
Abstract
Purpose:
Human 8-oxoguanine DNA glycosylase (hOGG1) is an ubiquitous protein. It initiates the DNA base excision repair (BER) pathway to repair the 8-oxoguanine lesion. This may be associated with chemotherapeutics. In this article, the effect of hOGG1 over-expression on cisplatin resistance in esophageal squamous cell carcinoma (ESCC) EC9706 and ET13 cells was investigated.
Methods:
Recombinant adenovirus pAd/CMV/V5-DEST-hogg1 and control adenovirus pAd/CMV/5-GW/lacZ were constructed and transferred into EC9706 and ET13 cells, respectively. The protein expression and localization were determined by Western blot and by immunofluorescence assay. The cell growth viability was determined by 3-(4,5-dimethylthiazol-2yl)-2,5diphe-nyltetrazolium bromide (MTT) assay and clonogenic survival assay. The apoptotic cells were detected by terminal deoxynucleotidyl transferase mediated dUTP nick end labeling (TUNEL) staining and flow cytometry. The oxidative DNA damage (8-Hydroxyguanine [8-oxoG] DNA level) was semi-quantified by immunohistochemistry assay.
Results:
The over-expression of hOGG1 protein was mainly in the nucleus in hOGG1 cells. After exposure to a common chemotherapeutic agent cisplatin, hOGG1 over-expression cells exhibited longer survival ability, lower cell apoptosis, and less 8-oxoG oxidative damage, compared with vector-treated cells and no-treated cells (p<0.05).
Conclusion:
BER pathway to repair 8-oxoG lesion may be associated with ESCC sensitivity to cisplatin, and over-expression of hOGG1 in the nucleus can repair more 8-oxoG oxidative damage. The findings implied that over-expression of hOGG1 can protect ESCC cells from cisplatin-induced apoptosis and prolong cancer cell survival time. Modulation of DNA damage repair activity in the nucleus or in the mitochondria may lead to a different approach regarding cisplatin-induced resistance to chemotherapy.
Introduction
Esophageal cancer is one of the common digestive malignancies in the world. There is a very high incidence of the esophageal squamous cell carcinoma (ESCC) in China. More than half of the ESCC patients are in China. 1,2 Although cisplatin is one of the active cytotoxic agents and has been extensively used in the treatment of cancer, cisplatin resistance of tumors impacts therapeutic results severely. Many studies suggest that cisplatin could mediate antineoplastic effects via DNA adduct formation. These platinum-DNA lesions are mainly removed by nucleotide excision repair (NER). 3 Mismatch repair (MMR) can actually lead to cell death by induction of mitotic crisis. 4 Some evidence indicates that platinum compounds also induce reactive oxygen species (ROS) production in a selected number of cells, resulting in lipids, proteins, nuclear and mitochondrial DNA (mtDNA) damage. 5,6
One of the most stable and deleterious ROS-modified products of oxidative DNA damage is 8-Hydroxyguanine (8-oxoG), also known as 8-oxo-7,8-Dihydroguanine. The unrepaired 8-oxoG lesions in DNA can lead to A/8-oxoG mismatches during DNA replication and result in G:C to T:A mutations. This type of mutation is common in several suppressor genes, including K-ras oncogene in human cancers. 7 The repair of A/8-oxoG mismatches is subject to processing by base excision repair (BER). The initial step in BER is usually catalyzed by formamidopyrimidine DNA glycosylase (Fpg) or MutM in Escherichia coli, 8 or 8-oxoguanine glycosylase (OGG1) in mammalian cells, or its human homologue OGG1 (hOGG1), which removes the chemically altered guanines. 9 These enzymes cleave the N-glycosylic bond linking the altered base to the DNA back bone and have an associated lyase activity that nicks the DNA backbone 3′ to the lesion. Studies have suggested that deficiency in hOGG1 repair capacity contributes to the accumulation of DNA damage and increases genome instability, thus maybe leading to human carcinogenesis. 10 Evidence has shown that cisplatin exposure increases intracellular ROS in tissues, and cisplatin-mediated toxicities are inhibited by over-expression of hOGG1 or its functional homolog, E. coli Fpg in human embryonic kidney cells. 11 A recent study showed that cells defective in BER display a cisplatin-specific resistant phenotype. 12
After oxidative stress and cisplatin exposure, hOGG1 is a specific target of the Ca2+-dependent protease Calpain I. 13 Therefore, in addition to NER and MMR, hOGG1 expression involving BER may also affect cell resistance to cisplatin. Zhang et al. 14 reported over-expression of hOGG1-2a in the mitochondria-sensitized hepatoma cells to cisplatin. However, over-expression of hOGG1-1a in the nucleus may be reverse. This concept has not been tested.
Our recent results showed that there were different expressions of hOGG1-1a among ESCC patients. Here, we explored whether over-expression of hOGG1-1a affects sensitivity of ESCC cells to cisplatin using EC9706 and ET13 cell lines as models. Our results suggested that the BER pathway to repair 8-oxoG lesion is associated with ESCC resistance to cisplatin, and over-expression of hOGG1 in the nucleus can repair more 8-oxoG oxidative damage. The findings imply that over-expression of hOGG1-1a in the nucleus can protect ESCC cells from cisplatin-induced cell apoptosis and, thus, enhance the survival of ESCC cells.
Methods
Adenoviral vector construction
Over-expression hogg1 in the nucleus was from the pcDNA3.0-hogg1 plasmid containing hogg1-1a type (a gift from Dr. Wei, Johns Hopkins University, USA). The 1090 bp segment was amplified by PCR using primer (Forward: CGGGATCCGAGA TGCCTGCCCGCG, Reverse: CGGGATCCGAGATGCCTGCCCGCG) to introduce a Kozak sequence and inserted into the BamHI and XhoI cloning site of the pENTR3C entry vector (Invitrogen). The segment was sequenced to confirm the integrity of the open reading frame of hogg1-1a. To generate a recombinant hogg1 adenovirus vector, pAd/CMV/V5-DEST destination vector (Invitrogen) and pENTR3C- hogg1 performed an LR recombination reaction before transformation of competence DH5α (Invitrogen). The recombinant destination vector pAd/CMV/V5-DEST-hogg1 was identified by PCR with primer (provided by product information): Forward: (T7 promoter/priming site): TAATACGACTCACTATAGGG, Reverse:(V5 reverse priming site): ACCGAGGAGAGGG TTAGGGAT.
Generation of adenoviral packaging lines and virus supernatant
The recombinant destination virus pAd/CMV/V5-DEST-hogg1 and control adenovirus pAd/CMV/5-GW/lacZ were digested with PacI to expose the viral inverted terminal repeats. The PacI-digested expression clone and the control were transfected into the 293A cell line (Invitrogen), respectively; so, two kinds of crude adenoviral stocks were produced. Lipofectamine 2000 (Invitrogen) was used to transfect 293A cells. The high titers of virus solutions were obtained, and measured by plaque formation assay. The ESCC EC9706 cells were purchased from Chinese Academy of Sciences Cell Bank. The ESCC ET13 cells were a gift from Dr. Fan (University of Southeast, Medical School, Nanjing). The cells were grown in Dulbecco's modified eagle's medium (Gibco) supplemented with 10% fetal bovine serum (Gibco) and antibiotics, such as penicillin and streptomycin.
Analysis of hOGG1 over-expression by western blot
Cells were collected 3 days after transfection. The cell pellets were suspended in RIPA buffer (10 mM Tris pH 7.4, 150 mM NaCl, 5 mM EDTA, 1% Triton-X-100, and 0.1% SDS) containing protease inhibitors and 1 mM PMSF (added fresh) and were sonicated for 1 minute. After centrifugation, supernatants were collected, and total protein concentrations were measured by Bradford assay. Fifteen mg of total protein lysates were separated by 10% polyacrylamide gel and were transferred to PVDF membrane. These membranes were blocked in PBS containing 5% BSA and 0.1% Tween-20 for 1 hour and then incubated with primary antibody (hOGG1 rabbit polyclonal antibody,1:200) (Santa Cruz Biotechnology) for 1 hour at 37°C followed by incubation with horseradish peroxidase-conjugated secondary antirabbit antibodies (1:1000) (DNA lab) for 30 minutes at room temperature. Protein bands were detected using enhanced chemiluminescence method using ECL kit (Sino Biological, Inc.). The loading control was performed by stripping membranes in a buffer containing 62.5 mM Tris-HCl (pH 7.4), 100 mM mercaptoethanol, and 2% SDS and reprobed with β-actin antibody (1:200; (Buke Company). The level of protein expression was quantified by densitometry using Molecular Dynamics Image Quant software Version 4.2.
Immunofluorescent staining for hOGG1-1a protein localization
To verify and localize hOGG1-1a protein expression, cells transfected with hOGG1 or vector were grown on grass coverslips, washed with PBS, and fixed with cold-ice methanol/acetone (1:1) for 10 minutes. Auto fluorescence was quenched by 0.1% sodium borohydride in TBS for 5 minutes. The coverslips were blocked with PBS containing 10% normal goat serum and 1% bovine serum albumin for 1 hour and stained with rabbit polyclonal anti-hOGG1 antibody at 1:100 dilutions for 2 hours at room temperature followed by a reaction with green fluorescent Alexa488-conjugated antirabbit secondary antibody (1:1000; DNA lab). Cells were then examined under a fluorescence microscope equipped with a photographic system.
Analysis of short-term cell viability by MTT assay
Three days after transfection, cells were treated with cisplatin (Sigma) at different concentrations (1, 2, 4, 8, and 10 μM) for 24 hours. Otherwise, the cells were treated with cisplatin at 2 μM doses for varying time periods (0, 12, 24, 36, and 48 hours) in serum-free media. 3-(4,5-dimethylthiazol-2yl)-2,5diphe-nyltetrazolium bromide (MTT) assay (SunShineBio) was used to analyze cell viability. The colored product was measured in absorbance A at 590 nm in a 96-well plate reader by enzyme-linked immunosorbent assay (ELISA). All kinds of cells were assayed at control (no cisplatin).
Analysis of long-term cell viability by clonogenic survival assay
A clonogenic survival assay was performed as previously described. 15 Three days after transfection, cells were harvested by trypsinization, and 300 cells were plated in new 60-mm plates. These cells were then exposed to 1 μM cisplatin for 1 hour. After the 1 hour exposure, the incubation medium was replaced with fresh normal culture medium, and plates were placed in an incubator (5% CO2 at 37°C) for 10–12 days. At the end, these plates were rinsed with PBS and fixed with cold-ice methanol/acetone (1:1) for 10 minutes. The colonies formed were stained with hematoxylin, and colonies with more than 50 cells were counted under a light microscope.
8-oxoG DNA damage assessment by immunohistochemistry staining
Three days after transfection, the cells were treated with cisplatin at 10 μM for the different time periods (1, 2, and 4 hours) in serum-free media. As described in our previous publications, 16 DNA was denatured by soaking the slides in 4 N HCl. The endogenous peroxidase was blocked by incubating slides with 3% H2O2 for 30 minutes at room temperature. The slides were incubated with the primary mouse antibody anti-8-oxoG monoclonal antibody (1:100; Santa Cruz Biotechnology) at 4°C for overnight followed by HRP-conjugated secondary antimouse antibody (1:100; DNA lab) for 1 hour at room temperature. The 8-oxoG positive cells were visualized with diaminobenzamide tetrahydrochloride (DAB; SunShineBio). The cells were examined under a light microscope, and the percentage of positive 8-oxoG cells to total cells was calculated.
Analysis of cell apoptosis by TUNEL assay
Three days after transfection, the cells were treated with 10 μM cisplatin for 2 hours. The terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay was performed in slides, according to our previously reported method, 17 by using an ApopTag in situ apoptosis detection kit (Univ-bio, Nanjing, China). After treatment with 0.3% H2O2 in distilled water for 5 minutes, the sections were incubated 30 mM Tris (pH 7.2), 140 mM sodium cacodylate, and 1 mM cobalt chloride containing 0.5 U/mL terminal deoxynucleotidyl transferase buffer enzyme and 30 mM cobalt chloride biotin-16-dUTP in a humidified chamber at 37°C for 120 minutes. The reaction was terminated by incubating with 300 mM NaCl and 30 mM sodium citrate for 15 minutes at 25°C. Sections were stained with DAB/H2O2 solution and counterstained with hematoxylin. The slides were examined under a light microscope. Apoptosis index (AI) was calculated as the percentage of apoptosis cells to total cells.
Analysis of cell apoptosis by flow cytometry
Three days after transfection, the cells were rinsed with HBSS. The cells were treated with cisplatin at 10 μM for 2 hours. As per manufacturer's instructions, the cells (1×105) were stained for 10 minutes with annexin V–fluorescein isothiocyanate (FITC) and propidium iodide (Sigma) at room temperature under darkness for apoptosis analysis. Stained cells were detected, and the percentage of apoptosis cells to total cells was analyzed by flow cytometry (Ex:488 nm; Em:530 nm).
Statistical analysis
Statistical analysis was performed for significance. Results between hOGG1 expressed cells and control groups were tested using the t-test for different treatments. The significance level was adjusted at 5% for each analysis.
Results
Construction and transfection of mammalian expression vectors
The fragment sequence-inserted recombinant plasmid pENTR3C-hogg1 is similar to hogg1 gene at GenBank (NM002542). The recombinant pAd/CMV/V5-DEST-hogg1 is obtained by performing an LR recombination reaction with pENTR3C-hogg1 and destination vector pAd/CMV/V5-DEST. The 293A cell line is a permanent line from primary human embryonal kidney transformed with sheared human adenovirus type 5 DNA. E1 protein is encoded by the E1 region of adenovirus, which participates in the transactivation of some viral promoters. E1 protein is highly expressed in 293A cells. It also complements E1 deletion in recombinant adenoviral vectors pAd/CMV/V5-DEST-hogg1 and allows recombinant viral replication. Human cytomegalovirus (CMV) in pAd/CMV/V5-DEST promotes early high-level constitutive expression of hogg1 gene in EC9706 and ET13 cells. The control vector pAd/CMV/V5-GW/lacZ is used as an expression control in EC9706 and ET13 cell lines. The titers of viral supernatants of pAd/CMV/V5-DEST-hogg1 and pAd/CMV/V5-GW/lacZ are 2.75×107 IU/mL and 2.8×107 IU/mL, measured by plaque formation assay, respectively.
Increased hOGG1 protein levels in EC9706 and ET13 cells harboring hogg1
To express different levels of hOGG1 in EC9706 and ET13 cells, we transfected EC9706 and ET13 cells with recombinant pAd/CMV/V5-DEST-hogg1 and control vector pAd/CMV/5-GW/lacZ, respectively. Then, hOGG1 cells and vector cells were obtained. To investigate whether hogg1-1a transgene was over-expressed in hOGG1 cells, total protein samples from hOGG1 cells, vector cells, and no-infected cells were subjected to Western blot with anti-hOGG1 antibody. The results showed one strong band, ∼39 kDa (in the nucleus) and 1 weak band ∼41 kDa (in the mitochondria) in hOGG1 cells and only 1 weak bands (in the nucleus) in vector controls and in no-infected cells (results shown in Fig. 1A). Otherwise, there was a difference of endogenous hOGG1 expression between no-infected EC9706 cells and no-infected ET13 cells. To confirm hOGG1 protein over-expression localization in the nucleus in hOGG1 cells, immunofluorescent staining was performed with anti-hOGG1 antibody. The results suggested that hOGG1 protein over-expression was mainly localized in the nucleus in hOGG1 cells, as results show in Figure 1B.
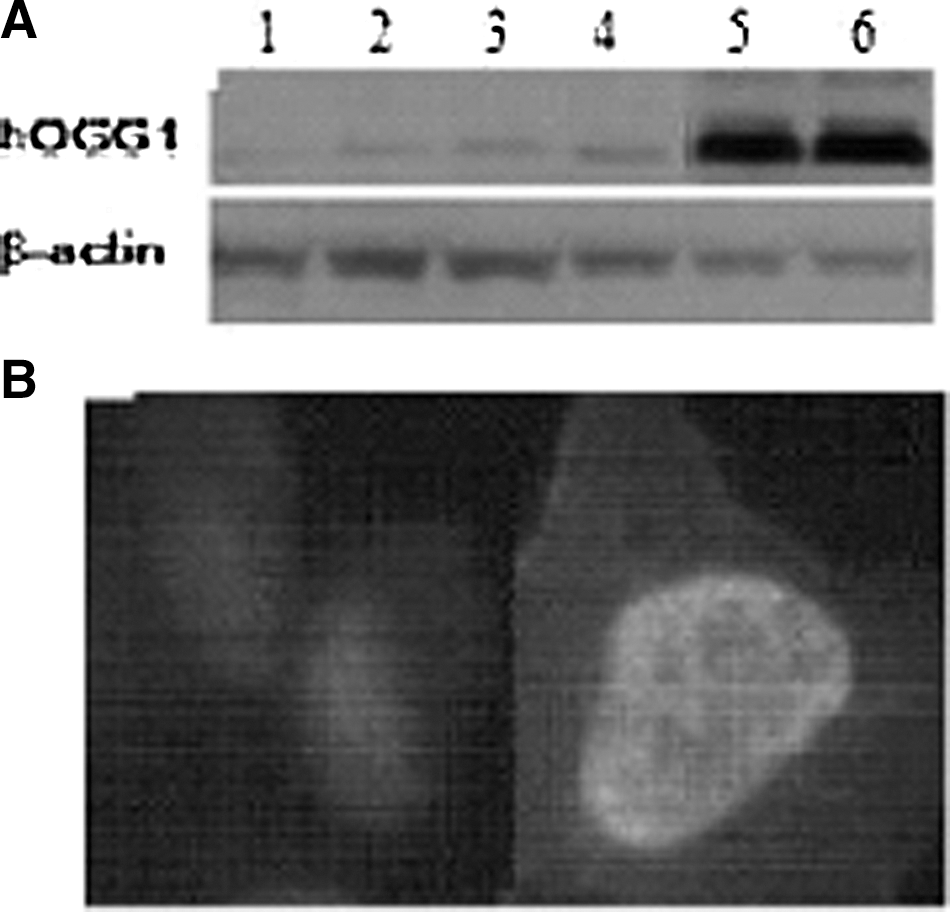
Human 8-oxoguanine DNA glycosylase (hOGG1) proteins expression by Western blot and localization by immunofluorescent stain.
hOGG1 over-expression enhances viability of EC9706 and ET13 cells to cisplatin
To determine whether hOGG1 over-expression could depress the cytotoxicity of cisplatin, we characterized the short-term survival of hOGG1 cells and vector cells and no-transfected cells, after cisplatin treatment at different concentrations (0, 1, 2, 4, and 8 μM) for 24 hours (results shown with EC9706 cells in Fig. 2A). We also treated cells with 2 μM cisplatin for varying time periods at 0, 12, 24, 36, and 48 hours, respectively (results shown with EC9706 cells in Fig. 2B). The survival rates in hOGG1 cells were much higher than those in vector cells and in no-transfected cells (p<0.05). However, MTT assay was only a transient measurement of cell viability, and clonogenic survival assay has been used for a long-term cell survival measurement. Due to the sparse plating conditions, these cells were more sensitive to cisplatin than to confluent cells. Therefore, the doses used in clonogenic survival assay were lower than the doses used in MTT assay. These cells were exposed at 1 μM cisplatin for 1 hour and then cultured for 10 days in normal medium. The colonies were not only viable but were also able to proliferate. Our results showed that hOGG1 cells possess a higher ability of colony formation than do the vector cells and no-transfected cells (p<0.05) (results shown with EC9706 cells and ET13 cells in Fig. 2C). The mean intensities were derived from three separate measurements.
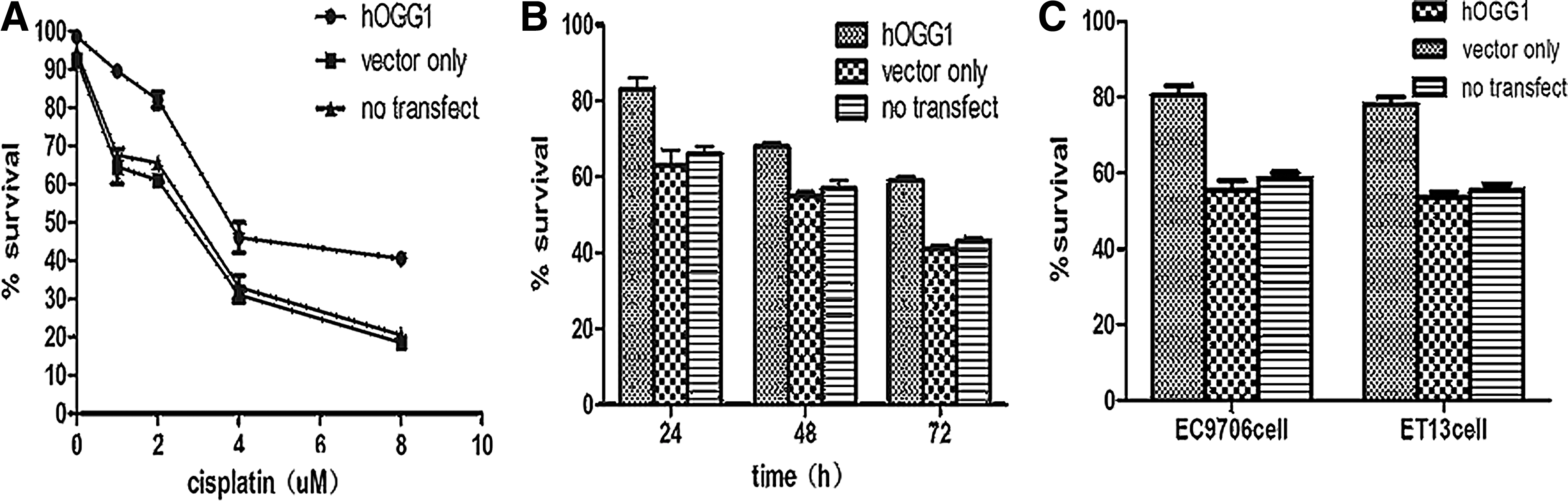
Enhanced viability of EC9706-hogg1 cells to cisplatin determined by the 3-(4,5-dimethylthiazol-2yl)-2,5diphe-nyltetrazolium bromide (MTT) assay. The short-term survival rate in hOGG1 cells or vector cells or no-transfected cells after different concentrations of cisplatin for 24 hours was analyzed by MTT assay
Decreased AI of hOGG1 over-expression EC9706 and ET13 cells to cisplatin
To determine whether the apoptotic cells in hOGG1 cells were significantly different from those in vector cells or in no-transfected cells, we calculated the percentages of three kinds of apoptotic cells with 10 μM cisplatin treatment after 2 hours by TUNEL assay and flow cytometric analysis. It measured the fragmented DNA of apoptotic cells by the enzyme terminal deoxynucleotidyltransferase incorporating biotin-16-dUTP at 3-OH DNA ends in TUNEL assay. The brown apoptotic cells were stained. As shown in Figure 3A, the number of apoptotic cells was much lower in hOGG1 cells than that in vector cells and in no-transfected cells for both EC9706 cells and ET13 cells. There was a little difference of the percent of apoptotic cells between EC9706 cells and ET13 cells. To further confirm whether the number of apoptotic cells was higher in hOGG1 cells, when comparing with vector cells and no-transfected cells, these three types of cells were stained with FITC followed by quantitative flow cytometric analysis. The percent of apoptotic cells in hOGG1 cells, and that in vector cells or in no-transfected cells after 10 μM cisplatin treatment for 2 hours were 3.95%, 11.59%, and 11.48%, respectively (results shown with EC9706 cells Fig. 3B). The former significantly decreased, compared with vector cells and no-transfected cells (p<0.05). Similar results were obtained in ET13 cells (data not shown). The results of TUNEL assay and flow cytometric analysis were similar. Three separate measurements were performed, and the percent of apoptotic cells was analyzed and displayed as the mean of these individual experiments.
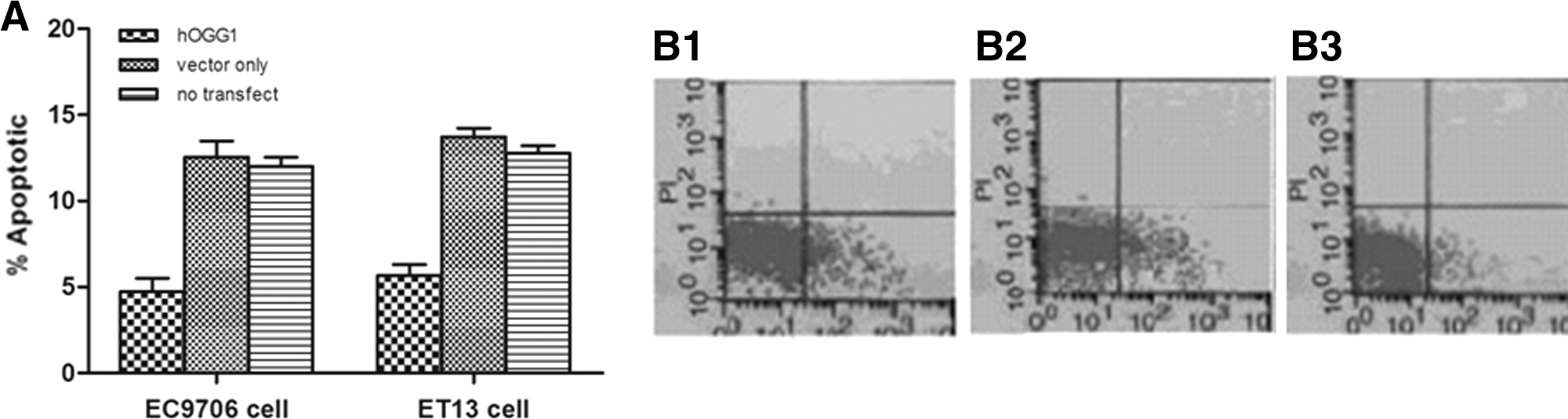
Decreased apoptosis index of EC9706-hogg1 cells and ET13 cells-hogg1 cells to cisplatin. After 10 μM of cisplatin treatment for 2 hours, apoptotic EC9706 cells and ET13 cells were evaluated by terminal deoxynucleotidyl transferase mediated dUTP nick end labeling (TUNEL) assay
Decreased 8-oxoG damage of hOGG1 over-expression in EC9706 and ET13 cells to cisplatin
To determine whether there was less 8-oxoG oxidative damage in hOGG1 cells in response to cisplatin treatment, we examined 8-oxoG oxidative damage in hOGG1 cells, in vector cells, and in no-transfected cells after cisplatin treatment at the specified concentrations (0, 2, 4, 6, and 8 μM) after 2 hours, or treatment with 4 μM cisplatin after the specified time (0, 1, 2, and 4 hours) using immunohistochemical staining for 8-oxoG positive cells with anti-8-oxoG antibody. The significant difference in the tests of overall interaction indicated that a proportion of 8-oxoG-positive cells in hOGG1 cells was significantly less than that in vector cells and in no-transfected cells (p<0.05). The differences with 4 μM cisplatin for 2 hours in hOGG1 cells, in vector-only cells, and in no-transfected cells were statistically significant (results shown in Fig. 4). There was no significant difference (P>0.05) between vector cells and no-transfected cells. Since there was a difference in endogenous hOGG1 expression between EC9706 cells and ET13 cells, a slight difference in the proportion of 8-oxoG-positive cells was in EC9706 cells as well as in ET13 cells. The mean intensities as presented here were derived from three separate measurements.
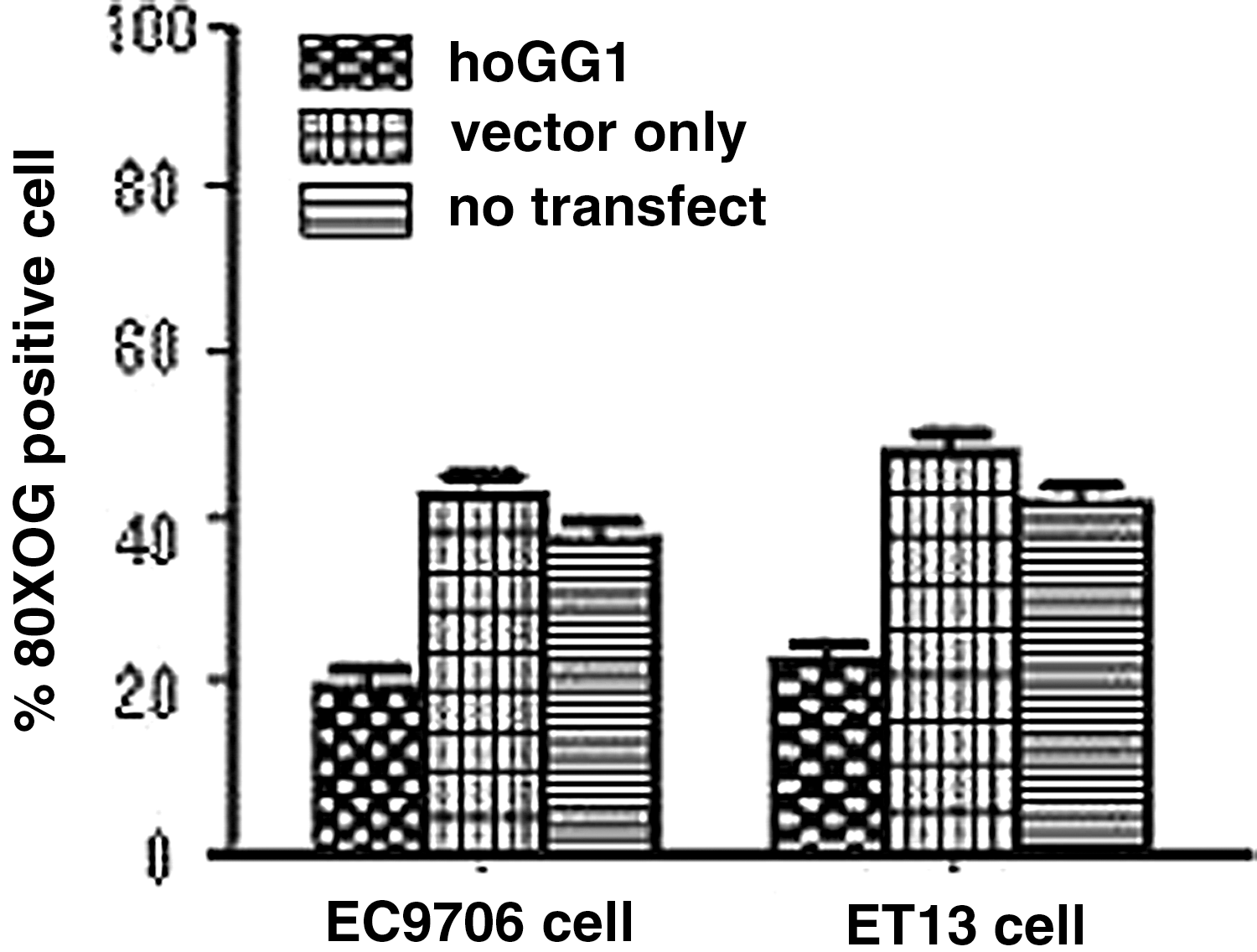
Decreased 8-Hydroxyguanine (8-oxoG) oxidative damage of EC9706-hogg1 cells and ET13 cells-hogg1 cells to cisplatin. Proportions of 8-oxoG oxidative damage EC9706 cells and ET13 cells were shown by immunohistochemical staining in hOGG1 cells or vector cells or no-transfected cells with 4 μM cisplatin for 2 hours.
Discussion
The hOGG1 is a ubiquitous and remarkably multifunctional protein of BER. The hogg1 gene lacks TATA or CAAT boxes, and is considered a housekeeping gene. 9 Defective 8-oxodG repair may lead to accumulate oxidative base damages and increase the risk of carcinogenesis. Loss of heterozygosity of hogg1 is found in several malignant tumors, such as ESCC. 18 A high accumulation of 8-oxodG DNA damage is related to age and chronic oxidative stress in ogg1-deficient mice. 19 The sequence variants in hogg1, such as Ser326Cys, are involved in repair activity and may be an important determinant of inherited susceptibility to cancer in humans. 20
Altered expression of hOGG1 and high accumulation of 8-oxodG oxidative DNA damage are often observed in several human tumors. We have previously shown a selective decrease in BER pathway in combined reflux and methyl-n-amyl nitrosamine–induced esophagus cancer in Sprague–Dawley rats. hOGG1 expression is several folds up in adenocarcinoma of rat esophagus, but there is significantly decreased expression in squamous cell carcinoma of rat esophagus. 16,17 Our recent results showed that there are different levels of hOGG1 expression among ESCC patients. In this study, we explored the association between hOGG expression levels in ESCC cells and tumor response to cisplatin therapy.
The hogg1 gene locates on chromosome 3p25.2 and spans 16.7 kb, with eight exons. Types 1a and 2a of seven alternatively spliced forms of hogg1 are major in various tissues. They are also called α-hogg1 and β-hogg1. Seven types of hogg1 carry a mitochondria targeting sequences (MTS), which consists of residues 9–26 at the common N-terminal region, but only hOGG1-1a has a nuclear localization signal (NLS) in the C-terminal end. 21 hOGG1-1a (α-hOGG1) proteins are mainly present in the nuclear and repair nuclear DNA damage, and hOGG1-2a (β-hOGG1) proteins are localized in mitochondria and play the role of mtDNA repair. 22 Zhang et al. 14 reported that over-expression of hOGG1-2a was mainly in the mitochondria. In this study, we targeted the foreign hogg1-1a gene to the nucleus by adenovirus. hogg1-1a has an NLS in the C-terminal end and an MTS in the N-terminal region. After transfection with hogg1-1a gene, hOGG1-1a over-expression and localization in hOGG1 cells were analyzed by Western blot and immunofluorescent staining with anti-hOGG1. Our results demonstrated that over-expression of hOGG1-1a were both in the nucleus and in the mitochondria, while hOGG1-1a may be mainly expressed in the nucleus. Shinmura et al. 23 also suggested that OGG1-1a (in the nucleus) was mainly expressed in cancerous and noncancerous human cells. Otherwise, there are differences of endogenous hOGG1-1a expression in different ESCC cells. Our recent results have shown that there are different kinds of hOGG1-1a expression among ESCC patients. After exposure to a chemotherapeutic agent cisplatin, BER is associated with ESCC resistance to cisplatin 12 and repairs more extensive 8-oxoG DNA damage. hOGG1 over-expressed tumor cells, thus, exhibit decreased sensitivity to the therapeutic agent cisplatin.
However, over-expression of hOGG1-1a in the nucleus may be reverse. It is noteworthy that some investigators demonstrate that over-expression of hogg1 in TK6 human lymphoblast cells increases the number of double-strand breaks in DNA and increases cellular sensitivity to irradiation. 24 Some reports suggested that over-expression of hOGG1-2a encoded by mitochondrial isoforms in mitochondria may promote mtDNA damage by creating an imbalance in the BER pathway and sensitize hepatoma cells to cisplatin. 14 When cells were exposed to oxidative damage, over-expression of mutant hogg1 in both nucleus and mitochondria resulted in decreased cellular survival, and mitochondrial-targeted mutant hogg1 resulted in more cell death than nuclear-targeted mutant hogg1. 25
Cisplatin is one of the most active cytotoxic agents that is used in the treatment of ESCC. A number of studies have suggested that cisplatin strongly inhibits DNA replication by the formation of mono- or difunctional DNA adducts in the position N7 of adenine or guanine. These adducts can form inter-strand bridges between DNA double strands and modify the 3-dimensional structure. 5,6 DNA repair plays an important role in removing cisplatin adducts. In addition, cisplatin is known to induce cell apoptosis and activate intracellular calpain. Its toxicity is also mediated partly by inactivating glutathione and antioxidant enzymes such as superoxide dismutase, catalase, glutathione peroxidase, and glutathione reductase, leading to ROS production. Therefore, cisplatin elevates genomic 8-oxoG damage, calcium influx, and calpain activation. hOGG1 is a target for calpain-mediated degradation in vivo during oxidative stress and cisplatin-induced apoptosis. 10 Our results suggest that over-expression of hOGG1-1a is not beneficial for cisplatin treatment of ESCC patients.
hOGG1 is a multifunctional DNA repair enzyme of BER that possesses DNA lyase activity and deoxyribose phosphotase (dRPase) activity. The dRPase activity of hOGG1 is to hydrolyze the dRP for subsequent enzyme activities. 26 Thus, hOGG1 plays different roles in BER. A high expression of hOGG1 protein in the transferred cells not only might enhance the release of ROS-modified bases, but also might facilitate DNA repair triggered by other DNA glycosylases.
It has been reported that BER repair capacity influences the effect of cisplatin-based chemotherapy. BER involves the tightly coordinated function of four main enzymes, including glycosylase, apurinic/apyrimidinic endonuclease (APE), polymerase, and ligase. APE1 over-expression in nonsmall cell lung cancer or in ovarian cancer is associated with the resistance of the cisplatin-based chemotherapy, and patients with a high expression level of APE1 displayed short survival times. 27,28 In addition, the inhibition of APE1 protein using RNA interference resulted in increasing sensitivity to cisplatin, H2O2, ionizing radiation, alkylating, and oxidizing DNA damaging agents. 29 –31 Our results are consistent with these reports.
The other experiments demonstrated that over-expression of hOGG1 reduces DNA damage and cytotoxicity to chemotherapeutic agents. Obstacles to effective therapy include cumulative and dose-limiting side effects of anticancer drugs, especially myelosuppression and damage to normal tissues. To decrease the cytotoxicity resistance of lung epithelial cells or hematopoietic cells to 1,3-N,N′-bis (2-chloroethyl)-N-nitrosourea (BCNU) or N,N*,N-Triethylenethiophosphoramide (thio TEPA), hogg1 or its functional homolog Fpg was transferred to cells by retroviral vector. 31 –33 To reduce cisplatin and oxaliplatin cytotoxicity, BER is enhanced in human embryonic kidney cells. 11 It is the main difference between our reports and foregoing reports that over-expression of hOGG1 in tumor cells increases the resistance of cancer to cisplatin in our experiment. It is possible that chemotherapeutic agent-induced cytotoxicity involves both tumor cells and normal cells.
The high expression of hOGG1 may have significant clinical implications, particularly when combined with targeted genes to cancer tissue by gene therapy. While initial courses in cisplatin chemotherapy are usually effective, obstacles to therapy include cumulative and dose-limiting side effects of cisplatin, especially myelosuppression. Over-expression of hOGG1 in normal tissues, such as the bone marrow, has been used to reduce cisplatin-mediated toxicities. However, when disease progression occurs and tumor resistance to cisplatin treatment is a common outcome, over-expression hOGG1 in tumor tissues increases resistance to cisplatin therapy. Our recent results demonstrated that there are differences in hOGG1-1a expression among ECSS patients. 15 Postoperative routine detection of hOGG1 expression with the patients' ESCC tissues may be useful for the guidance of making the patients' chemotherapeutic schedule. Accordingly, our findings supported a further evaluation of patient individuals of different hOGG1 expression with strategies for cancer therapy.
Footnotes
Acknowledgment
This work was supported in part by the National Natural Science Foundation of Jiangsu Province (BK2008478, BK2007108).
Disclosure Statement
The authors declare that they have no competing interests.