
Abstract
Background:
Downregulation of maspin expression has been linked to bladder cancer development, and that DNA methylation may be important for regulating maspin gene activation in bladder cancer cells. Thus, we attempted to explore the effects of the DNA methyltransferase inhibitor, 5-aza-2′-deoxycytidine (5-Aza-CdR), on the maspin expression and the biological behaviors in bladder cancer T24 cells.
Method:
The methylation status of maspin in T24 cells was investigated by methylation-specific polymerase chain reaction (PCR). After treated with different concentrations of 5-Aza-CdR (0, 0.25, 0.5, 1, and 2 μM), the maspin gene mRNA expression and protein expression were examined by real-time PCR and western blotting analysis. Cell proliferations were evaluated by the 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide assay. Flow cytometry was used to identify the apoptosis rates. Migration and invasive ability were determined by the transwell assay. Using the western blotting analysis, the changes of Cyclin D1, VEGF-C, VEGFR-3, MMP-2, MMP-9, caspase-3 p17, Bax, and Bcl-2 expression were measured.
Results:
Promoter DNA methylation of maspin was observed in T24 cells. The expression levels of maspin mRNA and protein in T24 cells were increased in a dose manner after treatment with increasing 5-Aza-CdR (p<0.05). The proliferation, migration, and invasion of cells were significantly inhibited with increasing 5-Aza-CdR, whereas the apoptosis was greatly increased (p<0.05). These were associated with the decreased ratio of Bcl-2/Bax, activation of caspase-3, and decreased expression of Cyclin D1, VEGF-C, VEGFR-3, MMP-2 and MMP-9.
Conclusions:
The present study demonstrates that maspin is silenced by DNA methylation in bladder T24 cells, and its expression can be reactivated by treatment with 5-Aza-CdR. 5-Aza-CdR could result in obvious inhibitions of the proliferation, migration, and invasion of T24 cells, which may serve as a potential strategy for the treatment of bladder cancer.
Introduction
Bladder cancer is one of the most common diseases among urogenital cancers. Invasive and metastatic bladder cancers are of aggressive malignancy. For these patients, combined chemotherapy after radical cystectomy is an attractive alternative, improving the therapeutic effect. However, despite recent advances in surgical and chemotherapeutic procedures, the 5-year survival rate in patients with invasive and metastatic bladder cancer remains very low. Therefore, the development of other therapeutic approaches for these patients is needed.
Maspin (a mammary serine protease inhibitor) is a member of the serpin superfamily of protease inhibitors. It has been shown to function as a tumor suppressor in several tumor types, by inhibiting tumor cell motility, invasion, metastasis, and angiogenesis. 1 However, in spite of its tumor suppressor activity, an increase of maspin expression has also been described to parallel tumor progression. 2,3 These paradoxical studies concerning maspin do not give us a clear idea about its possible role as a tumor suppressor or an oncogene. In case of bladder cancer, the function and mechanism by which maspin regulates cell growth were not clarified. Nevertheless, previous studies may provide some clues. Maspin expression was found in high quantities in normal urothelium, preserved in superficial bladder cancers, but was significantly diminished in invasive carcinomas. The low maspin expression level was coupled to an increased tumor cell growth in vivo. 4 These findings underline a potential role of maspin as a tumor suppressor in bladder cancer. Moreover, in maspin-negative RT4 bladder cancer cells, treatment with the DNA methyltransferase inhibitor, 5-aza-2′-deoxycytidine (5-Aza-CdR), resulted in distinct re-expression of maspin. 5 It shows a potential role of 5-Aza-CdR in the management of bladder cancer. However, the effects of 5-Aza-CdR on the biological behaviors of bladder cancer cells are not studied.
Based on the above descriptions, we speculate further that 5-Aza-CdR may lead to alterations in other proteins rather than maspin, which are associated with the biological behaviors of bladder cancer cells. In this study, the methylation status of maspin in bladder cancer T24 cells was confirmed. Then, different concentrations of 5-Aza-CdR were added into T24 cells, and the mRNA and protein expression of maspin was detected. The effects on proliferation, apoptosis, and migration and invasiveness of T24 cells were observed. The underlying mechanisms were also investigated.
Materials and Methods
Cell culture
The human bladder cancer T24 cell line was obtained from the Medical Research Center of Xiangya Hospital (Changsha, P.R. China). The cells were cultured in an RPMI-1640 medium (Gibco), including 10% fetal bovine serum (Gibco), in a humidified atmosphere with a 5% CO2 incubator at 37°C.
Methylation-specific polymerase chain reaction
Genomic DNA was isolated by means of a TIANamp Genomic DNA Kit (Tiangen). The EZ DNA Methylation-Gold Kit™ (Zymo Research) was used for bisulfite modification of genomic DNA. Primers for the methylated promoter of maspin were as follows: forward: 5′-AGGAGGATAAGAAT AGGTTTTACGA-3′, reverse: 5′-CCAATTACTTAAAAAACT AAAACGAA-3′; primers for the unmethylated promoter of maspin were as follows: forward: 5′-AGGAGGATAAGAATA GGTTTTATGA-3′, reverse: 5′-CCAATTACTTAAAAAACTA AAACAAA-3′. The size of both the unmethylated and methylated amplicon is 178 bp. The polymerase chain reaction (PCR) was run in the conditions as follows: 5 minutes at 95°C, then 40 cycles consisting of 30 seconds at 94°C, 30 seconds at 57°C, and 30 seconds at 72°C, and a final extension of 5 minutes at 72°C. PCR products were separated on 1.5% agarose gels and visualized by ethidium bromide staining. Three groups of T24 cells are setup as parallel samples, and H2O as a control.
Demethylation treatment
When the cell confluence reached 80%, the cells were treated with different concentrations of 5-Aza-CdR (0, 0.25, 0.5, 1, and 2 μM). The cultured cells with 0 μM 5-Aza-CdR served as a control.
Real-time PCR
Cells were harvested after 48 hours post-5-Aza-CdR treatment. Total cellular RNA was extracted from collected cells using Trizol reagent (Invitrogen). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) served as a control. The primer sequences used were as follows: maspin forward: 5′-GGTGGGGATTCCATAGAGGT-3′; maspin reverse: 5′-AATCGGCATCCACAGAAAAG-3′; GAPDH forward: 5′-CAATGACCCCTTCATTGACC-3′; GAPDH reverse: 5′- GACAAGCTTCCCGTTCTCAG-3′. Real-time PCR assays were carried out using the SYBR Green PCR Master Mix (ABI). PCR cycling conditions included 95°C for 5 minutes, 40 cycles consisting of 94°C for 20 seconds, 59°C for 20 seconds, 72°C for 20 seconds, then 72°C for 5 minutes, and finally 55°C for 10 seconds. A melting curve was generated after each PCR to determine the specificity of the PCR products.
MTT assay
The antiproliferative effect of maspin against different group cells was determined by using the MTT [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide] assay. T24 cells were cultivated in 96-well plates at a density of 1.0×104 cells per well. After overnight incubation, the reaction was stopped by addition of 50 μL dimethyl sulfoxide (Sigma) and cultured at 37°C for 4 hours. Then, the supernatant was removed, and 150 μL of DMSO was added into every well. The absorbance was recorded at a wavelength of 570 nm. Cell survival rate=(Treatment group absorbance/control group absorbance×100%).
Flow cytometric analysis of apoptosis
Cells were harvested after 48 hours post-5-Aza-CdR treatment. They were washed by PBS twice, followed by being resuspended in 1×binding buffer at a concentration of 1×106 cells/mL. The samples were incubated with 5 μL annexin-V-fluorescein isothiocyanate (FITC) and 5 μL propidium iodide for exactly 5 minutes at room temperature in the dark and then measured on a BD FACSAria cytometer (Becton Dickinson).
Transwell assay
Invasion assays were performed in a six-well transwell chamber (Corning) that contained an 8-μm pore-size polycarbonate membrane filter and was precoated with 50 mg/L Matrigel (Becton Dickinson) for the invasion assay. Cells were harvested after 48 hours post-5-Aza-CdR treatment. They were trypsinized and suspended in a serum-free medium containing 1% bovine serum albumin at a concentration of 5×104 cells/mL. A medium containing 10% fetal calf serum was added to the bottom chamber. After reculturing with 5% CO2 at 37°C for 24 hours, the transwell chambers were inverted and stained with hematoxylin. The migration assay was done in a similar manner, but without the Matrigel coating on the filters. Three independent experiments were done for each assay. Five fields were randomly selected, and the number of transmembrane cells was counted.
Western blotting
Cells were harvested after 48 hours post-5-Aza-CdR treatment. The cellular total protein was extracted by use of the Total Protein Extraction Kit (ProMab). Appropriate amounts of protein (20–40 μg) were separated on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Then, the proteins were transferred to nitrocellulose membranes (Pierce) and incubated at 37°C for 2 hours with the indicated antibodies. The specific primary antibodies were mouse Cyclin D1 antibody (Santa), goat VEGF-C antibody (Santa), rabbit VEGFR-3 antibody (Santa), rabbit MMP-2 antibody (Bioss), mouse MMP-9 antibody (Santa), rabbit caspase-3 p17 antibody (Abcam), rabbit Bax antibody (Cst), rabbit Bcl-2 antibody (Cst), and mouse GAPDH antibody (Santa). The secondary antibodies were rabbit anti-goat IgG/HRP (Zhongshan Goldenbridge), goat anti-rabbit IgG/HRP (SCBT), and goat anti-mouse IgG/HRP (Zymed). The relative levels of the target protein were represented as the density ratio versus GAPDH.
Statistical analysis
SPSS 13.0 statistical software was used for statistical analysis. Data were represented as the mean±SD. The significance of the data was determined by Student's t-test and one-way analysis of variance. The results were considered significant if the p-value was <0.05.
Results
Methylation of the maspin promoter region occurs in T24 cells
To investigate whether maspin hypermethylation was present in the bladder cancer T24 cell line, the promoter area of maspin gene was analyzed by a methylation-specific PCR analysis. We designed two pairs of methylation-specific primers to discriminate between the methylated and unmethylated alleles. Amplification of methylated DNA, but not unmethylated DNA, was observed (Fig. 1). It suggested that the maspin promoter methylation did exist in T24 cells.
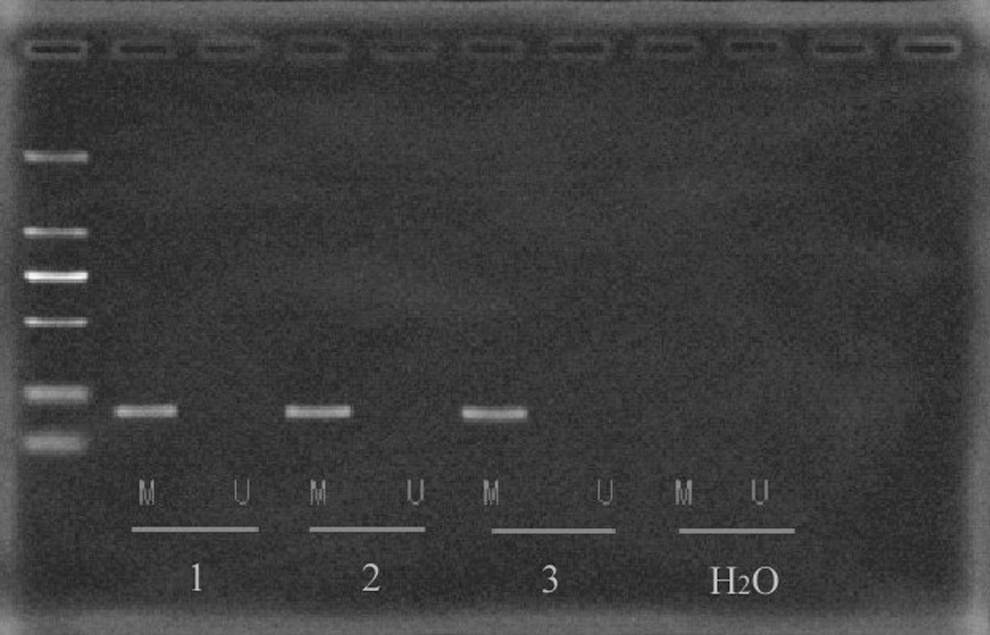
MSP analysis of maspin. The lanes indicated by M and U correspond to the products amplified by MSP primer setting specific for methylated and unmethylated DNA, respectively. 1, 2, and 3 represent the three parallel samples (T24 cells), and H2O served as a control. MSP, methylation-specific polymerase chain reaction.
5-Aza-CdR effectively enhances maspin expression in T24 cells
To examine the potential effects of 5-Aza-CdR on maspin expression in human T24 cells, cells were treated with 5-Aza-CdR at various concentrations and times. After 5-Aza-CdR treatment, maspin expression was evaluated by real-time-PCR and western blotting. As shown in Figure 2, upregulation of maspin mRNA and protein by 5-Aza-CdR was observed in a dose-dependent manner (p<0.05).
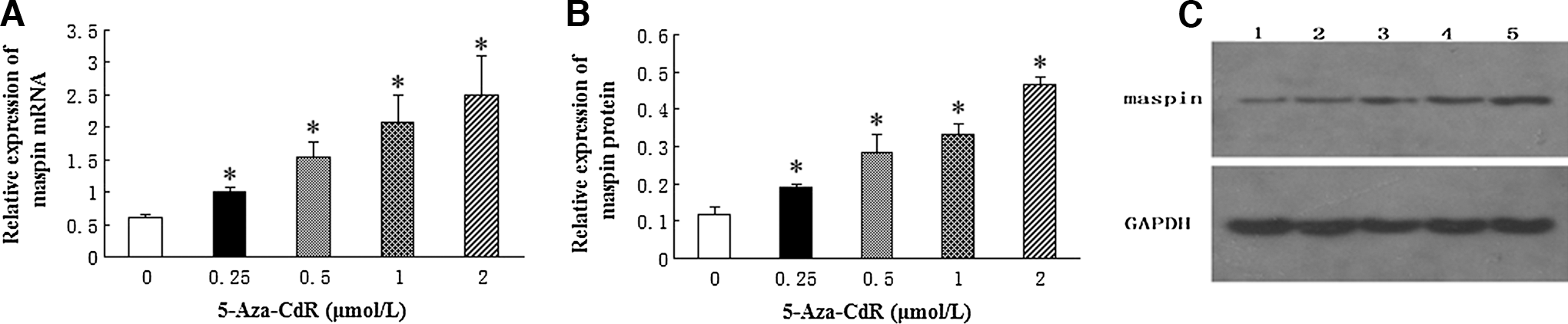
Upregulation of maspin expression in T24 cells by 5-Aza-CdR.
5-Aza-CdR significantly inhibits the proliferation in T24 cells
To investigate the cellular effects resulting from 5-Aza-CdR, the MTT assay was performed after adding different concentrations of 5-Aza-CdR to T24 cells. Data showed that 5-Aza-CdR had negative effects on cell proliferation in T24 cells. As shown in Figure 3, inhibition of T24 cell proliferation by 5-Aza-CdR was in a dose- and time-dependent manner (p<0.05).

Effects of 5-Aza-CdR on T24 cell proliferation. Reduced cell proliferations were observed with increasing 5-Aza-CdR treatment (0–2 μM) at 12, 24, and 48 hours. 5-Aza-CdR inhibited cell proliferations of T24 cells in a dose- and time-dependent manner (*p<0.05). The data are presented as means±SD (n=3).
5-Aza-CdR induces the apoptosis in T24 cells
To determine whether the 5-Aza-CdR-induced growth inhibition was mediated by apoptosis, we further used the flow cytometry to identify the cell apoptosis rate. As shown in Figure 4, T24 cells showed increasing apoptosis rates in a dose-dependent manner, including early as well as late apoptotic cell death (p<0.05). Approximately 3.21% of cells treated with 0 μM 5-Aza-CdR for 48 hours underwent apoptosis, as well as 10.45% of cells treated with 0.25 μM 5-Aza-CdR, 23.24% of cells treated with 0.5 μM 5-Aza-CdR, 38.82% of cells treated with 1 μM 5-Aza-CdR, and 50.44% of cells treated with 2 μM 5-Aza-CdR.
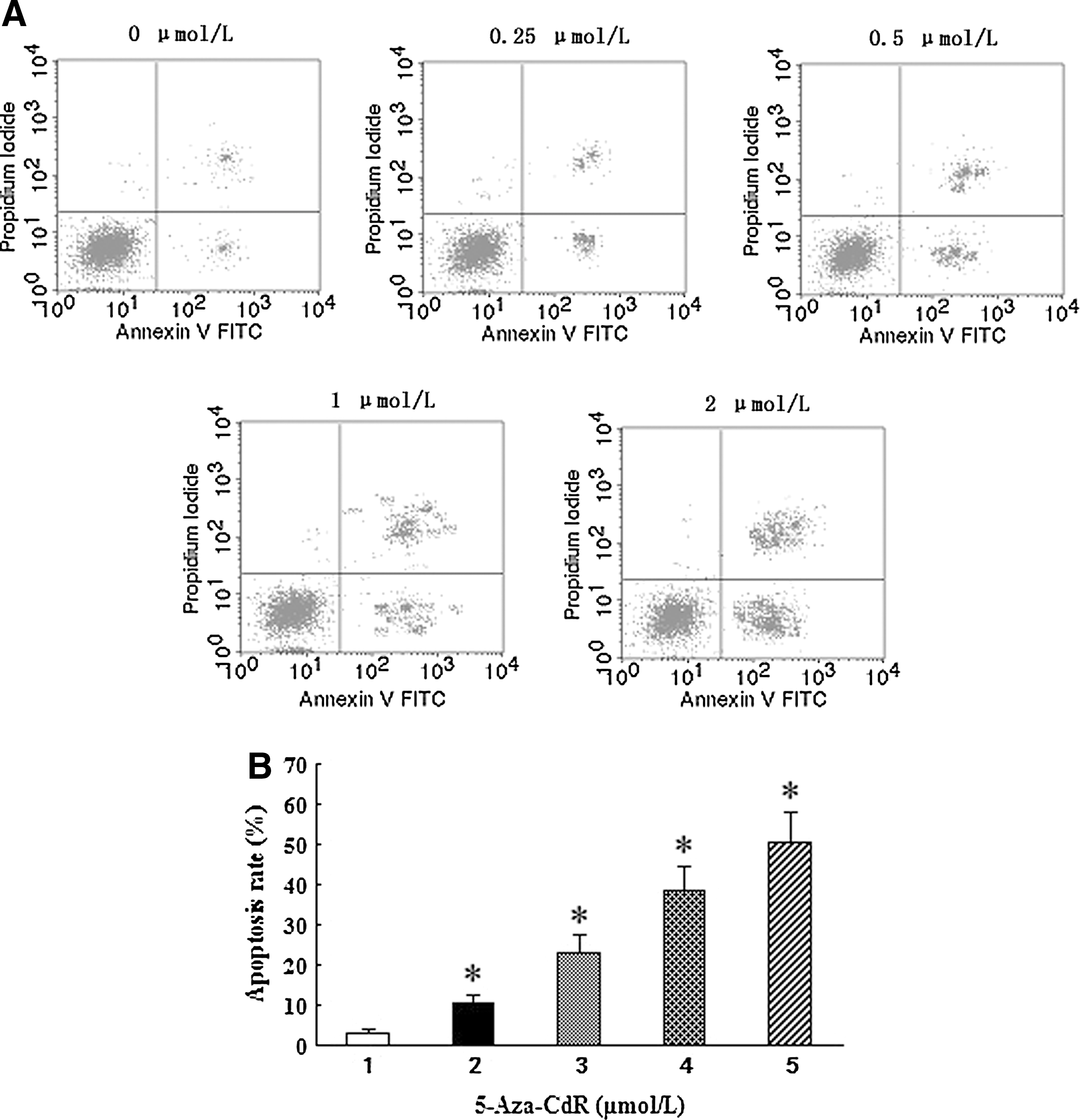
Effect of 5-Aza-CdR on T24 cell apoptosis.
5-Aza-CdR inhibits the migration and invasiveness in T24 cells
We examined the effects on the migration and invasiveness in T24 cells with increasing 5-Aza-CdR. The cells were treated with various concentrations of 5-Aza-CdR (0–2 μM) for 48 hours and analyzed by flow cytometry. The cells were observed under a microscope (original magnification ×200). As shown in Figure 5, the numbers of migrated and invasive T24 cells were significantly reduced in a dose-dependent pattern (p<0.05). This suggests that 5-Aza-CdR has a direct inhibitory effect on migration and invasion of T24 cells.
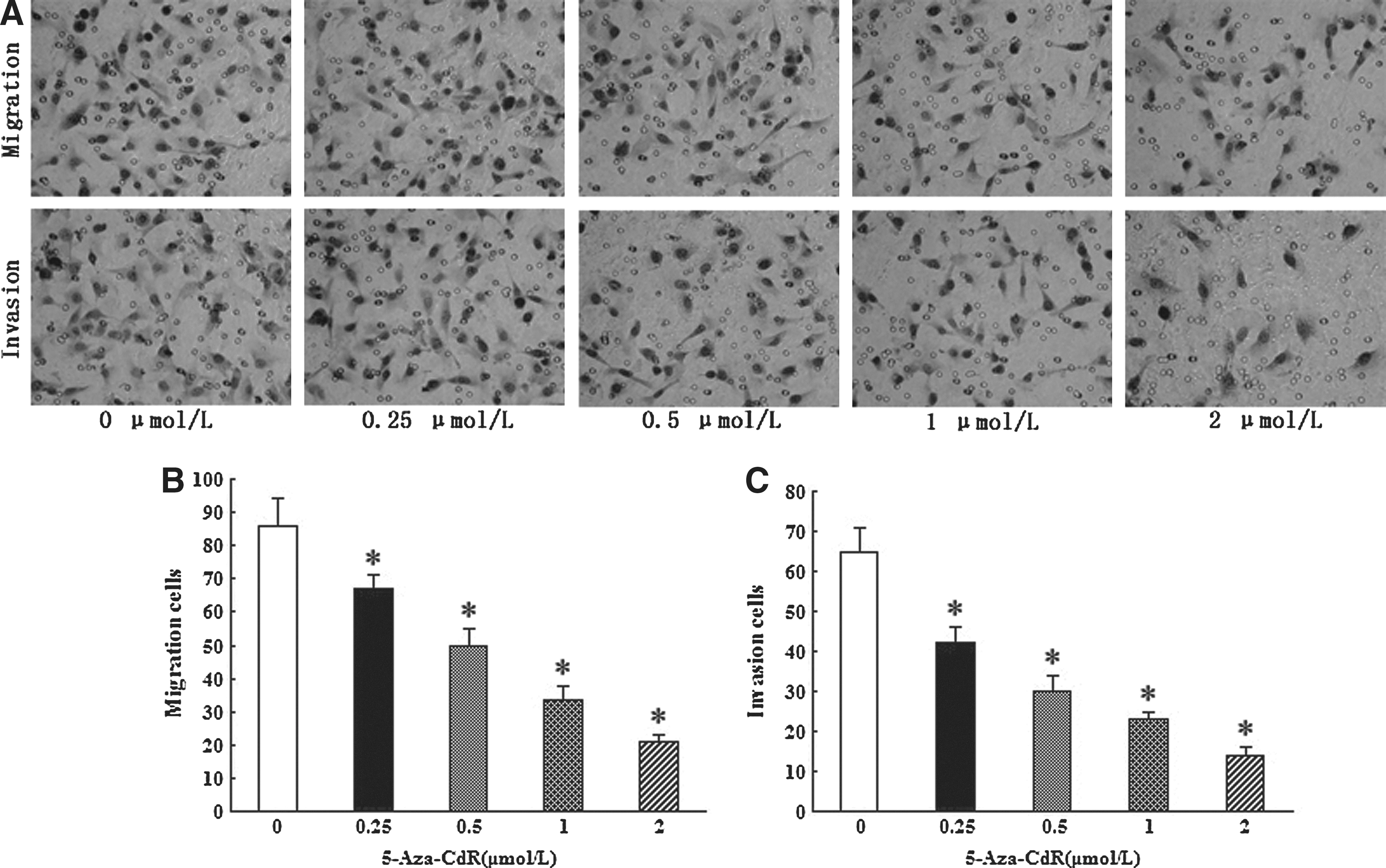
Effects of 5-Aza-CdR on T24 cell migration and invasion.
Effects of 5-Aza-CdR on the expression of tumor malignancy-related proteins
To further elucidate the probable mechanism of 5-Aza-CdR-mediated biological changes, western blotting analysis was used to examine the effects on VEGF-C, VEGFR-3, MMP-2, MMP-9, Bcl-2, Bax, and caspase-3 p17. Results showed that with increasing 5-Aza-CdR, VEGF-C, VEGFR-3, Bcl-2, MMP-2, and MMP-9 expression was significantly decreased, whereas Bax and caspase-3 p17 are greatly increased (p<0.05) (Fig. 6).
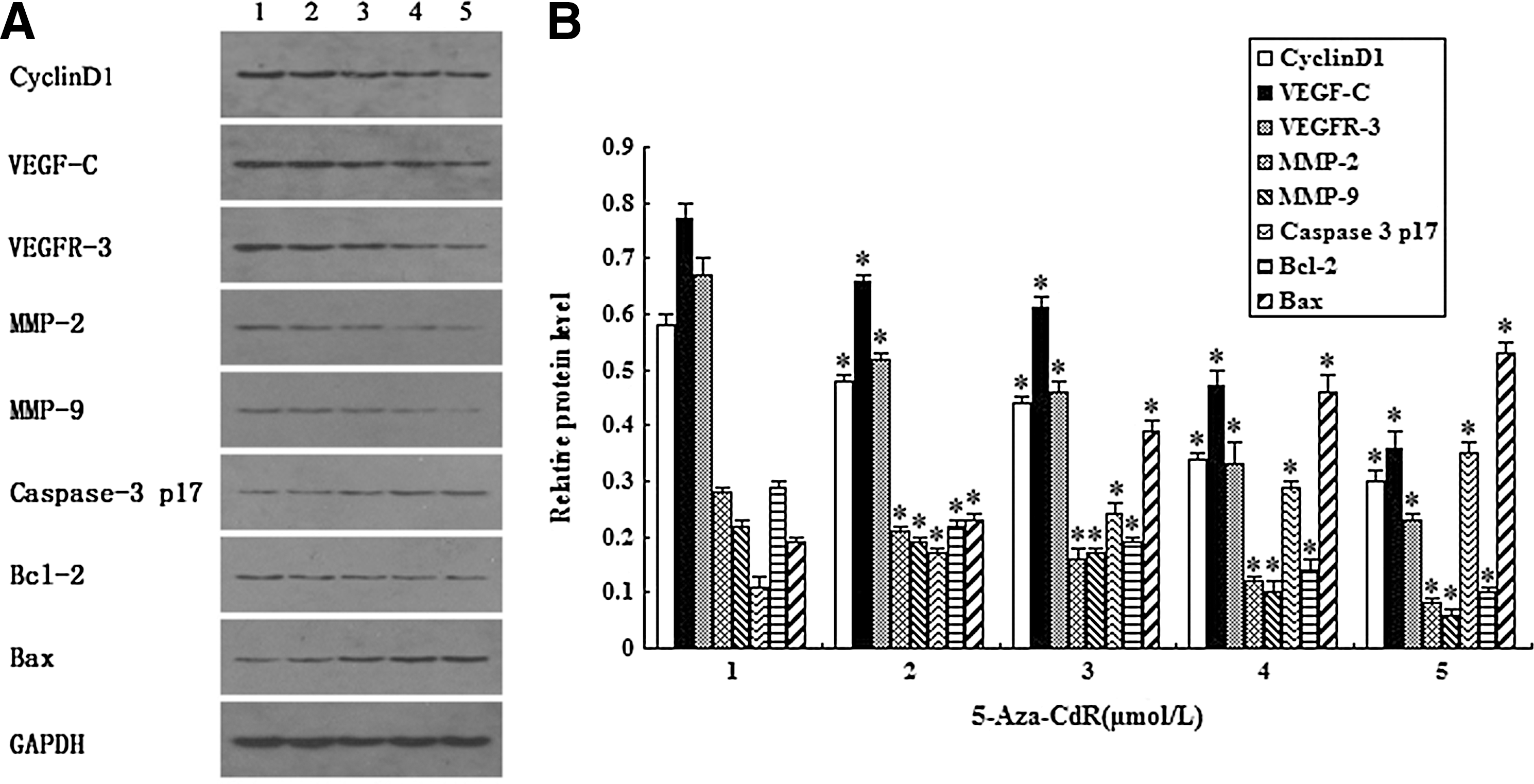
Western blotting analysis of proteins related to apoptosis, proliferation, and invasiveness in T24 cells.
Discussion
DNA methylation is the most commonly studied apparent genetic mechanism. Aberrant de novo DNA methylation may lead to abnormal gene expression and genomic instability, which are associated with cancer. 6 Epigenetic silencing of a gene can be reversed, resulting in reactivation of expression, by drugs such as the DNA methylation inhibitor 5-Aza-CdR. 5-Aza-CdR is a well-known demethylating agent in cancer therapy. 7,8 After adding to the cell culture medium, it is incorporated into the new strand during DNA replication in the cell. Then, it forms a covalent complex with the active sites of the DNA methyltransferase, depleting methyltransferase activity, which results in generalized demethylation. 9
Maspin has been characterized as a potent tumor suppressor in many in vitro and in vivo studies. DNA methylation of the promoter region is a critical mechanism for maspin gene inactivation. In a previous study, 5 maspin expression was detected in a bladder cancer T24 cell line, but not in bladder cancer RT4 cells. When RT4 cells were exposed to 5-Aza-CdR alone, maspin expression was weakly induced in RT4 cells. Whether maspin expression can also be induced by 5-Aza-CdR in T24 cell is unknown. In the present study, to address this question, we analyzed the methylation status of the promoter area of maspin in T24 cells. Our result did show that methylation of the maspin promoter region occurs in T24 cells. We then treated the cells with different concentrations of 5-Aza-CdR. Data demonstrated that 5-Aza-CdR treatment caused significantly elevated levels of maspin mRNA and protein in a dose-dependent manner. This effect is consistent with those observed in the RT4 cell line. In view of the reactivation of maspin expression by 5-Aza-CdR in T24 cells, we further explored what role 5-Aza-CdR may play in these tumor cells.
From MTT assay and flow cytometric analysis, we observed that 5-Aza-CdR suppressed the rates of cell proliferation in a dose- and time-dependent manner, which was associated with induced apoptosis. In mammalian cells, G-to-S transition in cell cycle is largely controlled by certain cyclins, cyclin-dependent kinases (CDKs), and cyclin-dependent kinase inhibitors (CDKIs). 10,11 Cyclin D1 is a major regulator of the progression of cells into the proliferative stage of the cell cycle. 12 Approximately 50% of mammary carcinomas have been shown to overexpress cyclin D1 protein. 13 To explore the role of 5-Aza-CdR in the proliferation of the T24 cells, detecting the expression of cyclin D1 was supplemented in our study. We found that decreased expression of cyclin D1 was associated with enhanced 5-Aza-CdR in our study, suggesting that 5-Aza-CdR may inhibit the proliferation of T24 cells by targeting cyclin D1. However, so far, little is known about that how 5-Aza-CdR is interacting with cyclins, CDKs, or CDKIs. It's reported that 5-Aza-CdR can suppress growth of Hep3B hepatocellular carcinoma cells dose dependently, accompanied by cyclin D1 downregulation via Cox-2 upregulation. 14 In another study, 15 p16 gene expression is increased dramatically in HepG2 hepatocellular carcinoma cells, which encodes for an inhibitor of CDK4, and was isolated and mapped to chromosome 9p21, 16,17 a locus frequently lost in diverse malignancies. 18 Based on these, 5-Aza-CdR is likely to regulate the proliferation of cells through cyclins, CDKs, or CDKIs, but the detailed mechanisms remain to be clarified.
It has been reported that 5-Aza-CdR is associated with enhanced apoptotic sensitivity of cancer cells to drug treatment, 19,20 and the links between 5-Aza-CdR and apoptotic molecules have been indicated. Liu et al. 21 showed that 5-Aza-CdR-induced apoptosis against gastric cancer BGC-823 cells was predominantly regulated via upregulation of DNA methyltransferase 1, 3a, and partially via reactivation of RUNX3, which was independent of the p53 status and its ability to activate p21Waf1/Cip1 expression. Xu et al. 22 disclosed that induction of TRAIL by 5-aza-CdR is critical for enhancing chemosensitivity of breast cancer cells to Adriamycin. Studies 23,24 revealed that 5-aza-CdR-mediated apoptosis is accompanied by changes in caspase proteins and the Bcl-2 family proteins. In the present study, our data showed that increasing 5-Aza-CdR is associated with decreased Bcl-2 and increased Bax and caspase-3 p17 subunit. In many cancers, the antiapoptotic protein Bcl-2 is found to be overexpressed, while the activity of proapoptotic Bax is counterbalanced by strong surviving signals. 25 Our finding suggests that one of the functions that 5-Aza-CdR plays is to change the level of the Bcl-2 family proteins in T24 cells. Further, the analysis revealed that the expression of the p17 subunit of caspase-3, which serves as the terminal effector molecule in many types of apoptosis, 26 increased with exposure to increasing concentrations of 5-Aza-CdR in a dose-dependent manner. Taken together, the decreased ratio of Bcl-2/Bax and activation of caspase-3 induced by 5-Aza-CdR are in line with the previous study.
Tumor metastasis is a fundamental and characteristic property of carcinogenesis, which involves adhesion, migration, and invasion. In this study, we demonstrated that 5-Aza-CdR could reactivate the expression of maspin. The clinical importance of maspin has been mainly investigated regarding tumor progression or metastasis. Hong et al. 27 demonstrated that the ability of Panc-1–maspin cells to migrate and invade through a Matrigel-coated filter was significantly reduced compared to that of Panc-1-mock cells. Shi et al. 28 showed that primary tumor growth and metastasis were significantly blocked in TM40D breast cancer cells that overexpress maspin as a consequence of plasmid or retrovirus infection. Maspin-transfected tumors tended to have tumor encapsulation and less necrosis, which were associated with better prognosis and lower invasiveness. A line of studies have been undertaken to disclose the mechanisms employed by maspin to produce its antimetastatic effects. Odero-Marah et al. 29 implied that maspin may inhibit cell motility by regulating Rac1 and subsequently PAK1 activity, and promote cell adhesion via the PI3K/ERK pathways. Qin et al. 30 uncovered a mechanism by which maspin exerts its effect on endothelial cell adhesion and migration through an integrin signal transduction pathway. Another anti-invasive mechanism reportedly utilized by maspin is that maspin reduced cell surface-associated urokinase plasminogen activator (uPA)/uPA receptor complex by inducing its internalization. 31,32 Besides, a study indicated that secretion of maspin and its deposition into the extracellular milieu deter matrix incorporation of cathepsin D, thereby reducing matrix degradation. 33 In this study, we found that increasing 5-Aza-CdR treatment is associated with decreased expression of MMP-2 and MMP-9. Extracellular matrix degradation by MMPs allows urinary bladder transitional-cell carcinoma cells to spread and diffusely infiltrate the bladder parenchyma. 34,35 MMP-2, together with MMP-9, degrades type IV collagen, the major component of the basement membrane. It has been reported that elevated expressions of MMP-2 and MMP-9 in bladder cancer tissue at the mRNA and protein levels are associated with advanced tumor stage, grade, and a decreased survival rate. 36 –38 Kanayama 36 found an elevated mRNA expression of MMP-2 and MMP-9 in tissue of muscular invasive bladder tumors compared to noninvasive tumors. It's reported that 5-aza-CdR can synergize with MPA to inhibit the invasion of endometrial cancer cells, perhaps owing to the downregulation of MMP-2 and MMP-9 expression and activity. 19 Huang showed that when the uroepithelial cells were pretreated with the inhibitor 5-aza-CdR, the upregulating effects of arsenite on RECK, MMP-9, MMP-2, uPA, and VEGF expression were suppressed. 39 These results are consistent with our study. We have previously demonstrated that VEGF-C can enhance the proliferation and invasiveness of bladder cancer T24 cells. 40 In the present study, decreased expression of VEGF-C and VEGFR-3 was also observed after 5-Aza-CdR treatment. In another study, 41 incubation of the NB cells Lan-5 with 5-Aza-2′-deoxycytidine led to the upregulation of VEGF-C mRNA expression, suggesting that the promoter of VEGF-C is methylated. These results indicate that the VEGF-C/VEGFR-3 axis may be involved in the inhibition of 5-Aza-CdR in the development of bladder cancer.
In conclusion, we demonstrated that 5-Aza-CdR treatment increased maspin expression in the bladder cancer T24 cell line and inhibited tumor proliferation, migration, and invasion in vitro. These findings indicated that 5-Aza-CdR may represent a potential therapeutic strategy to inhibit the progression of bladder cancer. To the authors' knowledge, the present study is the first comprehensive study to report 5-Aza-CdR-mediated induction of maspin expression and inhibition of development in the bladder cancer T24 cell line, as well as the functional mechanism. However, it's only an in vitro experiment using one kind of cell line. Further researches, especially in vivo experiments with multiple cell lines, are needed to verify the effects and clarify the mechanism by which 5-Aza-CdR is involved in the development of bladder cancer.
Footnotes
Acknowledgments
This study was supported by the Natural Science Foundation of Hunan Province (No. 11JJ2040) and the College students' innovation project of Central South University (No. 18) and the booting project of young teachers of central colleges (2012QNZT104).
Disclosure Statement
The authors declare that they have no conflicts of interest.