FA can be beneficial during a radiation accident.
Further study was extended to explore the effect of FA on in vivo survival of Swiss albino mice. Our in vivo survival study showed that FA has potential to enhance the survival upon radiation exposure at all the tested radiation doses. The result indicates that it has an SER 2.5 at a radiation dose 6 Gy (Fig. 4).
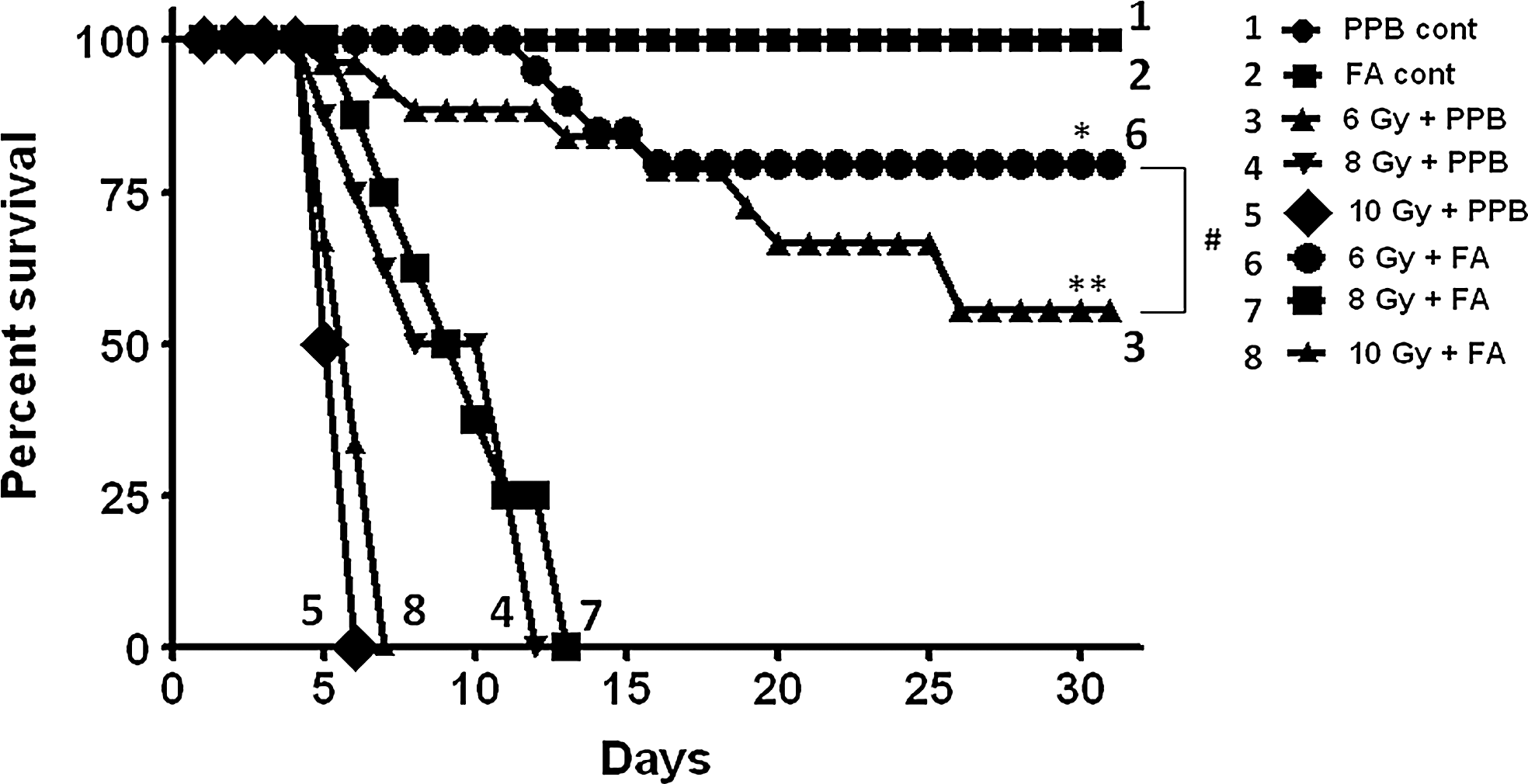
Effect of FA on survival of mice after exposure to different doses of γ-radiation. Number of animals taken per point for normal control and FA control was 5 and rest other groups was 10. The difference between the estimated survival times of the two groups was evaluated by the log-rank test. (Log-rank, *p=0.045, **p=0.0047 compared with control group and # p=0.245 between 6 Gy+potassium phosphate buffer and 6 Gy+FA).
In conclusion, the present study reveals that FA inhibits formation of radiation-induced micronuclei, which is possibly due to inhibition of DSBs as confirmed by γH2AX foci study. Postirradiation treatment of FA inhibits γ-radiation-induced DNA damage. It also enhances the survival of splenic lymphocytes and mice. Thus, it could possibly be used as an effective radioprotector during radiation accidents and during intentional exposures.
Footnotes
Acknowledgments
The authors thank Mr. Manjoor for his help in confocal microscopy and Ms. Jisha and Mr. Prayag for FACS study. The authors also thank Mr. P. M. Gonsalves, Mr. Deepak Kathole, and Ms. Pritam Patil for their help in animal experiments.
Disclosure Statement
The authors declare that there are no conflicts of interest.