
Abstract
Cisplatin (CP)-induced neurotoxicity is one of the major clinical problems in CP-based chemoradiotherapy, leading to its discontinuation depending upon their severity. In the present investigation, the photosensitizing property of riboflavin (RF) has been used to ameliorate the CP-induced neurotoxicity. According to dosing plan, the healthy mice were given RF, CP, and their combinations under photoillumination with their controls without any light exposure. After the treatment, antioxidant enzymes, cellular reductants, glutathione-S-transferase, brain markers, and oxidation products were assessed besides histopathology in their brain samples. These parameters revealed that RF ameliorates CP-induced neurotoxicity in a dose-dependent manner under photoillumination. Hence, inclusion of RF in CP-based chemoradiotherapy can be an effective strategy to counter CP-induced neurotoxicity.
Introduction
Cisplatin (CP) is one of the most commonly used chemotherapeutic drugs against various forms of cancers and solid tumors. Many side effects are associated with the patients undergoing CP-based long-term chemotherapy that includes nephrotoxicity, hepatotoxicity, neurotoxicity, ototoxicity, and many others. 1,2 The severity of these toxicities restricts its clinical use. CP induces neurotoxicity mainly in the form of peripheral neuropathy that has been manifested by neuropathic pain, sensory impairment, sensory ataxia, demyelination, and axonal degeneration of the nerve cells. The risk and the severity of neuropathological conditions depend on cumulative dose of the drug and duration of the therapy. 3
The anticancer activity of CP is attributed to its capacity to bind with N-7 of purine residues of cellular DNA, leading to formation of platinum–DNA adducts, which are further transformed into intra- and inter-DNA cross links in the target cells. If these lesions are not repaired timely, they can derail replication and transcription machinery of the cells that can trigger cell death as the final consequence, 4 although the exact mechanism of CP-induced neurotoxicity is still not vivid. Many literatures proclaim that DNA–platinum complex products are intensely found in the target nerve cells such as in sensory cells and dorsal root ganglion (DRG) cells leading to various pathological conditions of the nervous system. 5 Besides, CP is also known to generate reactive oxygen species (ROS) and reactive nitrogen species (RNS) upon accumulating in the target cells that can alter the normal physiological redox status of the cells resulting into such deleterious effects, including the drug-induced neurotoxicity. 6,7
Riboflavin (RF) or vitamin B2 is an essential vitamin occurring in all forms of life. It participates in various enzyme-catalyzed metabolic redox reactions in the forms of FAD and FMN. Furthermore, it is also a strong photosensitizer that can generate bioactive free radicals such as RF radicals, superoxide anions, and hydroxyl radicals. 8 RF has been used in ribophototherapy and photodynamic therapy extensively and effectively since long both in intact as well as in its photoactive form. 9 –11 Previous investigations indicate that RF deficiency can increase the susceptibility of animals, including humans, to cancer as well as it can affect the efficacy of anticancer drugs such as methotrexate in vivo negatively. 12 As CP is the drug of choice for treatment of various solid tumors and malignant cancers for chemotherapy as well as chemoradiotherapy, we were interested to investigate whether the photosensitizing property of RF can exploited to reduce CP-induced neurotoxicity. Recently, our laboratory has demonstrated that RF can blunt the toxic/damaging effects of CP in mice keratinocytes under photoilluminated condition. 13 Moreover, we also observed this ameliorative effect on the CP-induced nephrotoxicity and hepatotoxicity equally effective in vivo as well. 14,15
Neurotoxicity is one of the least-attended toxic abuses of this drug that is a challenge of paramount importance in cancer research. Our present work is aimed to investigate effect of RF supplementation on the neurotoxic effect of CP on high dose in healthy mice. As a great deal of literature indicates the involvement of oxidative and nitrosative stresses in CP-induced neurotoxicity; hence, we have tried to analyze the redox status post-treatment followed by histopathological examination in the treated mice to assess CP-induced neurotoxicity in the present investigation.
Materials and Methods
Chemicals
RF, CP, reduced and oxidized glutathione, nicotinamide adenine dinucleotide phosphate reduced (NADPH), and nicotinamide adenine dinucleotide reduced were bought from Sigma-Aldrich Chemical Company. Succinic acid, potassium dihydrogen and monohydrogen phosphate, glycine, pyrogallol, hydrogen peroxide, trichloroacetic acid, and ethylenediaminetetra-acetic acid were purchased from Qualigens Fine Chemicals.
Animal treatment and sample preparation
In the treatment, thirty-six adult Swiss albino male mice of 6 months weighing 48–50 g were purchased from the Central Animal House, Jamia Hamdard University, New Delhi, India. They were kept in sufficiently large cages and treated under humane and hygienic conditions (25°C±5°C and 12 hours day: night cycle maintained) according to Departmental Ethics Committee for Animal Experimentation, permitted under the registration number of 714/02/a/CPCSEA, issued by Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), Ministry of Environment and Forests, Government of India. The mice were acclimatized for 10 days before the treatment on standard pellet mice diet (Ashirwad Industries) and clean drinking water ad libitum.
They were randomly divided into six groups taking six mice in each group. They were designated as group I (control normal), group II (control), group III (RF [2 mg/kg body weight] treated), and group IV (CP [2 mg/kg body weight] treated), and the combination of CP and RF was given to group V (2 mg/kg body weight of CP with 1 mg/kg body weight of RF) and group VI (2 mg/kg body weight of CP with 2 mg/kg body weight of RF), respectively. All the doses were injected intraperitoneally with 1-mL syringe, and saline was used as vehicle for the treatment chemicals. RF was injected before CP at the interval of ½ hour in the combinational groups—V and VI. The control, group II, was given only saline under photoillumination, while the control normal (group I) was kept as such. Thus, groups II, III, IV, V, and VI were exposed to full-body irradiation by Philips fluorescent light from ∼10-cm distance at a fluence rate of 38.6 w/m2 during the day for 12 hours. The dorsal surface, including the head region of all the mice (except group I), was mildly shaved every week for maximum possible absorption of light through the skin. Along with these six groups, two additional unshaved combination groups—V′ (2 mg/kg of CP+1 mg/kg of RF) and VI′ (2 mg/kg of CP+2 mg/kg of RF)—without photoillumination were also maintained. The mice were given a daily injection of dose for 3 days followed by a gap of a week, then again a daily dose for 3 days with a week gap, and finally three more daily injections to mimic the current treatment strategy for the cancer patients. The treatment regime was chosen to study the chronic effect of the treatment at a moderately toxic dose of the drug. Their body weights were measured weekly. The treatment was planned in such a way that all the animals were sacrificed on the same day by the cervical dislocation method.
After the sacrifice, their brains were washed with a chilled saline buffer. Two brain samples from each group were separately kept in 10% formalin for their histopathological analysis. The rest of them were weighed followed by cutting them with clean scalpel blade separately. All of them were homogenized and then centrifuged separately at 3000 g in a potassium phosphate buffer (pH 7.36, 0.1 M) and stored in separate glass vials in refrigerator. At the time of homogenization, 1 mL of homogenate of each brain sample was separately saved for estimation of reduced glutathione (GSH) and melondialdehyde (MDA) levels. All the samples were labeled properly and kept in −4°C till further analysis.
Effect of the treatment on the body and brain weight of the mice
The body weights of all the mice were monitored weekly throughout the treatment and just before the sacrifice to observe the effect of the treatment on them. Their brain weight was also noted after the sacrifice to observe the effect of the treatment.
Assay of antioxidant enzymes
The activity of different antioxidant enzymes were assayed with standard protocols. Cu–Zn superoxide dismutase (CuZnSOD) was assayed by autoxidation of pyrogallol, 16 while the activity of catalase (CAT) was assayed by decomposition of hydrogen peroxide. 17 Glutathione reductase (GR) activity was carried out by the method involving oxidation of NADPH into NADP+ in presence of oxidized glutathione. 18
Estimation of level of MDA and nitric oxide
The extent of lipid peroxidation was estimated by the method of Beuge and Aust 19 involving the measurement of total MDA, one of the major products of lipid peroxidation. Nitric oxide (NO) level was measured by a Quantichrom™ Nitric Oxide Assay kit (DINO-250).
Estimation of GSH level and –SH groups
The level of GSH was estimated by a method of Jollow et al., 20 while the total sulfhydryl groups (–SH) were measured by the Sedlak and Lindsay method. 21
Assay of major brain function markers
The activity of glucose-6-phosphate dehydrogenase (Glu-6-PDH) and lactose dehydrogenase (LDH) was assayed by standard protocols. 22,23
Assay of glutathione-S-transferase activity
The activity of glutathione-S-transferase (GST) was assayed by the Habig et al. method. 24
Histopathological study of brain samples of the treated mice
For histopathology, carefully excised brains of the treated mice were kept in 10% formalin for immersion fixation. Their tissue blocks (10×5×3 mm of size) were processed for paraffin embedding. Their sections of 7-μm thickness were cut with rotary microtome and stained with Hematoxylin and Eosin stain. The sections were observed under a trinocular light microscope (Olympus BX40), and their photomicrographs were snapped at a magnification of ×400.
Statistical analysis
All the data have been expressed in the present article as mean±standard error of mean for 6 different preparations. Their statistical significance was evaluated by one-way analysis of variance analysis by GraphPad Prism 5 software. The probability of occurrence was selected at p≤0.05. The treatment and the experiments were repeated to check reproducibility of the results.
Results
After and during the photoillumination, no erythema or other visible change on skin was observed. The mice were sacrificed by the cervical dislocation method upon completion of the treatment, and their brains were taken out of their skull on the same day. The brains were washed with cold saline and homogenized in potassium phosphate buffer (pH 7.36, 0.1 M). Later, they were centrifuged to get supernatants that were stored at −4°C to be used for different biochemical assays and estimations. The treatment and the experiments were repeated to confirm reproducibility of the results. Similar pattern of results with minor fluctuations were observed in the parameters taken on repetition.
The control normal (group I) without shaving of its dorsal body surface that was neither given any treatment nor exposed to photoirradiation showed highest activities of all the antioxidant enzymes followed by group II (taken as the control). Group I was maintained to ruleout the possible consideration that light alone was affecting the parameters taken under study in the animals. Hence, the changes observed in the parameters in the treated mice are because of the combined effect of dose of the test chemicals as well as the exposure of light.
Effect of the treatment on brain weight
The mice under the CP treatment (group IV) showed maximum drop in brain weight by 13% followed by group III with respect to the control (group II). The combination groups V and VI exhibited normalization of the brain weight by 6.6% and 12.5%, respectively, as compared to group IV (Fig. 1). The combination groups without photoillumination, that is, group V′ and VI′, showed no significant gain in brain weights, although group VI′ exhibited increase in brain weight by 4.8% (Fig. 1).
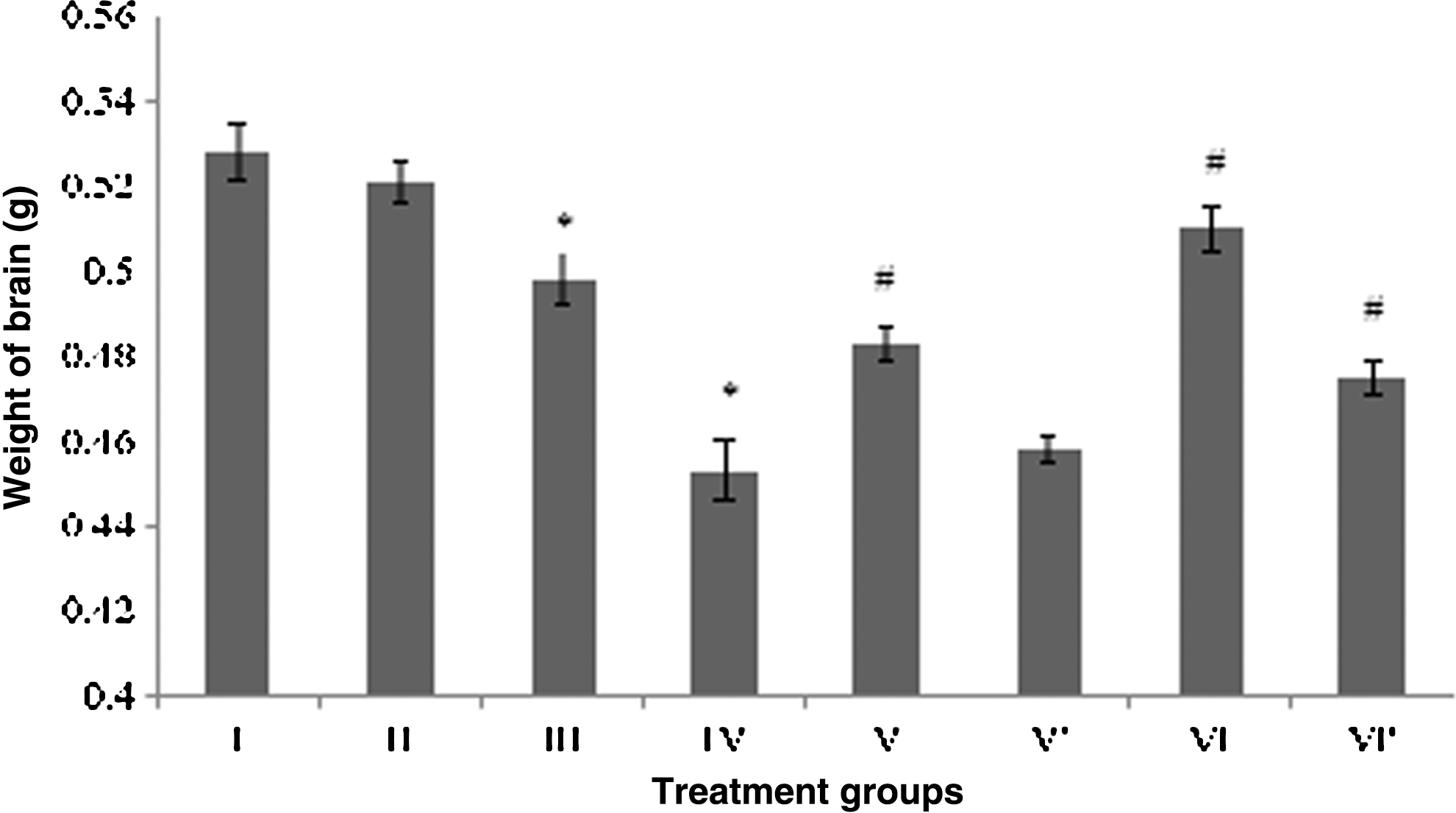
Effect of the treatment on the brain weight. Group I: control normal (neither treatment nor light exposure). Group II: control (treated with saline under photoillumination). Group III: riboflavin (2 mg/kg) treated under photoillumination. Group IV: cisplatin (2 mg/kg) treated under photoillumination. Group V′: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) under photoillumination. Group V′: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) without photoillumination. Group VI: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) under photoillumination. Group VI′: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) without photoillumination. * indicates significantly different from control at p≤0.05; # indicates significantly different from group IV at p≤0.05.
Effect of the treatment on antioxidant status of the brains
All antioxidant enzymes (SOD, CAT, and GR) after CP administration were found significantly compromised as shown by group IV, followed by group III, with respect to the control. Among the enzymes, SOD was found to be the most affected enzyme by the treatment in CP-treated group IV showing 49% decreased activity, while GR and CAT displayed the dip in the activity by 48% and 37%, respectively, in the same group. RF treatment also decreased the activity of all the three enzymes as evidenced by group III, but it was considerably less than the CP-treated group (Fig. 2).
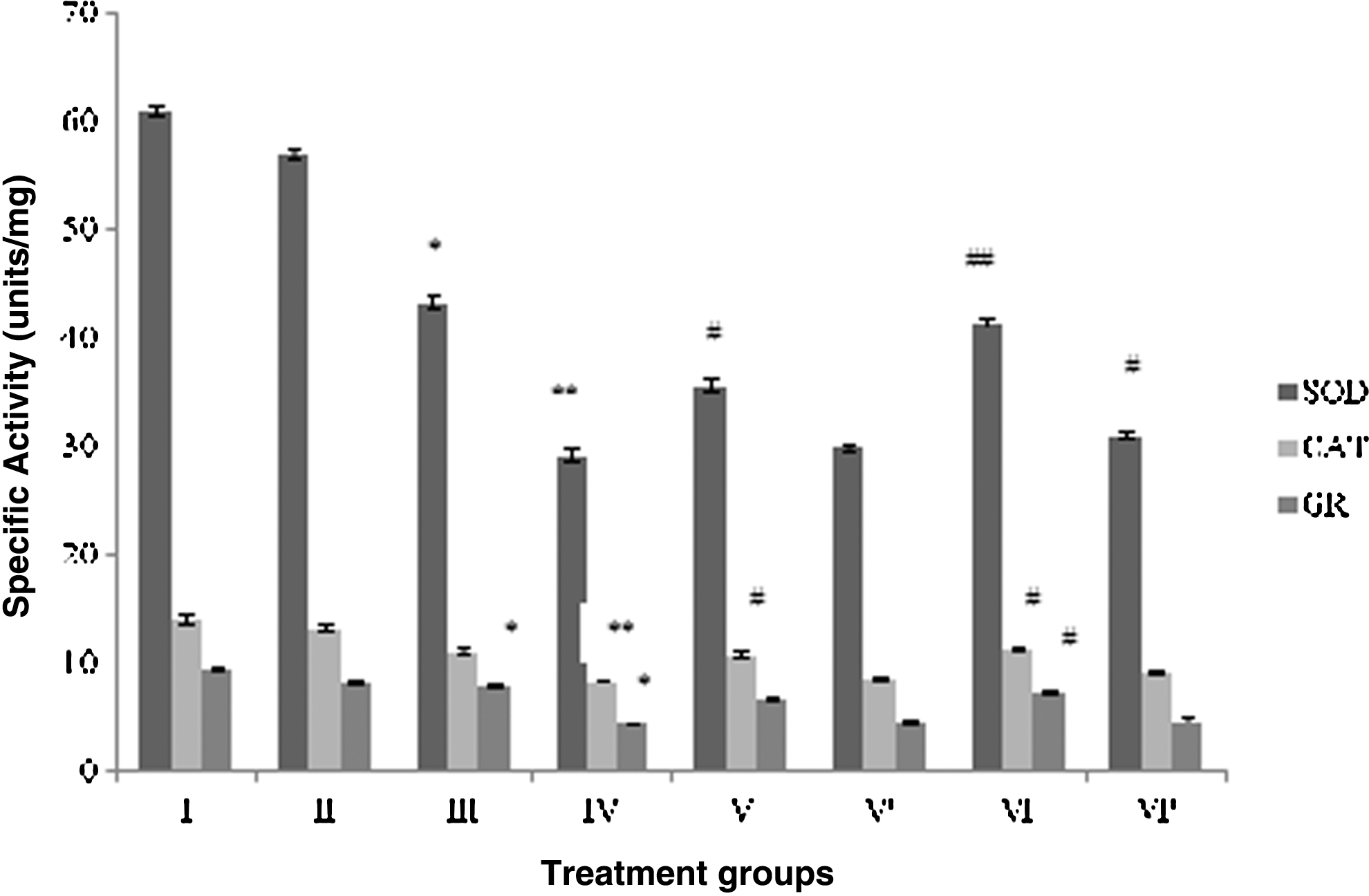
Effect of the treatment on the antioxidant status of brain. Group I: control normal (neither treatment nor light exposure). Group II: control (treated with saline under photoillumination). Group III: riboflavin (2 mg/kg) treated under photoillumination. Group IV: cisplatin (2 mg/kg) treated under photoillumination. Group V: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) under photoillumination. Group V′: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) without photoillumination. Group VI: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) under photoillumination. Group VI′: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) without photoillumination. * indicates significantly different from control at p≤0.05. ** indicates significantly different from control at p≤0.01. # indicates significantly different from group IV at p≤0.05. ## indicates significantly different from group IV at p≤0.01. SOD, superoxide dismutase; CAT, catalase; GR, glutathione reductase.
The combination group V demonstrated enhanced activity by 22.3%, 29%, and 50.7% for SOD, CAT, and GR, respectively, whereas group VI displayed remarkable gain in the activity of the same enzymes by 42.3%, 35.6%, and 68.2%, respectively, as compared to group IV (Fig. 2). Group V′ showed improvement in SOD, CAT, and GR activity by 2.5%, 3.6%, and 4%, respectively, while melioration in the enzymatic activity of group VI′ was found to be 6.7%, 8.8%, and 8.8%.
Effect of the treatment on level of MDA and NO
CP treatment enhanced the MDA and NO levels in the brain samples to a significantly higher level. MDA level was found to be increased by 175.8%, while the NO level was observed to be elevated by 140.4% in group IV with respect to the control. RF-treated group III also exhibited augmentation by 21.5% and 51% in MDA and NO levels, respectively.
Among the combination groups, % decrease in MDA and NO levels was found to be 31.6% and 35.5% in group VI followed by group V, displaying the dip by 12.8% and 25.4%, respectively, in both the parameters when compared to group IV (Fig. 3). Group VI′ also showed decrement in the MDA and NO level by 10.3% and 20%, respectively, while group V′ displayed lowering in the MDA and NO level by 8% and 7.9% (Fig. 3).
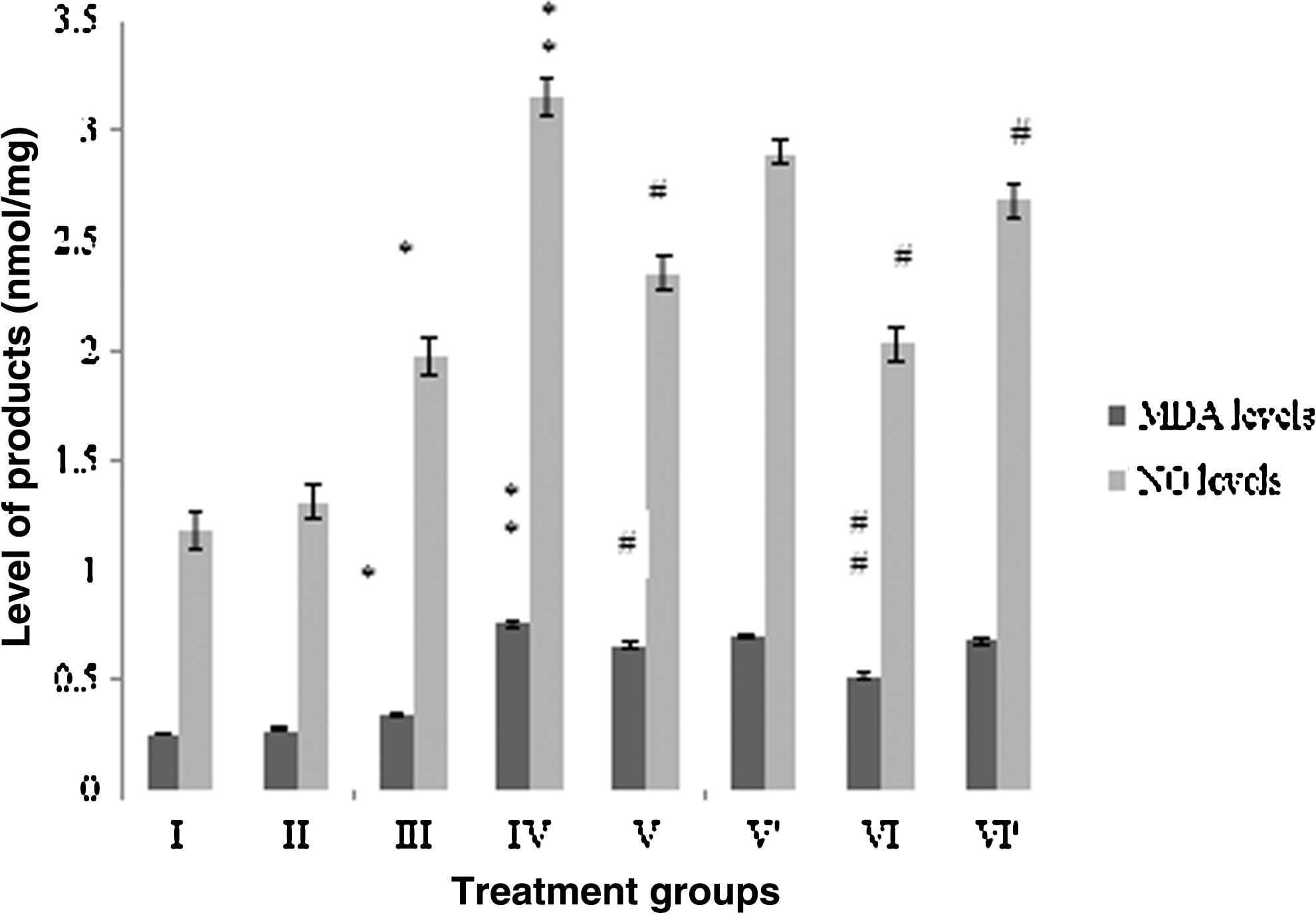
Effect of the treatment on the levels of melondialdehyde (MDA) and nitric oxide (NO) in brain. Group I: control normal (Neither treatment nor light exposure). Group II: control (treated with saline under photoillumination). Group III: riboflavin (2 mg/kg) treated under photoillumination. Group IV: cisplatin (2 mg/kg) treated under photoillumination. Group V′: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) under photoillumination. Group V′: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) without photoillumination. Group VI: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) under photoillumination. Group VI′: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) without photoillumination. * indicates significantly different from control at p≤0.05. ** indicates significantly different from control at p≤0.01. # indicates significantly different from group IV at p≤0.05. ## indicates significantly different from group IV at p≤0.01
Effect of the treatment on GSH and –SH levels
Levels of both the cellular reductants were also decreased significantly in group IV and III after the treatment. The decrease was 47.2% and 40% in group IV in the level of GSH and –SH, while it was 8.3% and 10% in group III, respectively. A pattern of progressive increase in their level was observed in the combination groups, as group V demonstrated the upsurge by 43.4% and 27.2% of GSH and –SH levels, whereas group VI showed the increase of 69% and 38.4% for the both parameters as compared to group IV (Figs. 4 and 5).
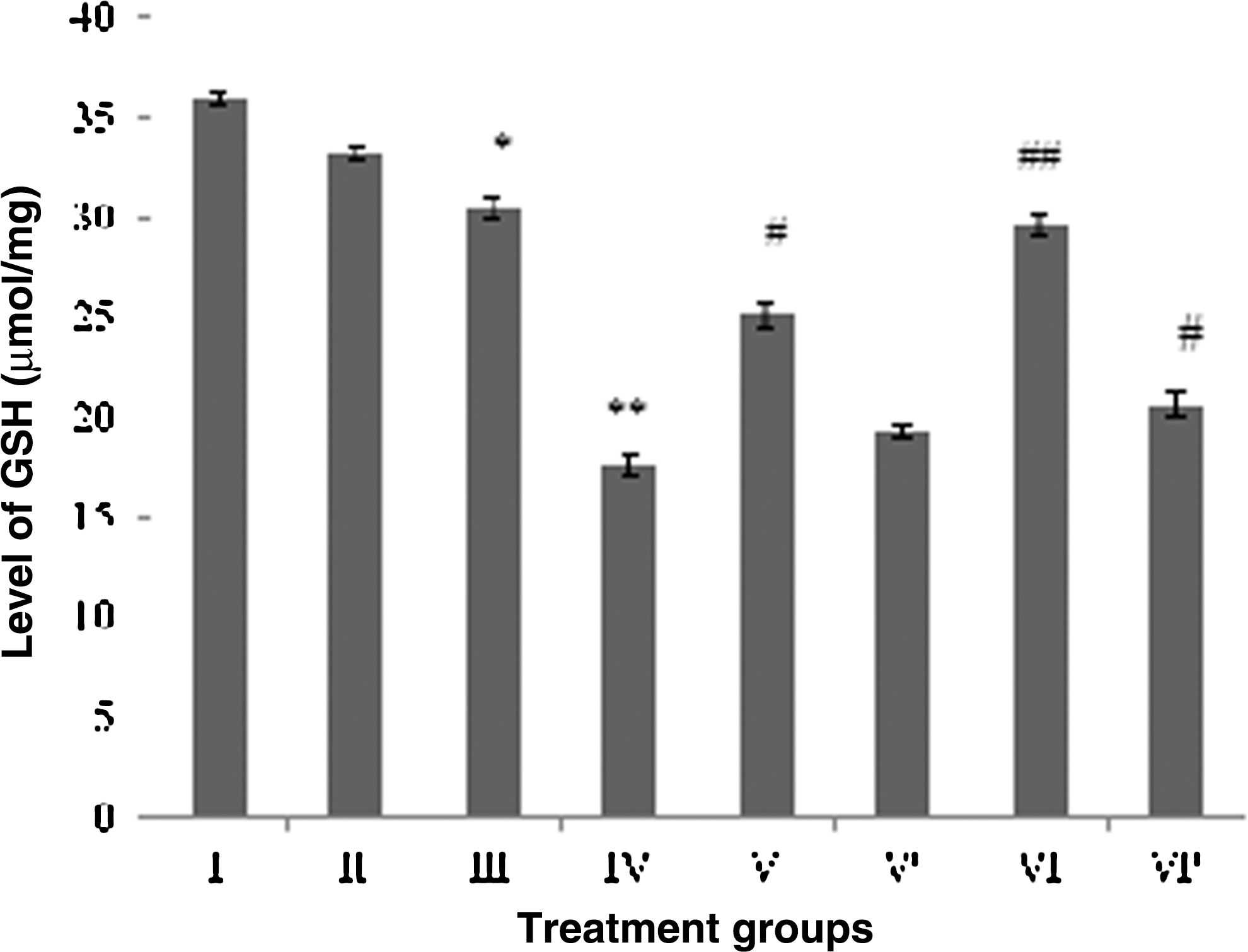
Effect of the treatment on GSH level in brain. Group I: control normal (neither treatment nor light exposure). Group II: control (treated with saline under photoillumination). Group III: riboflavin (2 mg/kg) treated under photoillumination. Group IV: cisplatin (2 mg/kg) treated under photoillumination. Group V: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) under photoillumination. Group V′: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) without photoillumination. Group VI: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) under photoillumination. Group VI′: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) without photoillumination. * indicates significantly different from control at p≤0.05. ** indicates significantly different from control at p≤0.01. # indicates significantly different from group IV at p≤0.05. ## indicates significantly different from group IV at p≤0.01.

Effect of the treatment on the –SH level in brain. Group I: control normal (neither treatment nor light exposure). Group II: control (treated with saline under photoillumination). Group III: riboflavin (2 mg/kg) treated under photoillumination. Group IV: cisplatin (2 mg/kg) treated under photoillumination. Group V: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) under photoillumination. Group V′: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) without photoillumination. Group VI: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) under photoillumination. Group VI′: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) without photoillumination. * indicates significantly different from control at p≤0.05. ** indicates significantly different from control at p≤0.01. # indicates significantly different from group IV at p≤0.05. ## indicates significantly different from group IV at p≤0.01.
The combination-treated groups without light exposure also showed replenishment in the level of both the cellular reductants, although not as significant as their counterparts (group V and VI) under photoillumination. Group V′ depicted enhancement in the GSH and –SH level by 9.6% and 7.2%, whereas group VI′ demonstrated 17.2% and 14.6% of upsurge in the level of GSH and –SH (Figs. 4 and 5).
Effect the treatment on brain function markers
The housekeeping enzymes LDH and Glu-6-PDH were taken as brain function markers, because their activity reflect health status of the brain. Group IV showed a prominent drop in the activity of LDH and Glu-6-PDH by 51.4% and 73.2%, respectively, whereas group III depicted the fall in their activity by 12.2% and 7%, respectively. Among the combination groups, group VI demonstrated remarkable increase in the activity of Glu-6-PDH by staggering 160.4%, while LDH activity was found to be raised by 58.4% followed by group V that showed 34% and 78.8% enhancement in Glu-6-PDH and LDH activity as compared to group IV (Fig. 6).

Effect of the treatment on the brain function markers. Group I: control normal (neither treatment nor light exposure). Group II: control (treated with saline under photoillumination). Group III: riboflavin (2 mg/kg) treated under photoillumination. Group IV: cisplatin (2 mg/kg) treated under photoillumination. Group V: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) under photoillumination. Group V′: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) without photoillumination. Group VI: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) under photoillumination. Group VI′: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) without photoillumination. * indicates significantly different from control at p≤0.05. ** indicates significantly different from control at p≤0.01. # indicates significantly different from group IV at p≤0.05. ## indicates significantly different from group IV at p≤0.01.
Among the combination-treated groups without photoillumination, group VI′ displayed increase in Glu-6-PDH and LDH activity by 8% and 12.3%, followed by group V′ depicting the increment by 4.3% and 6.5% for both markers, respectively (Fig. 6).
Effect of the treatment on GST
A prominent decline in GST activity was observed after treatment with CP and RF as group IV and III showed the dip in their activity by 51.6% and 16%, respectively. Between the combination groups, group VI showed significant rise in its activity by 43%, followed by group V displaying 24% of activity gain as compared to group IV (Fig. 7). The combination groups without photoillumination, group V′ and group VI′, showed 3.6% and 9.3% of improvement in GST activity, respectively (Fig. 7).
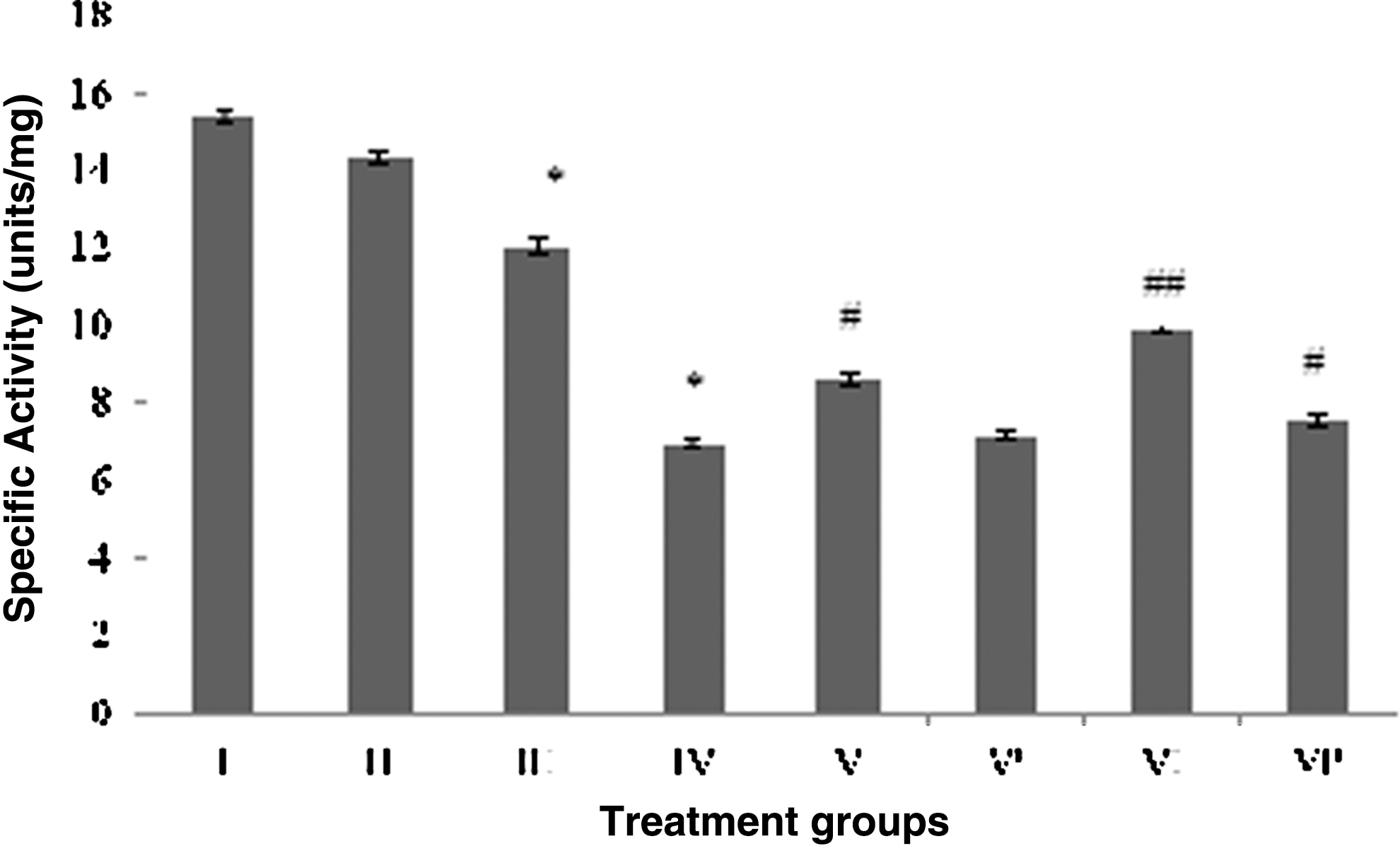
Effect of the treatment on glutathione-S-transferase activity in brain. Group I: control normal (neither treatment nor light exposure). Group II: control (treated with saline under photoillumination). Group III: riboflavin (2 mg/kg) treated under photoillumination. Group IV: cisplatin (2 mg/kg) treated under photoillumination. Group V: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) under photoillumination. Group V′: treated with combination of cisplatin (2 mg/kg) and riboflavin (1 mg/kg) without photoillumination. Group VI: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) under photoillumination. Group VI′: treated with combination of cisplatin (2 mg/kg) and riboflavin (2 mg/kg) without photoillumination. * indicates significantly different from control at p≤0.05. # indicates significantly different from group IV at p≤0.05. ## indicates significantly different from group IV at p≤0.01.
Effect of the treatment on brain histopathology
Representative photomicrographs of the cerebral cortex of the mice (group II to VI) have been depicted in Figure 8 (A to E). The photomicrograph of control group I depicts normal pattern and density of neuropil and neuronal distribution (Fig. 8A), while RF-treated group II exhibits only subtle changes in terms of perineuronal spaces and rarification of neuropil as compared to control (Fig. 8B). CP-treated group III reveals the perceptible structural changes in terms of rarification of neuropil, perineuronal spaces, and dilated vessels (Fig. 8C) besides sloughing of cells indicative of necrotic and apoptotic cell death. Interestingly, combination-I-treated group IV (Fig. 8D) shows features akin to group II, while combination–II-treated group V (Fig. 8E) shows features comparable to the control. Thus, group IV and V displayed dose-dependent recovery in the size of the cells as well as disappearance of sloughing (Fig. 8D, E) that entails RF to have an ameliorative effect for CP-induced neuronal damage.

Histopathological examination of the cerebral cortex of the brain of the treated mice. Photomicrographs showing part of the cerebral cortex: superficial cell-sparse molecular layer (striped blue arrow) and deeper cell-rich external granular (notched arrow) layer of mice cortical gray matter. Control group
Discussion
CP-induced neurotoxicity is one of the major side effects that poses a serious medical challenge of paramount concern forcing its discontinuation during long-term treatment. 2 Lipid- and protein-rich brain tissue dependent on oxygen-rich environment to function properly is at a greater risk of oxidative and nitrosative damage because of poor shield of antioxidant enzymes and low regeneration power as compared to other tissues in our body. 25 Neurons, after the damage, cannot be healed properly, as they depend totally on the stem cells for their maintenance and limited regeneration. Thus, amelioration of neurotoxicity is an important clinical issue for better life of the concerned patients, as many neuronal damages are irreversible in nature. Different strategies have been used since the therapeutic usage of the drug to counter this side effect, but an effective strategy is yet to be acknowledged. Recently, we have demonstrated that photoirradiated RF can ameliorate CP-elicited toxicity in situ as well as in vivo. 13 –15 The present work is an attempt to assess the ameliorative potential of this combination on CP-induced neurotoxicity in a mouse model.
In the present study, the body weight of the CP-treated group was markedly decreased (data not shown). It is well known that regular dosing of CP decreases the appetite of the treated animals besides accelerating their catabolic processes, leading to a prominent decline in the body weight. The systemic toxicity after multiple doses of CP may also affect the brain; so, brain weights of this group were also found to be moderately decreased. Their biochemical analysis entailed that CP was a strong neurotoxic agent as evidenced by elevated oxidative and nitrosative stress in brain tissue. Treatment with RF also exerted similar neurotoxic insults in the treated group, but the extent of damage was less than the CP-treated group. Moreover, the histopathological examination of brain of the CP-treated group confirmed significant tissue damage in the cortex region of brain. These findings are in accordance with earlier studies, 26 –28 in which they have reported that the accumulation of CP in various regions of brain might be due to its possible infringe into the brain, including the brain cortex, via free radical-mediated deterioration of the blood–brain barrier (BBB). Interestingly, the combination of RF and CP under photoillumination resulted into alleviation of the neurotoxicity indicated by normalization of various parameters taken under the study in a dose-dependent manner. The parallel groups treated with the CP-RF combination without photoillumination showed mild improvement in the same parameters with respect to their counterparts.
The most classical, yet one of the strongest, explanations for this drug-induced toxicity is the elevated oxidative and the nitrosative stresses. 25,29,30 Furthermore, it is established that a mild level of NO helps to maintains normal synaptic transmission and memory, but its elevated level leads to neurotoxicity by flipping its action from a secondary messenger to an invasive radical. 31 In the present study, high doses of CP might have also increased NO level to such extent resulting into neurotoxicity in the treated animals. Moreover, they can also exacerbate the prevailing pathological condition by coupling with oxidative stress to the worse. 32,33
It is also possible that ROS and RNS generated near the BBB can cause local inflammation inducing phagocytes to express iNOS via activation of proinflammatory cytokines. 26,27,33,34 Literature suggests that CP probably binds to the serum proteins, thereby providing a continuous contact to the BBB. This makes the neural cells occurring beyond the barrier vulnerable to the drug-induced toxicity. Hence, DRG cells, being not guarded by the BBB, are its major target cells followed by peripheral nerves, dorsal, and ventral nerve roots, while the spinal cord and frontal lobe are least affected, as they are guarded by BBB. 34 Histopathological and morphological analysis of nerve cells has revealed that the platinum component of the drug binds to DRG cells, leading to decrease in size of the nucleus, nucleolus, and somata of the neuronal cells. 29,35,36 It is thus possible that repeated doses of the drug can generate abundant free radicals in the blood of nearby region and also around the BBB. If these radicals are not countered effectively, their constant invasion on the BBB can facilitate the drug to infringe the brain tissues resulting into serious neuropathological conditions, including CP-induced neurotoxicity. 27,33 The histopathological examination under the study pleads this notion as CP-treated brain cortex section showed necrotic and apoptotic features such as sloughing as well as shrinkage of ganglion cells and their nucleus. Recent studies suggest that CP can also interfere with axonal mitochondrial membrane transport that can cause cytoskeleton dysfunction and derangement in the target cells inducing autophagy of mitochondria. 37 Moreover, such mitochondrial damages can enhance oxidative stress by spilling out the free radicals, thereby exhausting the cellular reductant-generating enzymes such as Glu-6-PDH and LDH, resulting into decline in their activity in the target cells. Upon accumulation, all these damages can trigger apoptosis as their final effect by release of cytochrome C and activation of the caspase pathway in the cells.
As an effective photosensitizer, RF has a great potential to be used in photodynamic therapy and ribophototherapy for treatment of various diseases, including cancer. 10,11,38 In addition, De Souza et al. (2006) and De Souza et al. (2007) in separate experiments have demonstrated that the photoproducts of RF can play as adjuvants for apoptosis induction in prostrate (PC3) and leukemia cancer cell lines under radiochemotherapy. Furthermore, RF, being an excellent photosensitizer, can also induce apoptosis via Fas–Fas ligand-mediated pathway involving caspase activation in the cells under high level of the stresses like those in the cancer cells. 10,11,38 These studies propelled us to design the present investigation in which we have used RF for alleviation of CP-induced neurotoxicity. Intriguingly, we observed that the mice treated with the combination of CP and RF under photoillumination showed significant recovery from the CP-induced neurotoxicity with respect to the CP-treated group. The extent of normalization of the parameters under the current study in the combination-treated groups with respect to the CP-treated group was found to be dependent on RF dose. This interesting effect exhibited by CP-RF combinations is in accordance with our previous findings in situ and in vivo. 13 –15
CP is a proved neurotoxic agent under many previous investigations. A study conducted on a human neuroblastoma cell line –SH5Y indicated that Bcl-2 phosphorylation was not involved in the action of CP, but its level was found significantly decreased. 36 The downregulation of this protein has been observed in many neuronal and non-neuronal models of apoptosis. The study also ruled out the possibility of its proteolytic cleavage involving activation of caspase 3/7. Moreover, involvement of p53 as a negative transcription factor for the Bcl-2 gene was anticipated in a similar study. 39 A great deal of literature suggests that high dose of CP activates the MAPK family that activates and stabilizes p53 by phosphorylation with concomitant assistance of other members such as the p38, JNK/SAPK and ERK pathways in parallel. They are responsive to the external stressful conditions for the cells, and once activated, they can act as proapoptotic factors. 40 Other members of MAPKs might have also been activated during exposure of cells to CP, hence possibility of their involvement in p53 activation and stabilization, as the later stage cannot be ruled out in the prevailing condition. Over all, these events consequently lead to downregulation of Bcl-2 expression. 39,41 Thus, the reduced amount of Bcl-2 is possibly the result of lowered expression rather than its degradation without involvement of caspase 3/7. 36
The explanation of CP-induced chronic neurotoxicity at cellular level is not possible solely on the basis of DNA damage rather it also involves the activation of complex array of various molecular pathways leading to apoptosis induction in the cells. Thus, CP activates particular members of MAPKs involved, inter-alia, in apoptosis-mediated cell death. At the same time, there is a quantitative reduction in Bcl-2, p53, and MAPK proteins as late event. 36 This effect of CP alone could possibly be dependent on DNA damage and consequent decrease in RNA synthesis also. Thus, DNA damage may induce an indirect and late interference with the availability and activation of specific molecular pathways regulating the equilibrium between apoptosis and cell cycle. In our study, CP when given with RF, all late events leading to DNA damage might not occur, as both the compounds interact mutually. 15 It might push the equilibrium toward apoptosis. Furthermore, as the CP-RF combination is less DNA damaging than CP alone, apoptosis is favored instead of necrosis as the final consequence. This notion has been found true in preliminary studies conducted by our laboratory on the BC-3 cell line where apoptosis-inducing capacity of CP was found to be enhanced by 60% when given with RF 15 (data not published).
It is well established now that CP at higher dose induces cell death by necrosis instead of apoptosis depending on the redox status and ATP level. 15 In the present investigation, we suggest that RF when present with CP interacts in such a way that lowers oxidative and nitrosative stress to an extent that favors apoptosis. As CP exerts its side effects mostly by elevation in oxidative and nitrosative stress, hence the interaction between active constituents of RF and CP may be the putative reason for diminished neurotoxicity in the combination-treated groups in the present investigation. This may occur even in ambient light or without illumination, but the effect is stronger in presence of light because of increased propensity of RF under photoillumination. 15 Hence, in the light of RF playing a pivotal role in cancer etiology, tumerogenesis, as well as efficacy of anticancer drugs, 10,12,38 the proposed RF-CP combination thus has great potential to be used as more effective and efficient treatment strategy for various cancers instead of CP alone under chemotherapy and chemoradiotherapy. 42,43
Conclusion
We hypothesize that continuous production of CP-induced free radicals if somehow is sustained for longer duration in blood and around the BBB can damage the surrounding tissues and even the barrier. In such condition, CP can probably infringe into the barrier, making the brain vulnerable to its toxic insults via membrane damage. In RF-CP combination, RF may counter the CP-induced oxidative and nitrosative stresses through their mutual interaction at the molecular level leading to decreased CP-induced neurotoxicity. 15
Finally, our hypothesis strongly supports the involvement of oxidative and nitrosative stresses in CP-induced neurotoxicity in vivo. However, further study is required to assess clinical feasibility of CP-RF combination under chemoradiotherapy.
Footnotes
Acknowledgments
We acknowledge the financial favor provided by the University Grant Commission (UGC), New Delhi, under SAP-II program, DST- FIST, and the facilities provided by Department of Biochemistry, Aligarh Muslim University. We are grateful to Dr. Eram Husain, Dr. Shams Tabrez, and Dr. Ghulam Md. Ashraf for their cooperation and motivation. We are also thankful to Syed Hassan Mehdi, Dilnasheen Meerza, Maria Salman, and all the friends who directly or indirectly helped us to produce this work.
Disclosure Statement
No competing financial interests exist.