
Abstract
Gypenosides (Gyps) are triterpenoid saponins contained in an extract from Gynostemma pentaphyllum Makino and reported to induce apoptosis in human hepatoma cells through Ca2+-implicated endoplasmic reticulum (ER) stress and mitochondria-dependent pathways. The mechanism underlying the Gyp-increased intracellular Ca2+ concentration ([Ca2+]i) is unclear. Here, we examined Gyp-induced necrosis and apoptosis in human hepatoma HepG2 cells. Gyp-induced apoptotic cell death was accompanied by a sustained increase in [Ca2+]i level. Gyp-increased [Ca2+]i level was partly inhibited by removal of extracellular Ca2+ by Ca2+ chelator EGTA, store-operated Ca2+ channel (SOC) inhibitor 2- aminoethoxydiphenyl borate (2-APB), and ER Ca2+-release-antagonist 3,4,5-trimethoxybenzoic acid 8-(diethylamino) octyl ester (TMB-8). The strongest inhibitory effect was observed with TMB-8. EGTA, 2-APB, and TMB-8 also protected against Gyp-induced apoptosis in HepG2 cells. The combination of 2-APB and TMB-8 almost completely abolished the Gyp-induced Ca2+ response and apoptosis. In contrast, the sarco/endoplasmic-reticulum-Ca2+-ATPase (SERCA) inhibitor thapsigargin slightly elevated Gyp-induced [Ca2+]i increase and apoptosis in HepG2 cells. Exposure to 300 μg/mL Gyp for 24 hours upregulated protein levels of inositol 1,4,5-trisphosphate receptor and SOC and downregulated that of SERCA for at least 72 hours. Thus, Gyp-induced increase in [Ca2+]i level and consequent apoptosis in HepG2 cells may be mainly due to enhanced Ca2+ release from ER stores and increased store-operated Ca2+ entry.
Introduction
Hepatocellular carcinoma (HCC) is the sixth most frequently diagnosed cancer worldwide and the third leading cause of cancer-related mortality. 1 The incidence of liver cancer varies around the world. More than 80% of HCC occurs in developing countries, with 55% in China alone. 2 In 2008 in China, the incidence was 401,000 cases, and more than 371,000 patients died of the disease. 3 Despite extensive research into treatment for liver cancer, the overall survival rate is not optimistic because most HCCs are detected at an advanced stage in patients with underlying liver dysfunction. The increasing incidence and poor prognosis of HCC highlights the urgent need for novel effective therapeutic approaches.
Several compounds purified from plants and their derivatives show anti-cancer activity. An example is paclitaxel derived from Taxus brevifolia, which has been used in clinical treatment for more than 20 years. 4 Gypenosides (Gyps), containing ∼90 dammarane-type saponin glycosides, are the major components extracted from Gynostemma pentaphyllum (Thunb) Makino (Cucurbitaceae), 5 a widely used traditional medicine in China. The effective ingredient of Gyp is the hydroxy group combined with the 20th or 21st carbon in the dammarane-type ring. With various pharmacological properties such as anti-inflammatory, 6 anti-oxidative, 7,8 immune regulatory, 9 anti-hyperlipidemic, 10 adhesion inhibitory, 9 and anti-cancer activity, 11 Gyp has been used for treating hepatitis, 12 hyperlipoproteinemia, 13 cardiovascular disease 14 , and tumor. 11,15,16
The main mechanism of the anti-cancer activity of Gyp is inhibiting proliferation and inducing apoptosis of tumor cells. Gyp could induce apoptosis in human hepatoma cells by regulating the ratio of Bcl-2 to Bax, releasing mitochondrial cytochrome c and activating a caspase cascade. 11 In human lung A549 cancer cells, Gyp exerted its anticancer properties by inducing G0/G1 arrest and apoptosis via activation of caspase-3. 17 Further, Gyp inhibited invasion and migration of human tongue cancer cells by downregulating NF-kappaB and matrix metalloproteinase-9. 18 Subsequent study showed that an increase in intracellular Ca2+ concentration ([Ca2+]i) plays a pivotal role in the initiation of Gyp-triggered apoptosis. 19 However, how this Ca2+ signal arose was unexplored. [Ca2+]i elevation in non-excitable cells mainly originates from increased intracellular Ca2+ release from endoplasmic reticulum (ER) Ca2+ stores and plasmalemma Ca2+ entry from store-operated Ca2+ entry. These two pathways of Ca2+ are implicated in apoptosis of cancer cells. 20,21 The molecular components of store-operated Ca2+ channels (SOCs) in hepatocytes are stromal interacting molecule 1 (STIM1) acting as a sensor for level of Ca2+ stored in the ER and calcium release-activated calcium channel protein 1 (Oria1) constituting pore-forming subunits of the SOCs. 22 For the ER, Ca2+ release is mainly mediated by type 1 inositol 1,4,5-trisphosphate receptor (IP3R), whereas Ca2+ uptake is mediated by sarco/endoplasmic-reticulum-Ca2+-ATPases (SERCAs), which pump calcium against the Ca2+ gradient into the ER. 23
We aimed to investigate the mechanisms underlying the Gyp-increased [Ca2+]i content and related apoptosis in the human HCC cell line HepG2 by using the Ca2+ chelator EGTA, SOC antagonist 2-aminoethoxydiphenyl borate (2-APB), ER Ca2+ release inhibitor 3,4,5-trimethoxybenzoic acid 8-(diethylamino) octyl ester (TMB-8), and SERCA inhibitor thapsigargin.
Materials and Methods
Chemicals and reagents
Gyp, dimethyl sulfoxide (DMSO), EGTA, 2-APB, TMB-8, and thapsigargin were from Sigma Chemical Co. Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum, penicillin–streptomycin, trypsin-ethylenediaminetetraacetic acid, and Hanks' Balanced Salt Solution (pH 7.4) were from Gibco. Cell Counting Kit 8 (CCK8) was from Dojin Laboratory. Fluo-3 AM and Pluronic F-127 were from Biotium. Antibodies for STIM1, Orai1, IP3R-1, SERCA, and horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G were from Santa Cruz Biotechnology. EGTA and TMB-8 were directly added to phosphate buffered saline (PBS), whereas 2-APB and thapsigargin were dissolved in DMSO 1:1000 as stock solutions. At this concentration, DMSO does not affect the induction of habituation. Just before each experiment, aliquots were diluted to the desirable concentration with culture medium.
Culture and treatment of cells
HepG2 cells were from the American Type Culture Collection and cultured in DMEM containing 100 U/mL penicillin and 100 μg/mL streptomycin at 37°C under humidified 5% CO2–95% air and subcultured every 4–6 days. When grown to ∼80% confluence, cells were washed twice with PBS before being treated with various agents in complete medium. Control cells were untreated. For inhibition experiments, cells were treated with the indicated reagent and 300 μg/mL Gyp for 24 hours.
Cell viability assays
Cell viability was estimated by use of a CCK based on the ability of cells to cleave tetrazolium salts WST-8 [2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H- tetrazolium nosodium salt] by dehydrogenases. Cells were seeded in 96-well plates at 104 cells/well in culture medium and treated with Gyp for the indicated times, then stained with 5 mg/mL CCK8 reagent at 37°C under 5% CO2–95% air for 2 hours. The absorbance was measured by use of a Varioskan Flash spectral scanning multimode reader (Thermo Electron Corp.) at 450 nm. The cell viability index (% of control) was calculated as follows: (ODexperiment−ODblank/ODcontrol−ODblank)×100%, where ODexperiment is the absorbance of cells exposed to different treatments, ODcontrol is the absorbance of control cells without treatment, and ODblank is the absorbance of wells without HepG2 cells.
Analysis of apoptosis
Apoptosis of HepG2 cells was determined by terminal transferase-mediated dUTP-fluorescensin nick end labeling (TUNEL; Millipore). Briefly, cells were fixed with 1% paraformaldehyde, washed twice in PBS, and post-fixed in freshly prepared ethanol plus acetic acid (2:1), before endogenous peroxidase was blocked in 3% H2O2. Then, cells were stained with terminal deoxynucleotide transferase reaction mixture for 1 hour at 37°C in the dark. After being washed, cells were incubated with anti-digoxigenin peroxidase-conjugated antibody for 30 minutes. Apoptotic cells were detected after incubation in 3,3′-diaminobenzidine chromogen. The percentage of TUNEL-positive cells (apoptotic cells) was analyzed by use of Image-Pro Plus 6.0.
Measurement of cytosolic free calcium ([Ca2+]i) concentration
[Ca2+]i content in HepG2 cells was evaluated as fluorescence intensity of Fluo-3/AM determined by flow cytometry. After treatment with Gyp and other agents in six-well plate for various times, HepG2 cells were loaded with fluo-3/AM and pluronic F127 (final concentrations 5 μM and 0.1%, respectively) at 37°C for 1 hour in the dark and collected with trypsin. The fluorescence intensity was measured with FACS Calibur flow cytometer (Becton Dickinson) by detecting green fluorescence with FL1. Results of 10,000 fluorescent events by CellQuest were analyzed with FCS Express 4.0 (De Novo Software) and expressed as mean fluorescence intensity of 10,000 cells.
Western blot analysis
Total protein was extracted from lysed cells and underwent sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Then proteins were transferred onto polyvinylidene fluoride membrane, which was blocked in 5% nonfat milk for 1 hour, then incubated overnight with the primary antibodies rabbit anti-STIM1 (1:1000), rabbit anti-Orai1 (1: 500), rabbit anti-IP3R-1 (1:1000), and rabbit anti-SERCA (1:300), then horseradish peroxidase-conjugated secondary antibody. Membranes were washed thrice with Q3Tris-HCl-buffered salt solution with Tween, and then visualized by use of the Immobilon Western Chemiluminescent HRP Substrate kit (Millipore). The intensity of the immunoreactive labeling was analyzed by use of ImageJ (National Institutes of Health) and standardized against β-actin.
Statistical analysis
Two-tailed t test was used for analyzing two groups and one-way analysis of variance for three or more groups. All data were represented as mean±standard deviation of three independent experiments. Analysis involved use of SPSS 17.0 (SPSS, Inc.). p<0.05 was considered statistically significant.
Results
Gyps effects on cell viability and apoptosis in HepG2 cells
Treatment with Gyp dose- and time-dependently decreased HepG2 cell viability (Fig. 1A, B). With 300 μg/mL Gyp, significant cell death was observed from 6 hours (p<0.01 as compared with no treatment). To detect cell apoptosis induced by Gyp, DNA fragmentation of HepG2 cells was evaluated by in situ DNA TUNEL. Compared with no treatment, 50–400 μg/mL Gyp for 24 hours dose-dependently increased the rate of apoptosis in HepG2 cells, which plateaued at high concentrations, and time-dependently increased apoptosis with 300 μg/mL Gyp.
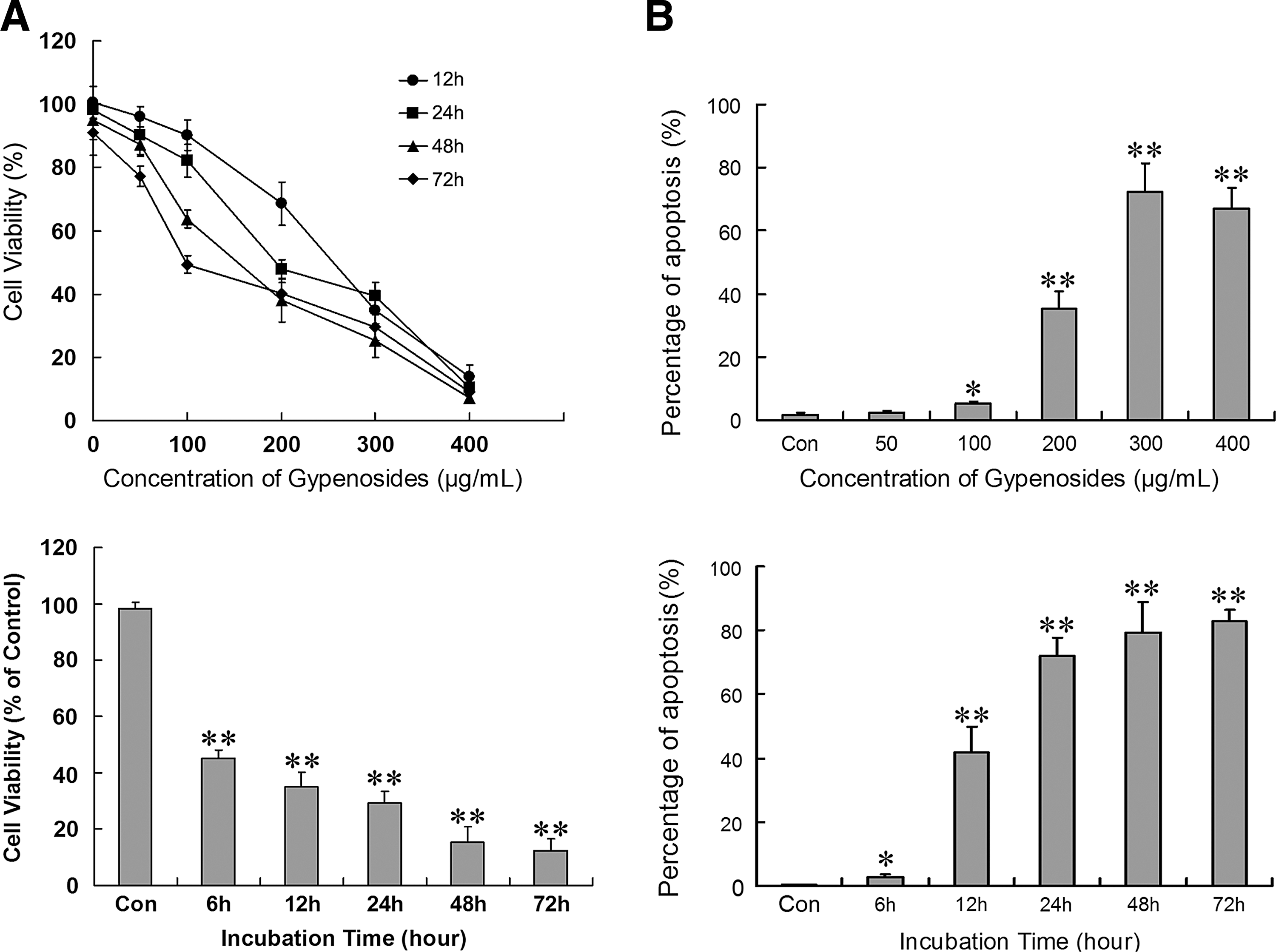
Effect of Gyps on cell viability and apoptosis in human hepatocellular carcinoma HepG2 cells.
Gyp effect on [Ca2+]i content in HepG2 cells
To determine the effect of 24-hours Gyp treatment on intracellular Ca2+ content, Ca2+ production was quantified by flow cytometry and analyzed by Fluo-3 fluorescence intensity. Gyp at 50–400 μg/mL for 24 hours dose dependently induced Ca2+ production in Ca2+-containing medium as compared with no treatment (Fig. 2).

Gyp-induced production of Ca2+ in HepG2 cells. HepG2 cells (5×105 cells/well) were treated with concentrations of Gyp for 24 hours, then stained with Fluo3/AM; Ca2+ content was determined by flow cytometry. The leftmost histogram represents the control group, the right five histograms represent various Gyp concentrations (50–400 μg/mL from left to right). Bottom results are mean±SD relative from three repeat experiments (n=6). *p<0.05, **p<0.01 compared with the control.
Effect of EGTA, 2-APB, TMB-8, and thapsigargin on Gyp-increased [Ca2+]i content in HepG2 cells
Chemical signals induce [Ca2+]i from two main sources: external milieu and internal Ca2+ stores, mainly associated with the ER in hepatocytes. We explored the Ca2+ pathway of Gyp-induced response in HepG2 cells. Removal of extracellular free Ca2+ by the Ca2+ chelator EGTA (0.5 mM) decreased 300-μg/mL Gyp-increased [Ca2+]i content by 42.1% (p<0.01), and the store-operated Ca2+ influx inhibitor 2-APB (50 μM) reduced Gyp-increased [Ca2+]i content by 38.9% (p<0.01), with no difference between the agents in inhibition (Fig. 3). Ca2+ from an extracellular area may participate in Gyp-increased [Ca2+]i content, with store-operated Ca2+ entry the primary source. To determine the contribution of Ca2+ release from intracellular stores to the Gyp-increased [Ca2+]i content, HepG2 cells were treated with 300 μg/mL Gyp for 24 hours and with TMB-8, the antagonist of IP3R-mediated ER Ca2+ release, or thapsigargin, a SERCA inhibitor. TMB-8 (100 nM) significantly suppressed Gyp-increased [Ca2+]i content, by 55.3%, and thapsigargin (10 μM) slightly increased Gyp-increased [Ca2+]i content, by 15.6%, although not significantly (Fig. 3). The combination of 2-APB and TMB-8 almost completely abolished the Gyp-induced Ca2+ response, so store-operated Ca2+ entry and IP3R-mediated ER Ca2+ release are two major sources of Gyp-increased [Ca2+]i content.
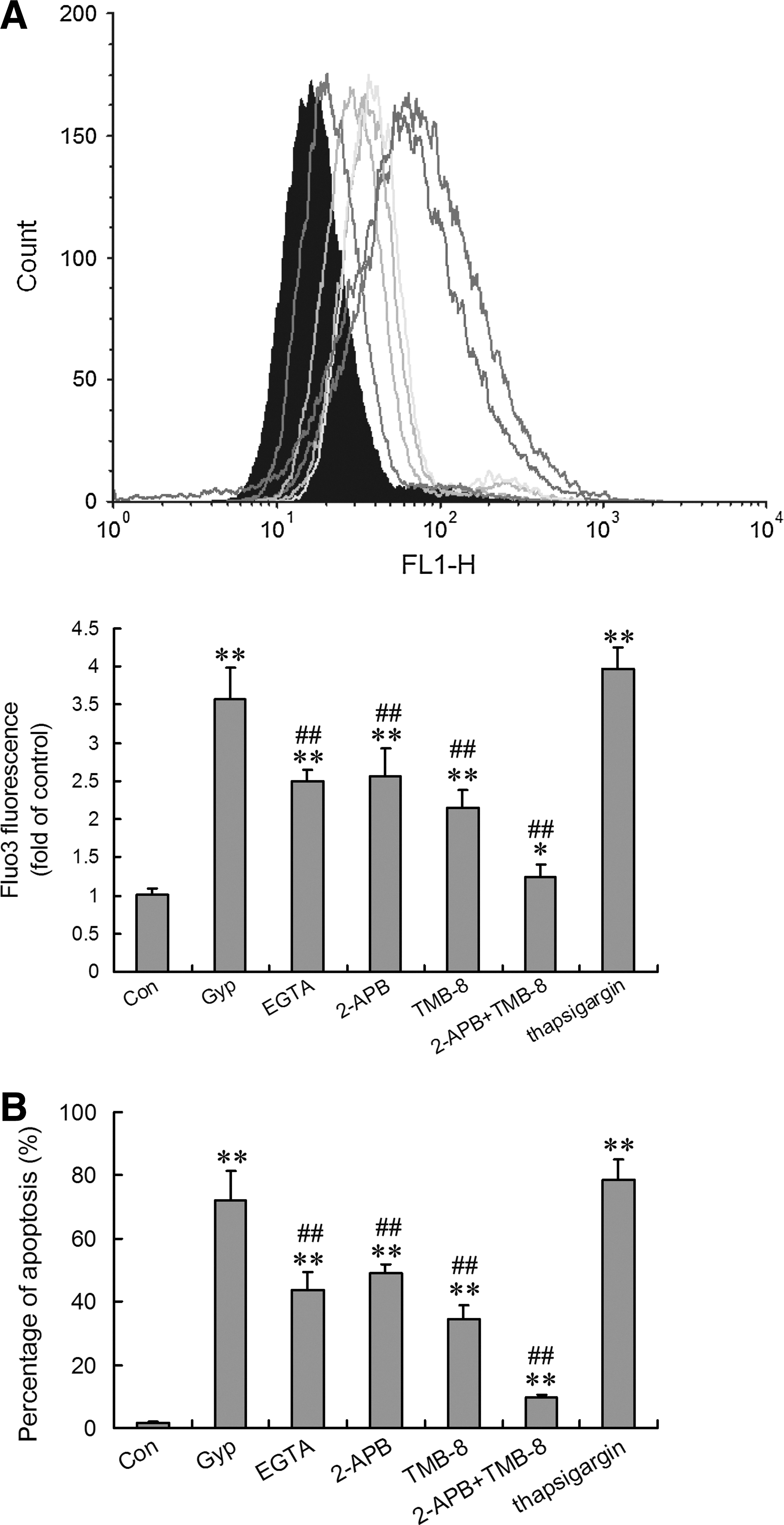
Effect of EGTA, 2-APB, TMB-8, and thapsigargin on Gyp-increased [Ca2+]i content and apoptosis in HepG2 cells. The experiments were performed in Ca2+-containing medium. In blocker- or modulator-treated groups, the reagent was added simultaneously with 300 μg/mL Gyp for 24 hours. The concentration was 0.5 mM for EGTA, 50 μM for 2-APB, 100 nM for TMB-8, and 10 μM for thapsigargin.
Effect of EGTA, 2-APB, TMB-8, and thapsigargin on Gyp-induced apoptosis in HepG2 cells
We identified the possible role of the two sources of [Ca2+]i increase, ER-mediated Ca2+ release and SOCs, in Gyp-induced HepG2 cell apoptosis. In accordance with enhanced [Ca2+]i, content incubating HepG2 cells with EGTA, 2-APB, or TMB-8 significantly suppressed Gyp-enhanced apoptosis, the strongest effect with TMB-8 (Fig. 3). Co-treatment with 2-APB and TMB-8 greatly reduced Gyp-induced apoptosis to almost the same level as no treatment. In contrast, thapsigargin moderately increased Gyp-induced apoptosis. Thus, Gyp-enhanced Ca2+ release from the ER and Ca2+ entry through SOCs played an important role in Gyp-induced apoptosis in HepG2 cells.
Gyps affect the expression of STIM1, Orai1, IP3R-1, and SERCA in HepG2 cells
The ER membrane (endomembrane) is endowed with SERCA pumps and Ca2+ release channels; the former accumulate Ca2+ in the lumen against its electrochemical gradient, whereas the latter allow Ca2+ to diffuse out of the store on opening of the channels. 24 STIM1 and Orai1 are two major molecular components of SOCs in hepatocytes and may be necessary and sufficient for SOCs. 25 To further study the mechanism responsible for the ER and SOCs in Gyp-increased [Ca2+]i content and cancer resistance, HepG2 cells were treated with 300 μg/mL Gyp for different times to detect the effect of Gyp on the protein levels of STIM1, Orai1, IP3R, and SERCA. HepG2 cells were also treated with PBS for the same times indicated to avoid the interference of time progress. Almost no changes could be found in the protein levels of STIM1, Orai1, IP3R, and SERCA when HepG2 cells were treated with PBS (data not shown). However, Gyp at 300 μg/mL significantly increased the levels of STIM1, Orai1, and IP3R-1 and reduced that of SERCA at all times (Fig. 4), which suggests that the effect of Gyp could last for at least 72 hours. Further, the increased expression of STIM1 was parallel to that of Orai1, so the combination of the two molecules may be needed in the Gyp-increased SOC level.
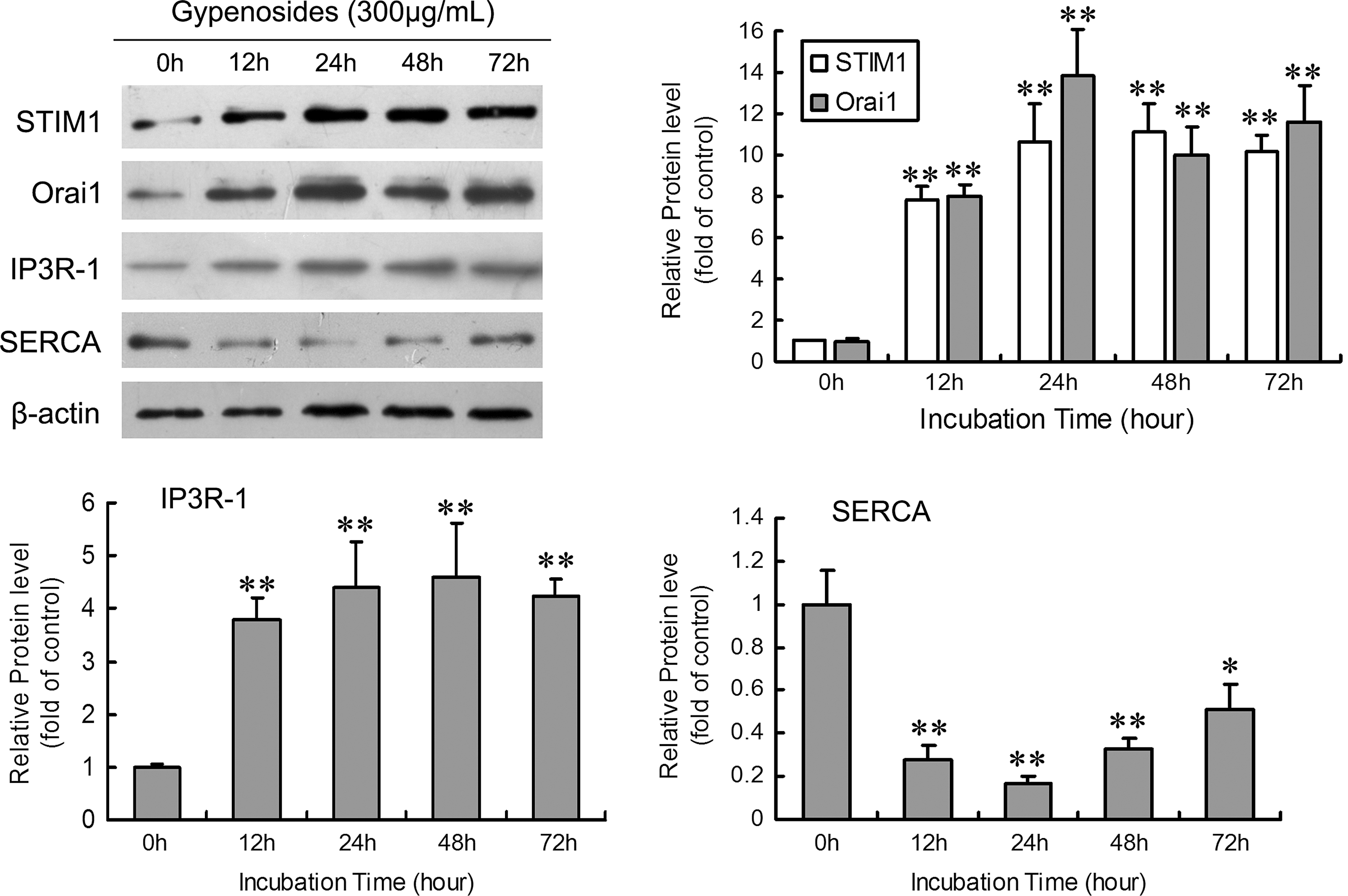
Representative western blot analysis of protein levels of Ca2+ channel-associated proteins in HepG2 cells with exposure to Gyp. HepG2 cells were treated with Gyp at 300 μg/mL for various times. Results are mean±SD relative to the control from three experiments (n=6). *p<0.05, **p<0.01 compared with the control.
Discussion
Various studies confirmed that Gyp has a remarkable preventive effect on the development of human cancers by inhibiting proliferation and inducing apoptosis of tumor cells. 19,26 Buffering the increased [Ca2+]i content in Gyp-treated Huh-7 cells blocked Gyp-induced mitochondrial dysfunction and apoptosis, which suggested the critical role of increased [Ca2+]i content in the initiation of Gyp-triggered apoptotic death. 19 Excess Ca2+ accumulation within mitochondria because of cellular Ca2+ overload is one of the main causes of mitochondrial permeability transition, which allows for release of mitochondrial apoptogenic factors, such as cytochrome c into the cytoplasm, which in turn activates an apoptosis-executing caspase cascade. 27 However, the precise source of Ca2+ in Gyp-exposed hepatocellular cancer cells remained unclear. Here, we explored the underlying mechanisms in Gyp-increased [Ca2+]i content and the associated apoptosis in HepG2 cells.
CCK8 assay of cell viability demonstrated that Gyp induced cell death from 50 to 400 μg/mL, the “safe” range of Gyp in human fibroblasts and primary rat heart endothelial cells. 11 In situ DNA fragmentation assay by TUNEL revealed Gyp-treated cells with increased apoptosis rather than necrosis. Gyp treatment greatly increased the number of TUNEL-positive cells dose and time-dependently as compared with no treatment, which suggests that Gyp induced apoptotic cell death in human HepG2 cells. Flow cytometry confirmed the effect of Gyp on increasing [Ca2+]i content in HepG2 cells. The [Ca2+]i elevation was concentration dependent between 50 and 400 μg/mL. The same effect of Gyp was observed in human tongue cancer SCC-4 cells 26 and human myeloid leukemia HL-60 cells 28 and may be a common mechanism underlying Gyp-induced apoptosis in cancer cells. The apoptotic-inducing effect of Gyp increasing [Ca2+]i content is associated with reactive oxygen species generation, Bcl-2/Bax expression and redistribution, and mitochondrial dysfunction. 19
In non-excitable cells, including hepatoma cells, Ca2+ influx from the extracellular area and Ca2+ release from the ER Ca2+ stores are two major sources of Ca2+ in hormone- and chemical-induced increase in [Ca2+]i content. 23 We found that removal of extracellular Ca2+ and ER Ca2+ release inhibitor TMB-8 partially reduced the Gyp-increased [Ca2+]I content in HepG2 cells, so the Gyp-increased [Ca2+]i content may result from both extracellular Ca2+ influx and depleted intracellular Ca2+ stores. Among all Ca2+-permeable channels confirmed to express in hepatocytes, SOCs provide a major pathway for Ca2+ entry. 29 SOCs are plasma-membrane Ca2+ channels activated by decreased Ca2+ content in the ER. The SOC antagonist 2-APB significantly reduced the Gyp-increased [Ca2+]i content in HepG2 cells, which did not differ from the EGTA effect, which suggests that SOCs may be the primary source of Gyp-induced Ca2+ influx through the plasma membrane. In addition, EGTA, 2-APB, and TMB-8 suppressed Gyp-induced apoptosis in HepG2 cells, so increased Ca2+ entry through SOCs and elevated Ca2+ release from the ER was implicated in Gyp-associated apoptosis in cancer cells. Combined treatment of 2-APB and TMB-8 almost completely abolished the Gyp-induced Ca2+ response and apoptosis, so SOC and ER Ca2+ release may be sufficient for Gyp-increased [Ca2+]i content and related apoptosis.
The most important molecular component of the Ca2+ handling machinery of the ER is IP3R, a ligand-gated channel that discharges Ca2+ from ER stores in response to stimulation. 30 The release of Ca2+ from ER stores by IP3R is directly responsible for mitochondrial Ca2+ overload and apoptosis because cells deficient in IP3R are resistant to apoptosis. 31 Calcium is actively accumulated in the ER by SERCA-dependent calcium transport, the only calcium uptake mechanism in this organelle. 32 Significant changes in SERCA expression during cell differentiation or tumorigenesis can lead to modified ER calcium storage. Function-based genetic screening by systematic RNA interference identified STIM1 and Orai1 proteins as 2 essential molecular players in SOCs. 25 STIM1 senses depletion of ER Ca2+ stores and transmits the signals to the plasma membrane, thus opening SOCs. 33 Orai1 acts as a pore subunit of SOCs that is gated directly or indirectly by STIM1. 34 We found the protein levels of IP3R-1, STIM1, and Orai1 in HepG2 cells increased with 300 μg/mL Gyp, which suggests that the enhanced ER Ca2+ release and SOCs probably resulted from increased IP3R and SOC protein expression. Gyp also significantly decreased the protein level of SERCA, so impaired uptake of Ca2+ in the ER may be involved in Gyp-increased [Ca2+]i content in HepG2 cells.
Conclusions
In conclusion, we confirmed the key role of sustained increase in [Ca2+]i content in Gyp-triggered apoptotic signaling in human HepG2 cells. Gyp upregulated IP3R and SOC proteins (STIM1 and Orai1) and downregulated SERCA protein, which led to increased extracellular Ca2+ influx and ER Ca2+ release and decreased ER Ca2+ uptake. This process perturbed intracellular calcium homeostasis and finally triggered Ca2+-dependent apoptosis. These findings provide new insights into the possible pathway of Gyp-induced apoptosis in human HepG2 cells.
Footnotes
Acknowledgments
We acknowledge the technical support provided by the staff of the Cancer Research Institute in the First Affiliated Hospital at China Medical University. We thank Dr. Laura Smales for linguistic advice. This work was financially supported by the Liaoning Provincial Natural Science Foundation (20042073) and Liaoning Provincial Scientific and Technological Project (2009225008-1).
Disclosure Statement
The authors declare that no competing financial interests exist.