
Abstract
Background:
Radiotherapy is an important treatment for the patients with advanced pancreatic cancer. Emerging studies determined apurinic/apyrimidinic endonuclease 1/redox factor-1 (APE1/Ref-1) might associate with the resistance of human pancreatic cancer cells to radiotherapy.
Aims:
To investigate whether downregulation of APE1/Ref-1 expression by ribonucleic acid interference would increase the sensitivity of chromic-P32 phosphate to pancreatic cancer cells.
Methods:
The plasmids containing APE-specific and unspecific short hairpin were transfected into Patu-8898 cells. Stable cell clones were selected by G418. The mRNA expression of APE1/Ref-1 was detected by semiquantitative reverse transcription–polymerase chain reaction and the protein expression of APE1/Ref-1 was detected by Western blot analysis; cell proliferation was studied by 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) and colony formation assay; apoptosis was detected by flow cytometry.
Results:
After 24 hours irradiation, APE1/Ref-1 mRNA and protein expression were upregulated, in a concentration-dependent manner. Suppression of APE1/Ref-1 by siRNA increased the pancreatic cancer cells hypersensitive to 32P-CP. In the combination of 32P-CP and siRNA group, MTT assay showed that the cell inhibition increased to (74.33%±9.02%), the surviving fraction in the colony formation assay was only 25.00%, and the apoptosis rate was up to (16.77%±0.98%).
Conclusions:
Knockdown APE1/Ref-1 gene expression may significantly sensitize the Patu-8988 cells to radiotherapy, which may be a useful target for modifying radiation resistance of pancreatic cancer cells to irradiation.
Introduction
Despite enormous and important progress in medicine, pancreatic cancer remains a major therapeutic challenge nowadays. Pancreatic cancer is the fourth leading cause of cancer-related death in the United States currently, with a relative 5-year survival rate of only 5%–6%. 1 The high mortality rate is due to the high incidence of metastatic disease at its initial diagnosis, the aggressive clinical course, and the failure of systemic therapies. Surgery may be the best therapy for a long-term survival, but only 15%–20% patients present with potentially resectable disease at presentation. 2 Therefore, radiation therapy and chemotherapy are the most common approaches for treating unresectable pancreatic cancer. Due to the increasing resistance of radiation therapy and chemotherapy, the median survival with radiation therapy and chemotherapy alone is only 42 and 44 weeks from the date of diagnosis. 3,4 Thus, for a better combat to this lethal and aggressive disease, investigate the novel molecular targets that are actively involved in the resistance of tumor cells to conventional therapeutics, including chemotherapy and irradiation, is necessary. One potential molecular target that has been extensively studied in numerous cancers for recent years is multifunctional apurinic/apyrimidinic endonuclease 1/redox factor-1 (APE1/Ref-1). 5 –9
Human apurinic/apyrimidinic endonuclease 1/redox factor-1 (Ape1/Ref-1, also termed APE1 and Ref-1), has both DNA repair activity and reduction–oxidation (redox) activity. Ape1/Ref-1 normally is an essential enzyme in the base excision repair (BER) pathway, which protects the cell from the cytotoxic and mutagenic apurinic/apyrimidinic (AP) sites that can accumulate in DNA via exogenous and endogenous sources. 10 Ape1/Ref-1, as a major AP-1 (Fos/Jun) reduction factor in cells, represents a novel redox component of signal transduction processes that regulates eukaryotic gene expression, activates numerous transcription factors, such as AP-1 (Fos/Jun), NF-κB, HIF-1α, CREB, p53, and facilitates their DNA binding via the reduction of a cysteine residue. 6,8 Thus, Ape1/Ref-1 is a fundamental factor in maintaining the genome and survival of cells. In contrast, Ape1/Ref-1 is constitutively active and results in overexpression of the Ape1/Ref-1 protein in multiple types of human cancers, including prostate, ovarian, cervical, germ cell tumor, rhabdomyosarcoma, and colon. 6 Its overexpression correlates with a poor outcome for chemo- and radiotherapy, a poor complete response rate, a shorter local relapse-free interval, poorer survival, and high angiogenesis. 5,7,9 Our previous study has shown an elevated level of APE1 in pancreatic cancer and gemcitabine increase the APE1 expression in a dose-dependent fashion; we significantly sensitized pancreatic cancer cells to gemcitabine and enhanced cell apoptosis when Ape1/Ref-1 expression was knocked down. 11
Nevertheless, the radioresistance of pancreatic cancer remains one of the reasons for the failure of treatment in clinical practice, which may require high doses that are beyond the normal tissue tolerance and with toxicity to the surrounding viscera, especially the small intestine. Hence, determining the cellular, molecular, and biochemical mechanisms underlying radiation resistance becomes critical for the successful treatment in this lethal disease. Phosphorus 32, which is a pure h-particle emitter with a physical half-life of 14.3 days and about 7.6 mm maximum range, comes close to the ideal unsealed therapeutic radionuclide. In present studies, we investigated whether APE/Ref-1 mRNA and protein levels were associated with colloid of chromic 32P (32P-CP), meanwhile, we investigated whether the suppression of APE1 would increase the sensitivity of pancreatic cancer cells to colloid of chromic 32P (32P-CP).
Materials and Methods
Cell culture and experimental reagents
Human pancreatic cancer Ptau-8988 cells (a gift from Ruijin Hospital, Shanghai Jiaotong University School of Medicine, Shanghai, China) were grown in the Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 mg/mL streptomycin (Gibco BRL) in a 5% CO2 incubator at 37°C.
The monoclonal antibody against human APE1 was purchased from Novus Biologicals (Novus). Anti-β-actin was purchased from Kangcheng Biological (Kangchen Biological). All APE1 siRNA and scrambled control siRNA were commercially obtained from Genepharm (Shanghai Genepharm). The DharmaFECT 1 siRNA Transfection Reagent was purchased from Dharmacon. Protease inhibitor cocktail, 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT), and all other chemicals were obtained from Sigma.
Radiation exposure and concentration treatment procedure
For concentration studies, cells were treated with 0, 0.11, 0.22, 0.45, and 0.90 MBq/mL of colloid of chromic 32P (32P-CP), respectively. At the end, reverse transcription–polymerase chain reaction (RT-PCR) and Western blot analysis were performed for the APE1 gene and protein expression.
Construction and transfection of recombinant plasmid vector and selection of stable cell clone
The pRNAT U6.1/NEO was purchased from Genescript. The siRNA targeting the APE1/Ref-1 was designed by using siRNA Target Finder on Genescript website. 12,13 Hairpin siRNA sequences were synthesized to two complementary DNA oligonucleotides annealed form BamHI and HindIII, which sticky ends in both terminals and ligated between the BamHI and HindIII sites of pRNAT U6.1/NEO, which had been cut with BamHI and HindIII. Three siRNA-transcripting DNA sequences were selected and designed in line with the nucleotide acid sequence of the APE1/Ref-1 gene, named as APEX-1siRNAI, APEX-1siRNAII, and APEX-1 siRNAIII; one unspecific sequence was designed as a negative control and named as pRNAT U6/NEO-negative-siRNA. Sequences are as follows:
APEX-1siRNAI (F: 5′
In addition, the underline position is the BamHI site, after annealed with its counterpart oligonucleotide, a Bam H1 sticky end could form in 5′ terminal of insert DNA double strands. The bold part is the HindIII site, after annealed with its counterpart oligonucleotide, a HindIII sticky end could form in 3′ terminal of insert DNA double strands. The vector also contains a PCR-amplified cDNA sequence for the green fluorescent protein (GFP).
After the plasmids were transformed into chemically competent Escherichia coli cells and after isolated plasmid DNA, a sequenator was used to sequence them; later plasmids were transfected into Patu-8988 cells with a transfection reagent (Qiagen), respectively. Patu-8988 cells were selected with G418 at the lowest killing concentration of 500 μg/mL. Cells were studied after the selected with G418 for 4 weeks. The knockdown effects of RNA interference were confirmed by RT-PCR and Western blot analysis.
Cell growth inhibition studied by MTT assay
Ptau-8988 cells (5×103) were seeded in a 96-well culture plate overnight and treated with various treatments for 24 hours: (1) the blank control group of cells cultured in the DMEM medium; (2) the negative-siRNA group of cells cultured with 50 nM scrambled control siRNA; (3) the si-APE1 (APEX-1siRNAII) group of cells cultured with 50 nM siRNA targeted to the APE1 gene; (4) the radiotherapy group of cells in culture with the addition of 0.22 MBq/mL 32P-CP; and (5) a combined group of cells cultured with 0.22 MBq/mL 32P-CP and 50 nM siRNA. After various treatments, the cells were incubated with the MTT reagent (0.5 mg/mL; Sigma) at 37°C for 2 hours according to the manufacturer's instructions. Cell viability was determined by scanning the absorbance with a microplate reader at 450 nm. Data were expressed as the percentage of viable cells and results are plotted as mean±SD of three separate experiments.
Colony formation and clonogenic assay
For colony formation assay, 1×104 cells were seeded on every 60-mm dish overnight and subsequently treated as the same as the above mentioned in MTT assay. After 14 days of incubation, plates were gently washed with PBS, fixed with ethanol, and stained with Giemsa. Colonies consisting of >50 cells were manually counted. Plating efficiency and the surviving fraction were calculated according to the following formula:
Apoptosis detected by flow cytometry
To determine the apoptosis, cells were seeded in six-well culture plates overnight and subsequently treated as the same with the above mentioned in MTT assay. After various treatments, both floating and trypsinized adherent cells were collected and washed in cold PBS twice and stained with Annexin V-APC/7-AAD (7-Amino-actinomycin) (BD PharMingen) in accordance with the manufacturer's protocol. Analysis was performed immediately by using a flow cytometer.
Reverse transcription–polymerase chain reaction
Total RNA extractions were performed by using the TRIzol reagent (Invitrogen) and treated with RNase-free Dnase I. The total RNA was reverse transcribed and amplified by PCR. APE1 gene sequence retrieved from PubMed and synthesized by Invitrogen Co. The corresponding APE1 primers were forwards: 5′-ACTTCAGGAG CTGCCTGG ACT-3′, reverse: 5′-AATGCAGGTAACAGAGTGGGA-3′ and the predicted product was 564 bp. β-actin was used as the house-keeping gene and its primers were forwards: 5′-CACCCACACTGTGCCCATC–3′and reverse: 5′-CCACA GGACT CCATGCCC-3′. The predicted product was 342 bp. PCR for the APE1 and β-actin gene was carried out as described in our previous study. 11
Western blot analysis
Western blot analysis was performed by using standard techniques, as described previously in our study. 11 Briefly, equal protein aliquots in each sample were resolved in 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and the proteins transferred into nitrocellulose membranes. After blocking with 5% skimmed dried milk, the membranes were incubated with a 1:2000 dilution of primary mouse monoclonal antibodies against human APE1 and β-actin. The membranes were then incubated with a horseradish peroxidase-conjugated secondary antibody (1:2000; Pierce, Thermo Fisher Scientific, Inc.). The proteins were detected by an enhanced chemiluminescence detection system (SuperSignal West Femto Substrate; Pierce), and light emission was captured on Kodak X-ray films.
Statistical analysis
Results are expressed as mean±SD. The statistical significance of differential findings between experimental and control groups was determined by one-way ANOVA. Post hoc tests were used at the second stage of ANOVA and the S-N-K test was used to get the p-value. p<0.05 would be considered significant.
Results
32P-CP induces concentration-dependent upregulation of APE1 expression
Patu-8988 pancreatic cancer cells were incubated for 24 hours with 0, 0.11, 0.22, 0.45, and 0.90 MBq/mL of 32P-CP, respectively. To investigate the association of APE1/Ref-1 levels and the irradiation dose in Patu-8988 pancreatic cancer cells, we performed RT-PCR and Western blot. As seen in Figure 1A and B, the APE/Ref-1 mRNA expression was elevated significantly and positively related to the irradiation dose of 32P-CP when irradiated for 24 hours, as determined by RT-PCR. Western blot analysis showed irradiation induced APE/Ref-1 protein expression in a concentration-dependent manner (Fig. 1C, D).

Evaluation of the effects of various concentrations of 32P-CP on the apurinic/apyrimidinic endonuclease 1/redox factor-1 (APE1/Ref-1) expression in Patu-8988 pancreatic cancer cells. The APE/Ref-1 mRNA
RT-PCR and Western blot detection of APE1 expression
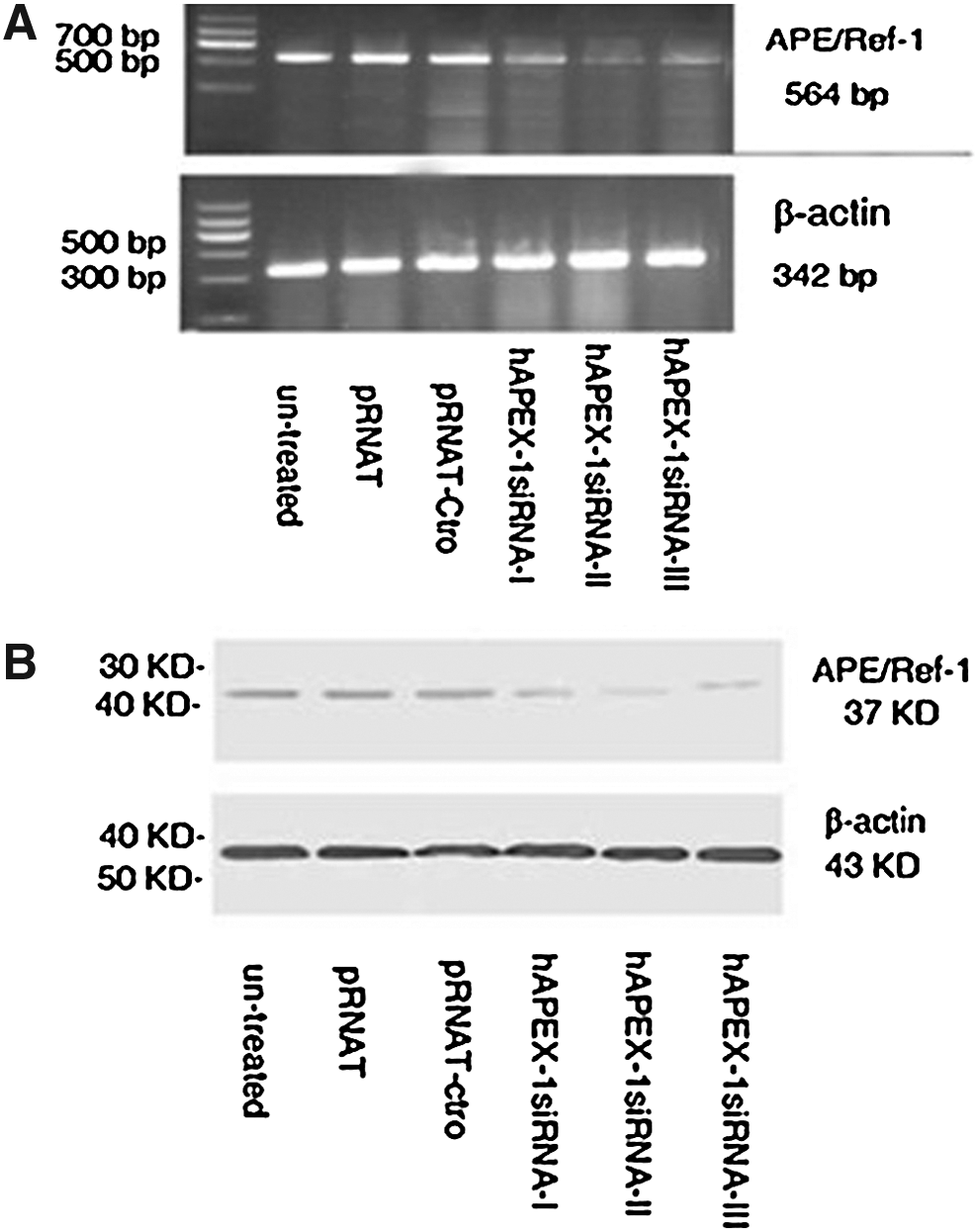
The manifestation figure of the transfection vector is showed in Figure 2. The cells that could express the GFP protein were successfully transfected with the transfection efficiency as follows: the hAPEX-1siRNA-I group (58.03%±3.17%), the hAPEX-1siRNA-II group (51.02%±7.92%), the hAPEX-1siRNA-III group (52.50%±5.14%), the pRNAT U6.1/NEO-Ctro group (50.02%±4.73%), the pRNAT U6.1/NEO group (52.00%±2.99%), and the blank control group without any vector transfected (0%). We further performed RT-PCR and Western blot analysis to determine that all recombinant plasmids suppress APE1 expression both in mRNA and protein levels in pancreatic cancer cells. As shown in Figure 3A, the inhibitory rates of APE1 mRNA expression were (26.47%±3.91%), (73.53%±5.13%), and (55.88%±12.31%) in Patu-8898 cells treated by plasmids hAPEX-1siRNAI, hAPEX-1siRNAII, and hAPEX-1siRNAIII, respectively. Figure 3B shows that APE1 protein expression was also significantly reduced in the hAPEX-1siRNAI, hAPEX-1siRNAII, and hAPEX-1siRNAIII groups, compared with the three control groups (the pRNAT U6.1/NEO-Ctro group, the pRNAT U6.1/NEO group, and the blank control group); the inhibitory rates were (50.00%±9.17%), (70.00%±1.98%), and (63.33%±5.22%), respectively. APE1 mRNA and protein expression did not significantly change in the three control groups. In addition, hAPEX-1siRNAII resulted in the most reduction among three siRNA groups, so we chose hAPEX-1siRNAII to knockdown the APE1 gene in the following experiments. (Fig. 3A, B)

The manifestation figure of transfection vector (×400). Transfection efficiency was about 50% in all groups except 0% in the control group without any vector transferred.
Electropherogram of the production of the reverse transcription–polymerase chain reaction and Western blot show that the APE/Ref mRNA
Knockdown of APE1 sensitizes Ptau-8988 cancer cells to radiotherapy by MTT assay
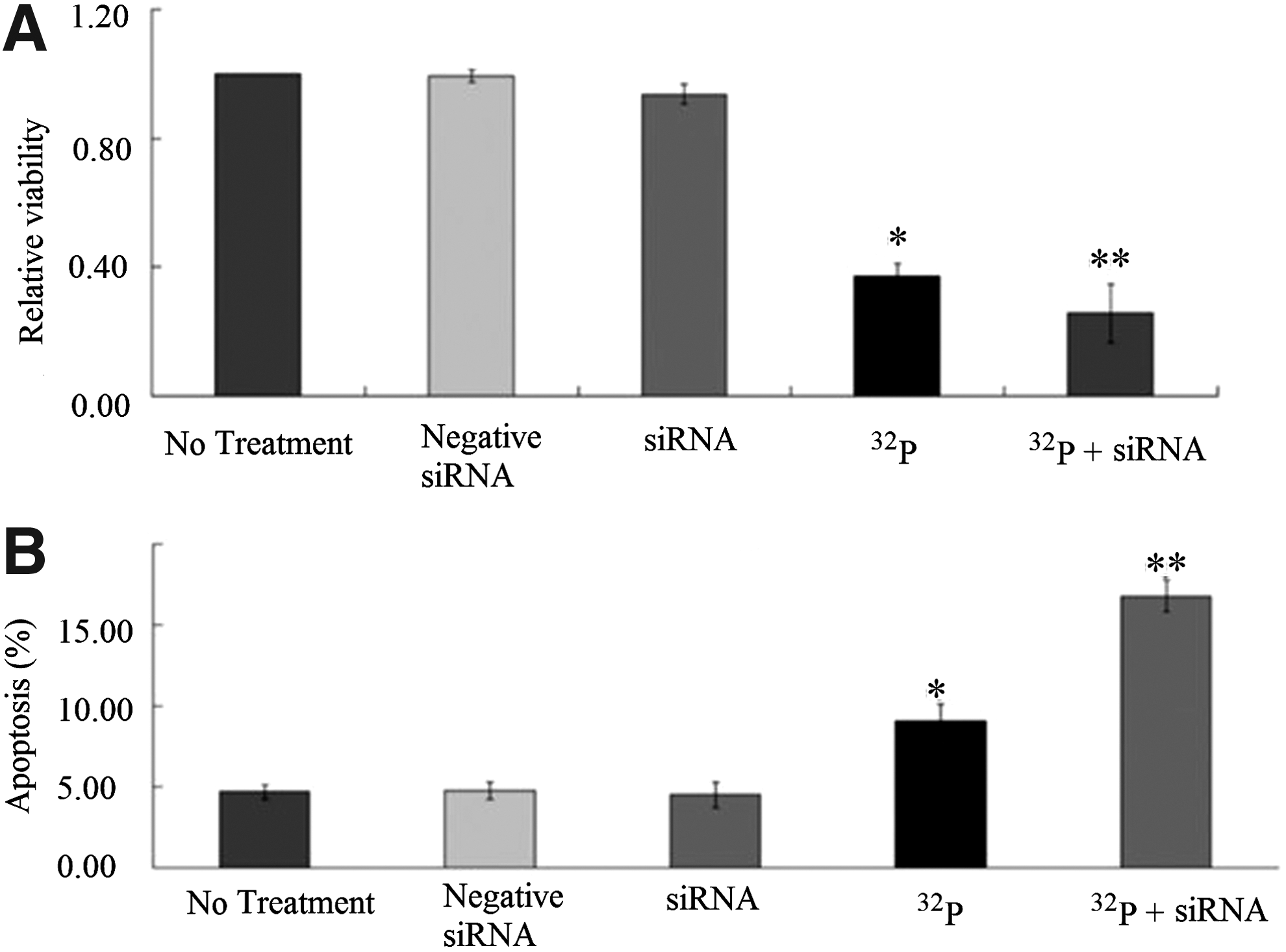
The important antiapoptotic function of APE1 suggests that downregulated APE1 expression may restore the sensitivity of pancreatic cancer cells to radiotherapy. To test this hypothesis, cells were treated with four treatments as mentioned above for 24 hours. High-dose 32P-CP together with APE1 siRNA might cause too large a degree of cell growth inhibition; thus, we chose low-dose 32P-CP. As shown in Figure 4A, the relative viability of cells were (94.67%±3.05%), (37.33%±3.79%), and (25.67%±9.02%) in the Ptau-8988 cells treated with siRNA alone, 32P-CP alone, or the combination of 32P-CP and siRNA, respectively. These mean that 32P-CP alone, or combination of both could significantly inhibit cell growth (p<0.05), and the combinatorial group produces a greater inhibition than the 32P-CP-alone group. In addition, IC50 is (0.55±0.09) MBq/mL.
Effects of the various treatments on cell growth by 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT)
Colony formation assay would be an additional confirmation of cell growth inhibition with various treatments. The plating efficiency (PE) was (70.6%±8.95%). The surviving fractions were (98.18%±3.12%) in the negative control, (91.37%±7.08%) in siRNA alone, (42.37%±10.17%) in 32P-CP alone, and (25.62%±9.33%) in the combination of 32P-CP and siRNA groups, respectively. Compared with the blank and negative control groups, 32P-CP alone and the combination both significantly inhibited cell surviving (p<0.05, Fig. 5); the combination treatment resulted in a better inhibition of cell surviving than two single agent groups. Our data demonstrated that the treatment with APE1 siRNA increased the irradiation-induced cell growth inhibition in Ptau-8988 cells.

Effects of various treatments on the clonogenic growth of Patu-8988 cells.
We are also interested in determining whether knockdown of the APE1 will increase the radiotherapy-induced apoptosis in Ptau-8988 cells. Cells were treated with five treatments as mentioned above for 24 hours; apoptosis was assessed by flow cytometry analysis. The apoptosis rate were (4.68%±0.46%), (4.77%±0.52%), (4.52%±0.79%), (9.05%±1.05%), and (16.77%±0.98%) in the blank control group, negative siRNA, siRNA, 32P-CP, a combination of both, respectively (Fig. 4B). We found that there was a greater degree of induction of apoptosis in the combination treatment group than those caused by either alone agent (Fig. 4B).
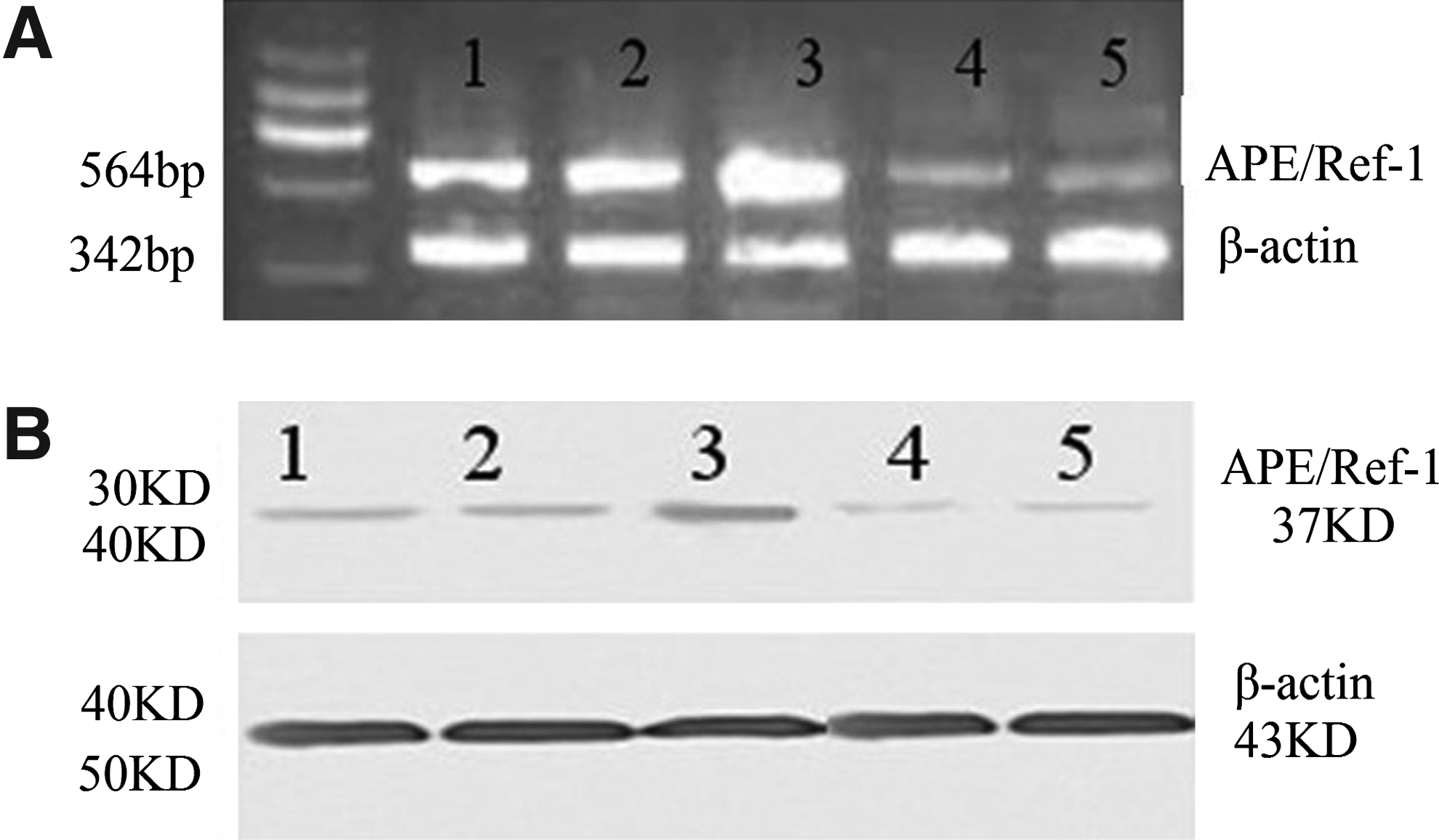
Is there a relationship between sensitivity of irradiation and knockdown of APE1 by siRNA? We further performed RT-PCR and Western blot analysis to detect the mRNA and protein expressions of APE1 in various groups. The relative expressions of APE/Ref-1 mRNA were (100.00%±7.15%), (100.00%±9.68%), (130.64%±14.44%), (30.79%±4.71%), and (23.97%±3.64%), respectively. The relative expressions of the APE/Ref-1 protein were (100.00%±8.58%), (100.00%±7.75%), (141.13%±16.53%), (35.43%±5.12%), and (26.89%±4.59%), respectively. As expected, both of the mRNA and protein expression of APE1 in the combinatorial group significantly decreased, compared with the 32P-CP-alone group (Fig. 6). In line with the above-mentioned results, the mRNA and protein levels elevated in the 32P-CP group compared with the untreated control group. The findings may suggest that there is a direct link between the sensitivity of irradiation and knockdown of APE1 by siRNA in Ptau-8988 cells.
Discussion
Radiation therapy remains a primary treatment modality for pancreatic cancer, which is a highly aggressive malignant disease and is currently treated with limited success and dismal outcomes without useful therapies. Intrinsic or acquired resistance to radiation is a major contributor to the lethal outcome. Hence, the characterization of mechanisms underlying resistance is of a significant interest to identify the potential targets for antiresistance therapies to develop potential prognostic markers to guide the adjuvant therapies. The possibility of the relationship of APE1 and radiotherapy responses has been observed in numerous tumors for several decades. Experimental data showed suppression of APE1/Ref-1 by various methods reduced the resistance of tumors and established cell lines to chemotherapeutic agents and radiotherapy. For example, DNA antisense methodology has implicated APE1/Ref-1 in cellular resistance to a variety of agents that cause alkylation and oxidative DNA damage. 6,9 Suppression of APE1/Ref-1 by specific antisense oligonucleotides increases the mammalian cells hypersensitive to gemcitabine, temozolomide (TMZ), and bleomycin. 14,15 Consistent with Fung and Demple's results, 16 we have found out that synthesized siRNA chemically directed against the APE1/Ref-1 resulted in increased proliferation inhibition and apoptosis in pancreatic cancer cells. 11 Conversely, elevation of the APE1/Ref-1 level is accompanied by decreased abasic site abundance and decreased sensitivity to chloroethylators 1,3-bis(2-chloroethyl)-1-nitrosourea (BCNU) and temozolomide. 17
In addition, emerging clinical data support that there is a link between intrinsic radioresistance and expression of APE1. One clinical study analyzed the relationship of the AP endonuclease activity and the time to tumor progression (TTP) after adjuvant therapy (alkylating agent-based chemotherapy or radiotherapy) in adult gliomas. 18 They found out that AP endonuclease activity promoted resistance to alkylation and radiation in grade II and III gliomas and it would be a useful predictor of progression after adjuvant therapies in a subset of gliomas. Similar results were noticed in cervical carcinomas. 19 Interestingly, in another report, they failed to determine the cellular levels of this enzyme in relation to survival after ionizing radiation in 11 human tumors and fibroblast cell lines. 20 In the same study, the authors also upregulated the APE1 in a rat cell line that was relatively deficient in endogenous AP endonuclease activity, but this had no measurable effect on radiation. The clinical report that mentioned above did not observe an association between AP endonuclease activity and TTP in grade IV gliomas. 18
The molecular mechanism of the process, which APE1 is responsible for escaping the deleterious effects of conventional treatments and producing the resistance to them, remains unknown. We knew that APE1/Ref-1 has two main functions: DNA repair activity and redox activity. In the BER pathway, apurinic/apyrimidinic endonuclease is a key DNA repair enzyme that cleaves DNA at cytotoxic abasic sites caused by various physical and chemical agents. APE1 accounts for >95% of the total AP endonuclease activity. 21 APE1 exhibits a 3′-phosphodiesterase activity for removal of fragmented sugar moieties, which are found at the 3′ end of DNA strand breaks induced by certain drugs, such as bleomycin, and by ionizing radiation. 22 However, the present study and the above-mentioned studies did not allow us to distinguish which function of APE1/Ref-1 is essential for the great anticancer potential. Emerging attempts of discovering specifically blocking the APE1 redox or DNA activity by small inhibitor are underway. For example, 7-Nitroindole-2-carboxylic acid (NCA) inhibits the AP endonuclease, 3′-phosphodiesterase and 3′-phosphatase activities of APE1 at low micromolar concentrations, and is a specific inhibitor of the exonuclease III family of enzymes to which APE1 belongs. 21 Lucanthone, has been used to treat Schistosomiasis, is an inhibitor of APE1/Ref-1 in repairing activity in discussion nowadays. Luo et al. 23 demonstrated that Lucanthone potentiated the activity of MMS and TMZ, probably due to its inhibition of the DNA repair activity by APE1/Ref-1. Alkoxyamine derivative Methoxyamine (MX) was another attractive inhibitor of APE1/Ref-1's DNA repair activity and was also widely researched in many experimental studies. MX potentiated the cytotoxicity of TMZ in various tumor cells, such as colon, ovaries, and breast as well as in colon cancer xenograft models by blocking Ape1/Ref-1 cleavage. 9,24 –26
As importantly, APE1/Ref-1 also functions as a redox factor, which maintains the transcription factors in an active reduced state and stimulates the DNA binding activity of numerous transcription factors that are involved in cancer promotion and progression, such as AP-1 (Fos/Jun), NF-κB, hypoxia-inducible transcription factor (HIF-1α), CREB, p53, and others. 27 Generally, inhibition of APE1/Ref-1's redox activity will result in the inability of these transcription factors to bind with DNA, and therefore, it would stop the tumor cell signaling of angiogenesis and uncontrolled growth. Some studies have established that the small molecule inhibitor E3330 was a selective antagonist of the APE1/Ref-1 redox domain function, 28,29 which inhibits the growth and the migration and the angiogenesis of pancreatic cancer cells. 30,31 Additional potential modulators of redox signaling via APE1/Ref-1, soy isoflavones, acts as a radiosensitizer and potentiates prostate tumor cell killing by radiation both in vitro and in vivo. 32 However, a clinical report did not found that the addition of soy isoflavones to gemcitabine and erlotinib increased the survival rate of patients with advanced pancreatic cancer. 33 Thus, the use of this compound is questionable nowadays. Another natural occurring compound with reported effects on the redox activity of Ape1/Ref-1 is resveratrol, which is found in various fruits and vegetables, including grapes, peanuts, berries, and pines. 9 Resveratrol suppressed cyclooxygenase-2 (COX-2), cytochrome p450, and c-fos expression accounting for its ability to inhibit tumor promotion. 34 –36 Resveratrol also induced cell cycle arrest at the G0-G1 phase with concomitant decrease of various tumor cell lines at the S or G2/M phase contributes to the antiprogression ability. 37
For our knowledge, this is the first report that correlated APE1 levels with radioresistance in pancreatic cancer cells. We conclude that knockdown APE1/Ref-1 enhances the sensitivity of pancreatic cancer cells to radiation. However, RNA interference possessed some potential toxic side effects in the metabolic process; small molecule inhibitors may be a safer and more effective choice. Therefore, we are performing an experiment to detect the effects of the combinations of resveratrol, a specific inhibitor of APE1/Ref-1 and radiotherapy in our laboratory. Consistent with the present study, we have got similar results.
Footnotes
Acknowledgments
Supported by Shanghai Leading Academic Discipline Project, Project Number Y0205 and the 2009 Doctoral Fund of the Ministry of Education of China, Project Number 20090073120092.
Disclosure Statement
No competing financial interests exist.