
Abstract
Lung cancer is the leading cause of death from malignancy in people and over 85% of these patients eventually die from disseminated disease. Paclitaxel (TAX) is widely used as an antimicrotubule agent for the treatment of lung cancer. Unfortunately, the resistance to this antimicrotubule agent occurs frequently. Stathmin (STMN1) is a ubiquitous microtubule destabilizing protein linked to cancer and cell health and its expression level often correlates with cancer stage progression and prognosis for survival. Overexpression of the antiapoptotic protein Bcl-2 has been shown to prolong drug-induced growth arrest, potentially inducing resistance. In this study, we used a short hairpin RNA (shRNA) approach to evaluate the effect of STMN1 and Bcl-2 downregulation in the sensitivity to TAX in lung cancer cells. We achieved significant downregulation of STMN1 and Bcl-2 mRNA and protein expression by a combination of double shRNA treatment strategy. Our experimental data showed that inhibition of STMN1 and Bcl-2 expression with RNA interference can sensitize lung cancer cells to TAX. These findings suggest a novel approach to improve the efficacy of certain antimicrotubule agents against lung cancer by regulating the function of STMN1 and Bcl-2.
Introduction
Lung cancer is the leading cause of death from malignancy in both males and females and over 85% of these patients eventually die from disseminated disease, accounting for 29% of all cancer deaths, more than prostate, colorectal, and breast cancers combined. 1,2 Sixty percent of patients are considered inoperable at the time of diagnosis and 20% are poor surgical candidates. 3 The overall 5-year survival rate of 10%–13% has not changed appreciably over the last two decades.
Paclitaxel (TAX) is widely used as an antimicrotubule agent for the treatment of lung cancer. 4 It stabilizes microtubules and inhibits depolymerization back to tubulin, resulting in mitotic inhibition. Such an effect causes cell cycle arrest in G2/M phase and induces apoptosis. 5 Unfortunately, the resistance to this antimicrotubule agent occurs frequently. Nonspecific resistance mechanisms of include those mediated by the overexpression of drug efflux pumps, whereas mechanisms specific to antimicrotubule agents include alterations in tubulin through mutation, differential expression of tubulin isotypes, post-translational tubulin modifications, and altered expression of microtubule regulatory proteins. 6 One such microtubule regulator is stathmin (STMN1), an 18-kDa protein that destabilizes microtubules through catastrophe promotion and/or tubulin sequestration. 7 STMN1 has been shown to regulate the dynamics of the microtubules that make up the mitotic spindle, a role associated with carcinogenesis that indicates that STMN1 is one of the fundamental cancer-associated genes and a potential target for diagnosis and treatment. 8 Preliminary studies have suggested that an increased expression of STMN1 can affect the stability of the microtubular system and may affect the ability of TAX to bind to the microtubular system and is associated with resistance to TAX. 9
Decreased apoptosis contributes to cancers. 10 The Bcl-2 family includes both proapoptotic (Bax, Bak) and antiapoptotic (Bcl-2, Bcl-xL) proteins, and plays a crucial role in mitochondria-driven cell death. 11,12 Overexpression of the antiapoptotic protein Bcl-2 prolongs drug-induced growth arrest, potentially inducing chemotherapeutic resistance. 13 Bcl-2 is an appealing drug target as it can be used to increase cell death in cancer.
RNA interference (RNAi) technology can downregulate targeted genes and has been evaluated as a potential therapeutic strategy in human cancer therapy. 14 It involves post-transcriptional gene silencing via a process in which double-stranded RNA inhibits gene expression in a sequence-dependent manner through degradation of the corresponding mRNA. 15 Its blocking action on gene expression has been successfully observed in rat and human cells cultured in vitro, and the knockdown of genes in cells has been achieved. Inhibition of several targets using short hairpin RNAs (shRNAs) has been reported to suppress growth of various cancer cell lines, and offers an important nucleotide-based approach at suppressing gene products for loss-of-function analyses. 16
Based on these studies, we designed two plasmids encoding shRNA against STMN1 and Bcl-2 and investigated a combination of double shRNA interferences efficacy in regard to overcoming TAX resistance in lung cancer cells and suppression of STMN1 and Bcl-2 expressions in vitro. Our findings suggest that STMN1 and Bcl-2 may be two important target proteins involved in lung cancer chemosensitivity. An approach to inhibit both STMN1 and Bcl-2 may help to augment the cytotoxic effect of TAX.
Materials and Methods
Cell culture and transfection
Cells from the lung cancer cell line A549 and the TAX-resistant derivative A549/TAX were gifts from Cell Center Shanghai Fudan University. They were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco BRL) containing 100 μg/mL penicillin and 100 μg/mL streptomycin and supplemented with 10% calf blood serum (SijiQing Laboratories) at 37°C in a humidified atmosphere with 5% CO2. 0.05 mg/L TAX (Hubei Willman Pharm.) was consistently added to the medium of the A549/TAX to maintain the TAX resistance. Cells were grown at 37°C/5% CO2 in DMEM containing 10% fetal calf serum (FCS), 4500 mg/L glucose and nonessential amino acids (Gibco BRL). Transfection studies were carried out with 6×105 cells/mL plated on a six-well plate. Two point five μg DNA was mixed with 5 μL Lipofectamin (Invitrogen) and 250 μL DMEM and incubated for 5 minutes at RT. For transfection, cells were washed with phosphate-buffered saline (PBS) and incubated for 5 hours in starving medium with the related plasmid. After 5 hours, medium DMEM with 20% FCS was added to the cells. Forty-eight (48) hours after transfection, cells were washed with PBS and fixed with methanol for 3 minutes, washed with PBS and dried. Cells were analyzed for GFP expression and transfection efficiency with fluorescent microscope and flow cytomitry.
Plasmids and RNAi
Different shRNAs targeting to the genes of STMN1 and Bcl-2 were designed and constructed into pPGU6-GFP-plasmid (Shanghai Gene Pharma, Inc.). Eight siRNA sequences from various coding regions of STMN1 gene and Bcl-2 gene were selected as the following: STMN1-294, 5-GAGGAAATTCAGAAGAAAT-3 (from 294 bp), named as S1; STMN1-392, 5-GGCTGAGAAACGAGAGCACGA-3 (from 392 bp), named as S2; STMN1-428, 5-GAAGGCAATAGAAGAGAACAA-3 (from 428 bp), named as S3; STMN1-530, 5-GGATAAGCACATTGAAGAA-3 (from 530 bp), named as S4. Bcl-2-506, 5-GGGAGAACAGGGT ACGA T AAC-3 (from 506 bp), named as B1; Bcl-2-928, 5-GAGGA TTGTGGCCTTCTTTGA-3 (from 928 bp), named as B2; Bcl-2-1048, 5-GCACACCTGGA TCCAGGAT AA-3 (from 1048 bp), named as B3; Bcl-2-1107, 5-GCA TGCGGCCTCTGTTTGA TT-3 (from 1107 bp), named as B4. Then shRNA plasmids named pPGU6-GFP-STMN1 and pPGU6-GFP-Bcl-2 were identify using enzyme digestion and further confirmed by direct sequencing (Invitrogen) according to the manufacturer's protocol.
RNA extraction and quantitative polymerase chain reaction analysis
Total RNA was isolated from organs or cells using the TRIzol reagent according to the manufacturer's instructions (Invitrogen). RNA reverse transcription and polymerase chain reaction (PCR) amplifications were performed as described. Primers used in these studies are indicated here: GAPDH (GCACCGTCAAGGCTGAGAAC, TGGTGAAGACGCCAGTGGA, 138 bp in total); STMN1 (ACAAATGGCTGCCAAACTGGA, CTCGTCAGCAGGGTCTTTGGA, 94 bp in total); Bcl-2 (TCAGCA TGGCTCAAAGTGCAG, GAAACAGA TGTCCCTACCAACCAGA, 151 bp in total). All those primers were from Invitrogen.
Protein extraction and western blot analysis
The Monoclonal antibodies, STMN1 and Bcl-2 were purchased from Abcam Biotechnology, Inc. and Cell signaling, Inc., respectively. Monoclonal antibody of β-action was purchased from Santa Cruze Biotechnology, Inc. Protein lysates were separated by 10% SDS-PAGE, transferred onto PVDF membrane (Hybond-P; GE Healthcare), probed with specific antibodies (1:1000 for β-action, 1:50,000 for STMN1, 1:1000 for Bcl-2) overnight at 4°C, and revealed using the AP–conjugated antimouse or antirabbit secondary antibody (1:500) from Beijing golden Bridge Inc. They were then developed with an enhanced chemiluminescence Western blot detection kit (Amersham ECL plus Western blotting Detection System; GE Healthcare).
Apoptotic cell staining
A549 and A549/TAX cells were seeded in flat-bottomed six-well plates and were treated with recombinant shRNA plasmids. Untreated cells served as controls. After 48 hours of treatment, cells were incubated with Hoechst33342 and propidium iodide (PI; Nanjing Keygen Biotech) for 10 minutes, washed with PBS twice and observed under a fluorescence microscope.
Proliferation analysis
Proliferation was measured by using the Cell Counting Assay Kit-8 (Dojindo Molecular Technologies) according to the manufacturer's protocol. One hundred microliters of A549 or A549/Tax cells were plated on 96-well plates at a density of 1.5×104 cells per milliliter and cultured for 24 hours. Cells were then transfected with recombinant shRNA plasmids. Twenty-four hours later, they were exposed to different concentrations of TAX (0.3, 3.0, 30.0 and 300.0 mg/L) for 48 hours. Ten microliters of Cell Counting Assay Kit-8 solution was added to each well, the cells were incubated for another 2 hours, and the absorbance at 450 nm was measured by using a microplate reader (BioTek Instruments). The amount of the formazan dye, generated by the activities of dehydrogenases in cells, is directly proportional to the number of living cells. Inhibitory rate of cellular growth was calculated as the following formula: Inhibitory rate (%)=(1−A value in experimental group/A value in control group)×100%. The 0 μM group was used as black control group. The IC50 value (the concentration of the drug which is capable of bringing about 50% inhibition of cell survival) of the drug used for treatment was determined by plotting a graph with inhibitory rate of cell growth (Y-axis) against the concentrations of the TAX (X-axis).
Evaluation of apoptosis
Apoptosis was determined by staining cells with Annexin V-FITC and PI using Annexin V-FITC apoptosis detection kit (BD Pharmingen). A549 cells were treated with recombinant shRNA plasmids for 96 hours, and then harvested, washed twice with cold PBS. Untreated cells served as controls. The prepared cells were resuspended in binding buffer (10 mM HEPES/NaOH [pH 7.4], 140 mM NaCl, 2.5 mM CaCl2) at a concentration of 1×106 cells/mL. Then, 5 mL each of Annexin V-FITC (Pharmingen) and PI were added to these cells, which were analyzed by FACStar flow cytometer.
Statistics
Data were expressed as the mean±SD, and were analyzed using independent samples t-test and variance (ANOVA) as appropriate by statistical software (SPSS Base 13.0 for Windows; SPSS, Inc.). p-Values<0.05 were considered significant different.
Results
Inhibition of STMN1 expression by plasmid encoding a STMN1 shRNA
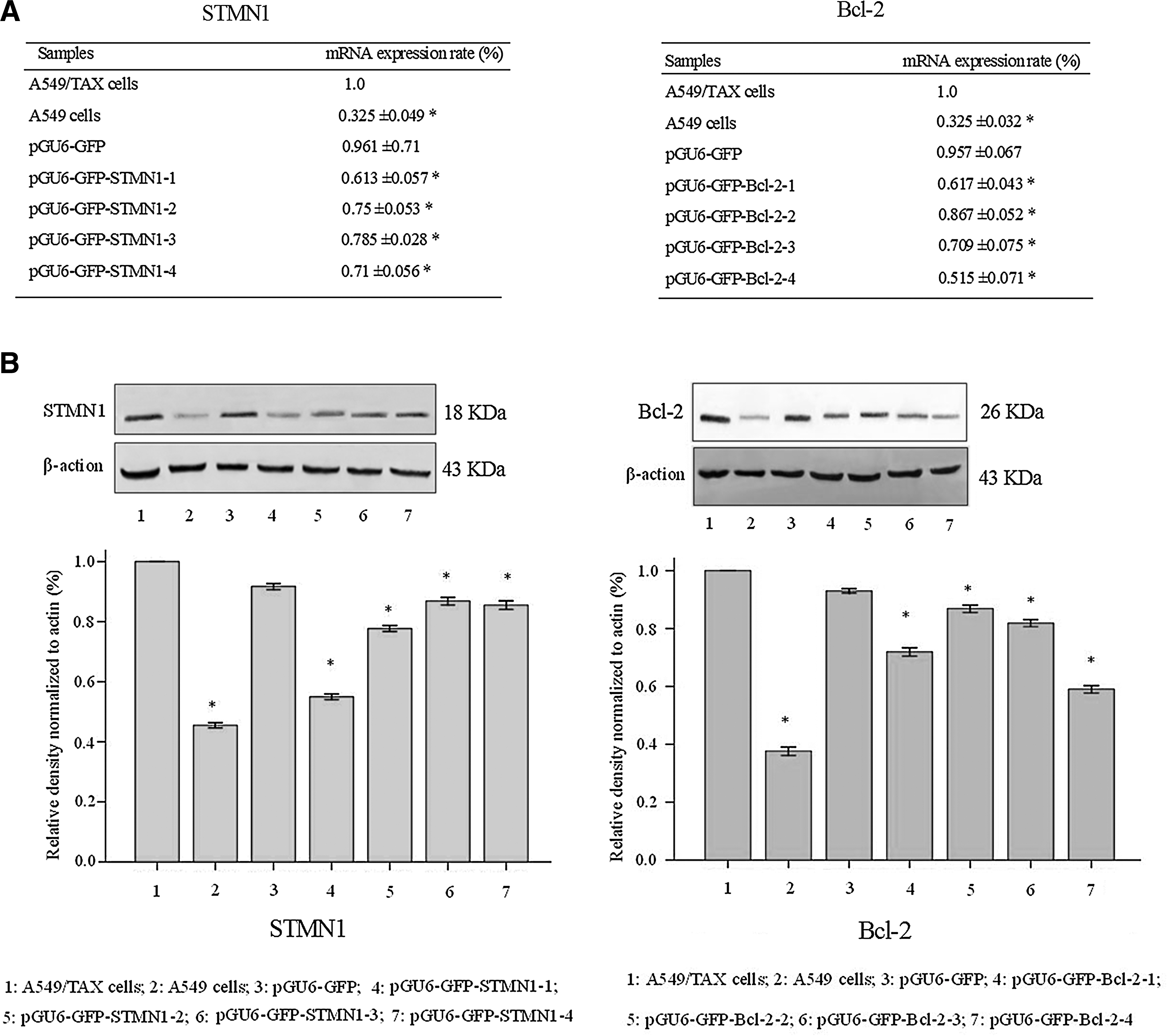
Four plasmid vectors encoding shRNAs against different target sites within the STMN1 transcript, pGU6-GFP-STMN1-1 (S1), pGU6-GFP-STMN1-2 (S2), pGU6-GFP-STMN1-3 (S3), and pGU6-GFP-STMN1-4 (S4) (Fig. 1), were constructed. To verify the effective target site in the STMN1 coding region, four different plasmid vectors targeting STMN1 were transfected into A549/TAX cells. Reduction of STMN1 mRNA expression was monitored by real-time quantitative reverse transcription-PCR and western blot analysis, respectively. As shown in Figure 2, S4 caused more decrease of mRNA and protein level of STMN1 than the other three plasmid vectors. Thus, the shRNA synthesized by S4 was used in the silencing system.

The schematic diagram of the shRNA plasmids pGU6-GFP-STMN1 and pGU6-GFP-Bcl-2.
The expression of STMN1 and Bcl-2 in lung cancer cells was assessed by real-time RT-PCR analysis and western blot analysis.
Inhibition of Bcl-2 expression by plasmid encoding a Bcl-2 shRNA
Four plasmid vectors encoding shRNAs against different target sites within the Bcl-2 transcript, pGU6-GFP-Bcl-2-1(B1), pGU6-GFP-Bcl-2-2 (B2), pGU6-GFP-Bcl-2-3 (B3), and pGU6-GFP-Bcl-2-4 (B4) (Fig. 1), were constructed. To verify the effective target site in the Bcl-2 coding region, four different plasmid vectors targeting Bcl-2 were transfected into A549/TAX cells. As shown in Figure 2, B1 treatment decreased Bcl-2 mRNA and protein expression compared with the other three plasmid vectors treatment in the real time RT-PCR assay and western blot analysis. So in the following we chose B1 in the study of reversing TAX resistance.
Inhibition of STMN1 and Bcl-2 expression by a combination of double shRNA interferences
To investigate whether a combination of double shRNA interferences can effectively inhibit STMN1 and Bcl-2 expression, A549/TAX cells were treated with S1+B4, S1, B4, and pGU6-GFP, respectively. As shown in Figure 3, a combination of double shRNA interferences targeting STMN1 and Bcl-2 treatment strategy decreased STMN1 and Bcl-2 mRNA and protein expression in the real time RT-PCR assay and western blot analysis, but the control plasmid had little effect on STMN1 and Bcl-2 expression.

Inhibition of STMN1 and Bcl-2 expression by a combination of double shRNA interferences was assessed by real-time RT-PCR analysis and western blot analysis. A549/TAX cells were treated with S1+B4, S1, B4, and pGU6-GFP, respectively.
Effect of a combination of double shRNA interferences on inhibition of A549/TAX cell growth
To investigate whether a combination of double shRNA interferences can reverse TAX resistance in lung cancer cells, A549/TAX cells were treated with B4+S1, B4, S1, and pGU6-GFP, respectively. Twenty-four hours later they were exposed to different concentrations of TAX for 48 hours.
As shown in Figure 4, the different treatment schemes induced inhibition of growth in A549/TAX cells. Under the concentrations of 0.3 mg/L of TAX, the rank of inhibition potency was, A549 (IC50: 0.26±0.018)>A549/TAX cotransfected with B4 and S1 (IC50: 0.57±0.052)>A549/TAX transfected with S1 (IC50: 2.37±0.087)>A549/TAX transfected with B4 (IC50:2.63±0.396)>A549/TAX transfected with empty plasmid (IC50:10.93±0.44)>A549/TAX (IC50: 11.19±0.42). The relative reverse rate to TAX was 78.28%, 80.71%, and 97.17% for the group of B4, S1, and B4+S1 group, respectively (Fig. 4). Cotransfection with interfere RNA of Bcl-2 and STMN1 reversed the TAX resistance.
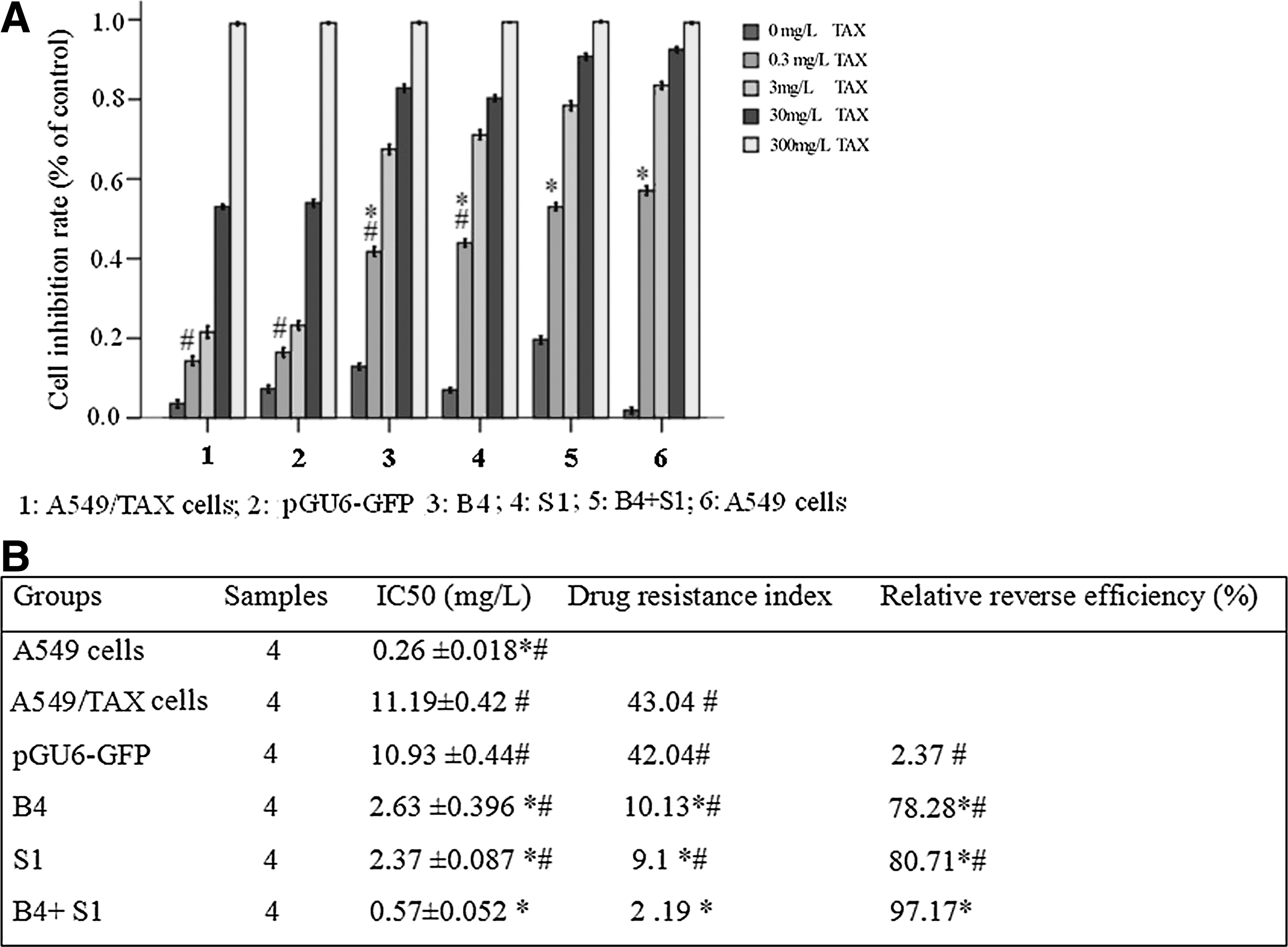
Effect of treatment with a combination of double shRNA interferences on inhibition of A549/TAX cell growth.
A combination of double shRNA interferences contribute to TAX-induced apoptosis
To test whether this enhanced cytotoxicity caused by combined double shRNA interferences and low dose TAX treatment was due to increased apoptosis, we examined the apoptotic changes in A549/TAX cells after different treatment schemes. A549/TAX cells were treated for 96 hours with 0.3 mg/L TAX, B4, S1, B4+0.3 mg/L TAX, S1+0.3 mg/L TAX, and B4+S1+0.3 mg/L TAX, respectively. Enhanced permeability of cellular membrane, an early sign of apoptosis was assessed by FACS analysis using Annexin V-FITC staining. 0.3 mg/L TAX treatment alone only caused membrane instability in a small number of cells with ratios slightly above the basal level in A549/TAX cells, whereas B4+0.3 mg/L TAX and S1+0.3 mg/L TAX treatments caused 36.69% (*p<0.05) and 33.93% (*p<0.05) of A549/TAX cells to lose membrane integrity. The combined treatment of 0.3 mg/L TAX with B4 with S1 resulted in significant 41.10% (*p<0.05) of A549/TAX cells (Fig. 5). The asterisk “*” designates p<0.05 versus A549/TAX cells treated groups (n=6). In line with the results from FACS analysis, the majority of A549/TAX cells displayed typical apoptotic changes, such as DNA fragmentation and chromatin condensation after transfected with a combination of recombinant shRNA plasmid as illustrated by Hochest 33342 and PI staining (Fig. 6). These results suggest that double shRNA interferences treatment induce more lung cancer cells to undergo apoptosis.

Induced apoptotic changes examined by Annexin V-FITC staining. A549/TAX cells were treated for 96 hours with 0.3 mg/L TAX, B4, S1, B4+0.3 mg/L TAX, S1+0.3 mg/L TAX, and B4+S1+0.3 mg/L TAX, respectively. After 96 hours, cells were stained with FITC-labeled Annexin V and immediately analyzed by flow cytometry. Data are presented as mean±SD of the percentage of annexin positive cells from three independent experiments.
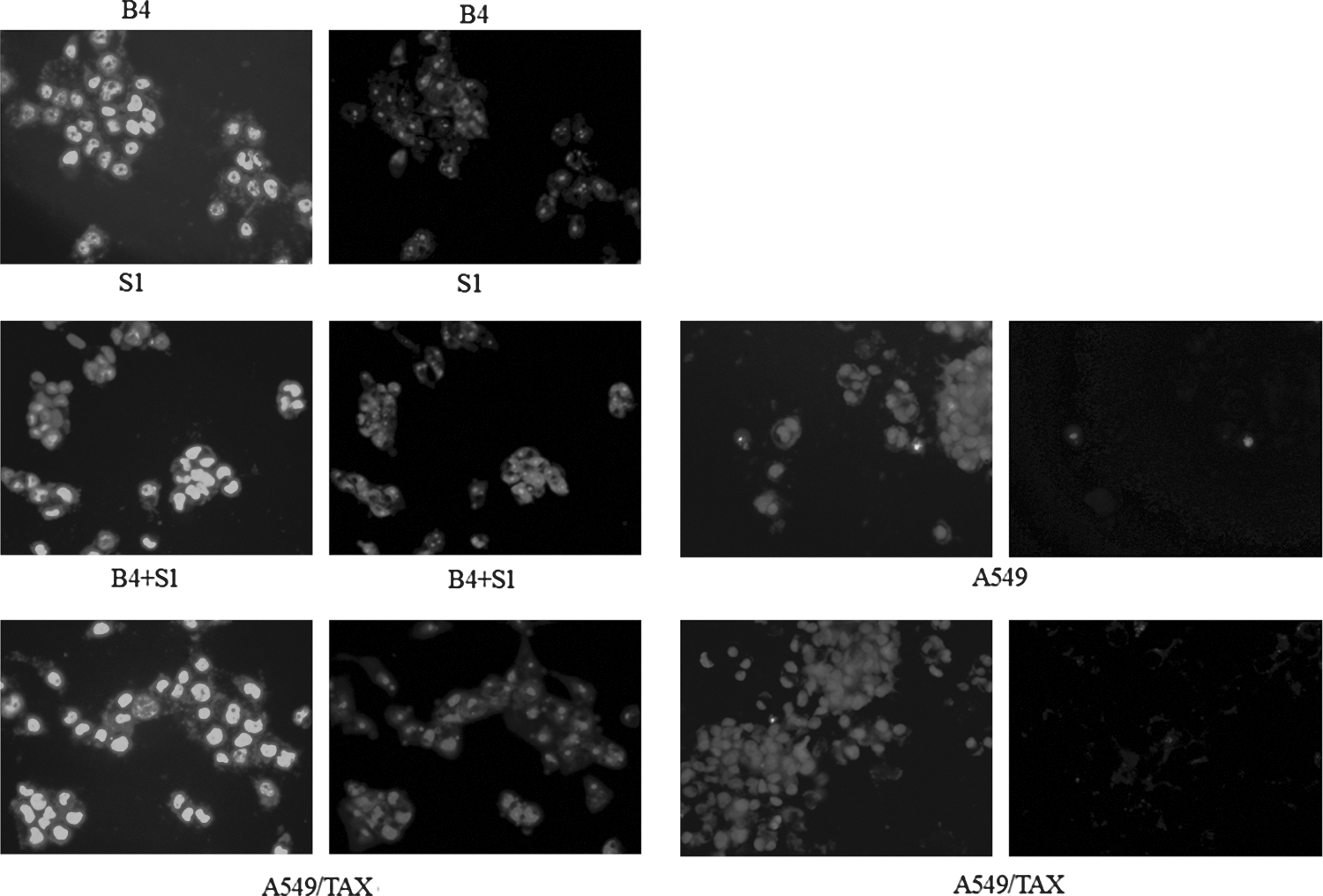
Morphological changes of the nuclei examined by Hoechst 33342 and PI staining. A549/TAX cells were treated with B4+S1, B4, S1, respectively. After 48 hours, A549/TAX cells were incubated with Hoechst 33342 and PI for 2 hours, and the nuclei condensation and fragmentation were observed under a fluorescence microscope. Original magnification, 400×. PI, propidium iodide.
Discussion
Lung cancer is a leading cause of cancer death worldwide. Lung cancers are classified into small cell lung cancer (SCLC) and nonsmall cell lung cancer (NSCLC). 17 Approximately 85% of lung cancers are NSCLC and the majority of patients are diagnosed at an advanced stage. The prognosis in NSCLC is poor, and half of patients present with stage IIIA or stage IIIB disease. Despite significant advances in oncology over the last several decades, lung cancer remains highly lethal. The limited efficacy of cytotoxic chemotherapy and radiotherapy and anticancer drug resistance remain as a major obstacle for the treatment of patients with advanced lung cancer. 3,18
Antimicrotubule agents are commonly used chemotherapy drugs for the treatment of lung cancer. 19 STMN1 is a ubiquitous microtubule destabilizing protein linked to cancer cell health. Its expression level often correlates with cancer stage progression and prognosis for survival. It has been called a cell survival factor because its level of over expression correlates with cancer stage progression, invasion, and metastasis for many cancer types. 20,21 Bcl-2 is an antiapoptosis factor and it is also thought to be involved in resistance to conventional cancer treatment. It is the first oncogene that functions through production of an inhibitor of programmed cell death. Overexpression of Bcl-2 is common in many human cancers, and contributes to increased resistance to chemotherapy. 13
RNAi is a post-transcriptional gene silencing approach first discovered in the nematode Caenorhabditis elegans. 22 Because of its high specificity and efficiency in down regulating gene expression, it has now become an excellent tool for exploring gene function. Many groups have worked on cancer gene silencing using RNAi in cell lines derived from different tissues, which lead to significant inhibition in cancer cell growth. 23,24
This study demonstrated that a combination of double shRNA interferences targeting Bcl-2 and STMN1 treatment strategy significantly decreased Bcl-2 and STMN1 expression of lung cancer cells. The drug sensitivity increased after the two RNA interference plasmids were cotransfected into A549/TAX cells. The cytopathic effects and apoptosis of a combination of double shRNA interferences strategy on the tumor cells were more apparent than these of other treatment groups.
Based on our results, a combination of double shRNA interferences strategy downregulates significantly STMN1 and Bcl-2 RNA and protein expression. Targeting STMN1 and Bcl-2 expression with RNA interference sensitizes lung cancer cells to TAX. This strategy may be an effective approach to the antitumoral potency and provides a promising treatment paradigm for lung cancer.
Footnotes
Disclosure Statement
The authors declare no conflict of interest.