
Abstract
Cationic antimicrobial peptides (CAPs) with antitumor activity have potential for use as novel antitumor agents because of their lower risk for induction of resistance. Of these peptides, magainin II (MG2) exhibited cytotoxicity in tumor cells only at high concentrations, likely due to the inefficiency of MG2 in cell membrane binding and cell entry. Conjugation to a cell-penetrating peptide (CPP) might enhance the cytotoxicity of MG2 in tumor cells. Here, we constructed a fusion peptide MG2A by conjugating MG2 to the N-terminus of the CPP penetratin (Antp). It was found that the fusion peptide MG2A is more potent than unconjugated MG2 at tumor cell killing. The IC50s of MG2A for the tumor cells tested were at least 30 times lower than the IC50s of unconjugated MG2. These data indicate that conjugation to Antp significantly enhanced the cytotoxicity of MG2 in tumor cells. Moreover, the IC50s of MG2A for tumor cells are within 2 to 3 μM, which are about three to five times lower than the IC50 for normal cells. Furthermore, chondroitin sulfate (CS) was found to be overexpressed on the surface of the tested tumor cells, and the cytotoxicity of MG2A could be inhibited by the addition of exogenous CS. These results suggest that binding of Antp to CS on tumor cells might be one important cause for the selective cytotoxicity of MG2A in tumor cells. Taken together, conjugation of MG2 to Antp can significantly enhance its antitumor activity, and the fusion of CAP to Antp might be an alternative for cancer-targeted therapy.
Introduction
Cationic antimicrobial peptides (CAPs) with antitumor activity have been suggested as promising agents for antitumor therapy. These peptides are able to kill chemo-resistant tumor cells and have a low risk for inducing multi-drug resistance in tumor cells. 1,2 Moreover, they exert a higher specific cytotoxicity to tumor cells than to normal healthy cells when compared with conventional cancer chemotherapy. 3,4 The reason for the higher specificity of CAPs to tumor cells likely lies in their interaction with the plasma membrane. The plasma membrane of many tumor cells overexpresses negatively charged molecules, such as phosphatidylserine and glycosylated mucins, 5,6 whereas the plasma membrane of normal cells is neutrally charged overall. 7 The positive charge of the CAPs enables easier binding to the negatively charged tumor cell membrane through electrostatic interactions. 2 Recent studies have indicated that CAPs with antitumor activity appear to cause necrosis by disruption of cell membrane. 8,9 In addition, after binding to a tumor cell, it is likely that some CAPs could subsequently gain access to the cells and cause apoptosis by disrupting mitochondria. 1,3,10,11 Therefore, membrane binding and cell entry is thought to play a crucial role in the cytotoxicity of CAPs to tumor cells.
One member of the membrane-bound CAP family is magainin 2 (MG2), which was isolated from the skin of the African clawed frog Xenopus laevis. 12 This peptide displays specific cytotoxic activity against a variety of tumor cells in vitro. Nevertheless, MG2 did not display robust antitumor activity, with an IC50 value of more than 100 μM. 8,13 Thus, it is necessary to enhance the cytotoxic activity of MG2 for its further development as cancer treatment. Previous studies have revealed that MG2 can induce tumor cell death by cell membrane and mitochondrial membrane lytic effects. 1 Taking into account the weak electrostatic attraction of MG2 to the membranes of tumor cells, we hypothesize that the observed cytotoxicity of MG2 in tumor cells at high concentrations results from its poor membrane binding and poor cell entry. Early studies have shown that changing the amino-acid composition of MG2 by increasing the surface positive charge can enhance its antitumor activity. 13,14 In addition, a vector that can mediate effective membrane binding and cell entry may also increase the cytotoxicity of MG2 toward tumor cells. 15,16
Cell-penetrating peptides (CPPs) are effective and nontoxic vectors for drug delivery. 17,18 Of these peptides, penetratin (Antp), derived from the third helix of the homeodomain of Antennapedia, is efficient in the translocation of conjugated cargo across the plasma membrane. 19,20 In addition, our group found that Antp can bind chondroitin sulfate (CS), a proteoglycan that is overexpressed on the tumor cell surface. 21 Consequently, conjugating MG2 to Antp might enhance the cytotoxicity of MG2 to tumor cells through improving cell membrane binding and cell entry ability.
We report a fusion peptide MG2A constructed by conjugation of MG2 to the N-terminus of Antp. First, the cytotoxicity of MG2A and MG2 in tumor cells was compared. Subsequently, we compared the cytotoxicity of MG2A in tumor cells and normal cells. Lastly, the mechanism of MG2A-mediated cell death and the in vivo antitumor effect of MG2A were investigated.
Methods
Preparation of peptides
In order to study whether the Antp peptide can enhance the cytotoxicity of MG2 to tumor cells, we constructed the fusion peptide MG2A by conjugation of MG2 to the N-terminus of Antp. According to the description by Evers et al., a flexible linker containing two glycine residues was inserted into the fusion peptide to reduce the interaction of MG2 and Antp. 22 A control peptide named URP that should show no cytotoxicity and the unconjugated MG2 and Antp peptide were also synthesized by Genescript, Inc. In the standard quality control analysis, all peptides were identified with high-performance liquid chromatography and mass spectrometry. As summarized in Table 1, the peptides used were >95% pure, initially dissolved to 400 μM in serum-free RPMI 1640 medium and separated into aliquots in sterile microcentrifuge tubes.
amino acid.
Cell culture
Cells used in the experiments, including African green monkey kidney cells (Vero E6), human proximal tubular cells (HK-2), human cervical carcinoma cells (HeLa), human lung non-small-cell carcinoma cells (A549), human skin malignant melanoma cells (A375), and rat glioma cells (C6), were purchased from the American Type Culture Collection (ATCC). The cells were cultured in either Dulbecco's modified Eagle's medium or RPMI 1640 containing 10% fetal bovine serum, 2 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin and grown at 37°C in 5% CO2.
Cytotoxicity assays
Aliquots of medium containing 1×104 cells/well were distributed into a 96-well plate and incubated for 12–18 hours to allow for attachment to the plate. After washing once with phosphate-buffered saline (PBS), increasing concentrations of peptides in 100 μL of serum-free medium supplemented with 2% bovine serum albumin were added, and then, cells were incubated at 37°C for 30 minutes. Next, the cytotoxicity assay for the peptides was performed using the colorimetric tetrazolium salt-based Cell Counting Kit-8 (CCK-8; Dojin Laboratories). 16,21 Briefly, 10 μL of CCK-8 solution was added to each well, and the plates were again incubated at 37°C. After 2–4 hours of incubation, the absorbance in each well was measured at 450 nm. Cells incubated with culture medium alone represented 100% viability and were regarded as the control to allow for estimation of the percent viability in the cell samples. IC50 values were calculated as the concentration at which cell viability was reduced to 50% when compared with control cells. Each sample was performed in triplicate.
To further assess the cytotoxicity of MG2A, a live/dead viability kit based on two nucleic acid stains, including SYTO 9 green-fluorescent stain and the propidium iodide (PI) red-fluorescent stain, was used. All cells stained fluorescent green, and cells with a damaged membrane stained fluorescent red. After incubation with peptides for 20 minutes, the cells were stained with SYTO 9 and PI for 5 minutes in the dark and then observed under a fluorescence microscope.
Assessment of apoptosis by annexin V and PI staining
Phosphatidylserine (PS) expression in the outer leaflet of the plasma membrane starts early after the onset of apoptosis in cells. Annexin V can bind to PS with high affinity. Hence, cells that are undergoing apoptosis are often identified by staining with FITC-annexin V (appearing green) and PI (appearing red). With this method, vital cells, apoptotic cells, and necrotic cells will be annexin V−/PI−, annexin V+/PI−, and annexin V+/PI+, respectively. 16,21 After incubation with peptides for 15 minutes, the cells (2.5×105) were stained with FITC-annexin V and PI and observed under a fluorescence microscope or analyzed by flow cytometry.
Assessment of apoptosis by monitoring mitochondrial depolarization
Changes in the mitochondrial membrane potential of cells was monitored using the membrane potential-sensitive fluorescent JC-1 dye. JC-1 forms fluorescent red aggregates on localization to healthy mitochondria, while the monomer with green fluorescence is localized to the cytoplasm. Thus, a decrease in the red-to-green fluorescence ratio reflects mitochondrial membrane depolarization. 16,21 We simultaneously measured the red and green fluorescence by flow cytometry. The staining of mitochondria and cell nuclei was performed with JC-1 and DAPI, respectively, and then observed by a fluorescence microscope.
Assessment of apoptosis by nuclear morphology
Apoptosis at each time point was analyzed by nuclear morphology changes based on karyopyknosis and nuclear fragmentation. Nuclei were stained with DAPI, a blue fluorescent dye. After treatment with MG2A for 0–16 hours, cells were stained with DAPI for 5 minutes and observed under a fluorescent microscope.
Involvement of caspase in peptide-mediated cell death
The pan-caspase inhibitor z-VAD-Fmk was used to investigate involvement of the caspase in peptide-mediated cell death. HeLa cells were preincubated with 100 μM of inhibitor for 2 hours and then treated with MG2A for an additional 30 minutes. Cell viability was determined by the CCK-8 kit. The cytotoxicity of MG2A to cells that were preincubated with inhibitor and those that were not was compared.
The expression of CS on cells and the effect of exogenous CS inhibition on the cytotoxicity of MG2A
The expression of CS on tumor cells and normal cells was detected by FACS using an antibody against CS (CS-56; Santa Cruz Biotechnology, Inc.) with an isotype antibody as a control. 21 Cells (3×105) were incubated with anti-CS antibody at 37°C for 2 hours, then incubated with a FITC-labeled secondary antibody, and analyzed by flow cytometry. To investigate the inhibition of exogenous CS on MG2A-induced cell death in HeLa and A549 cells, MG2A was incubated with increasing concentrations of CS at 37°C for 5 minutes. Subsequently, the mixture was added to the cells, and the cytotoxicity was determined by CCK-8. Sodium hyaluronate (HA) was used as a glycosaminoglycan control.
In vivo antitumor efficacy of MG2A in tumor xenografts
All protocols for the animal experiments were approved by the University Animal Care and Use Committee. HeLa cells (6×106) suspended in 100 μL of PBS were transplanted subcutaneously in the right flank of BALB/c (nu/nu) mice. At the onset of a palpable tumor (approximate volume of 50 mm3), mice were randomly divided into two groups. The MG2A-treatment group (n=4) was injected intraperitoneally with 15 mg/kg MG2A, and the control group (n=4) was injected with the equivalent volume of PBS daily for 5 days. The tumor volume (mm3) was calculated as length×width2×0.5.
To directly explore the death of tumor cells in vivo, a single dose of MG2A (100 μg) in PBS (100 μL) or PBS alone was injected intratumorally into the tumor (volume: 1000 mm3). Twenty-four hours postinjection, the animals were euthanized, and the tumor tissues were excised, paraffin embedded, sectioned, and stained with H&E to examine the histological architecture.
Statistical analysis
All observations were confirmed by at least three independent experiments. Results were expressed as the means±SD, and analysis of variance was used to analyze the differences. The significant level was defined as p<0.05.
Results
Antp enhances cytotoxicity of MG2 in tumor cells
After treatment with 5 μM of MG2A for 30 minutes, HeLa and A549 cells became round and nonadherent, and the intercellular contents leaked out from the cells. However, no obvious morphological changes could be observed for HeLa and A549 cells with 5 μM of Antp, MG2, and the control peptide (UPR) (Fig. 1A). The CCK-8 assay revealed that 5 μM of MG2A caused an 80% death of HeLa and A549 cells. The IC50 of MG2A in HeLa and A549 cells was 2–3 μM. In contrast, more than 80% of HeLa and A549 cells were still alive after treatment with 15 μM of Antp, MG2, or UPR (Fig. 1B). These data show that coupling MG2 with Antp can enhance the cytotoxicity of MG2 to tumor cells.
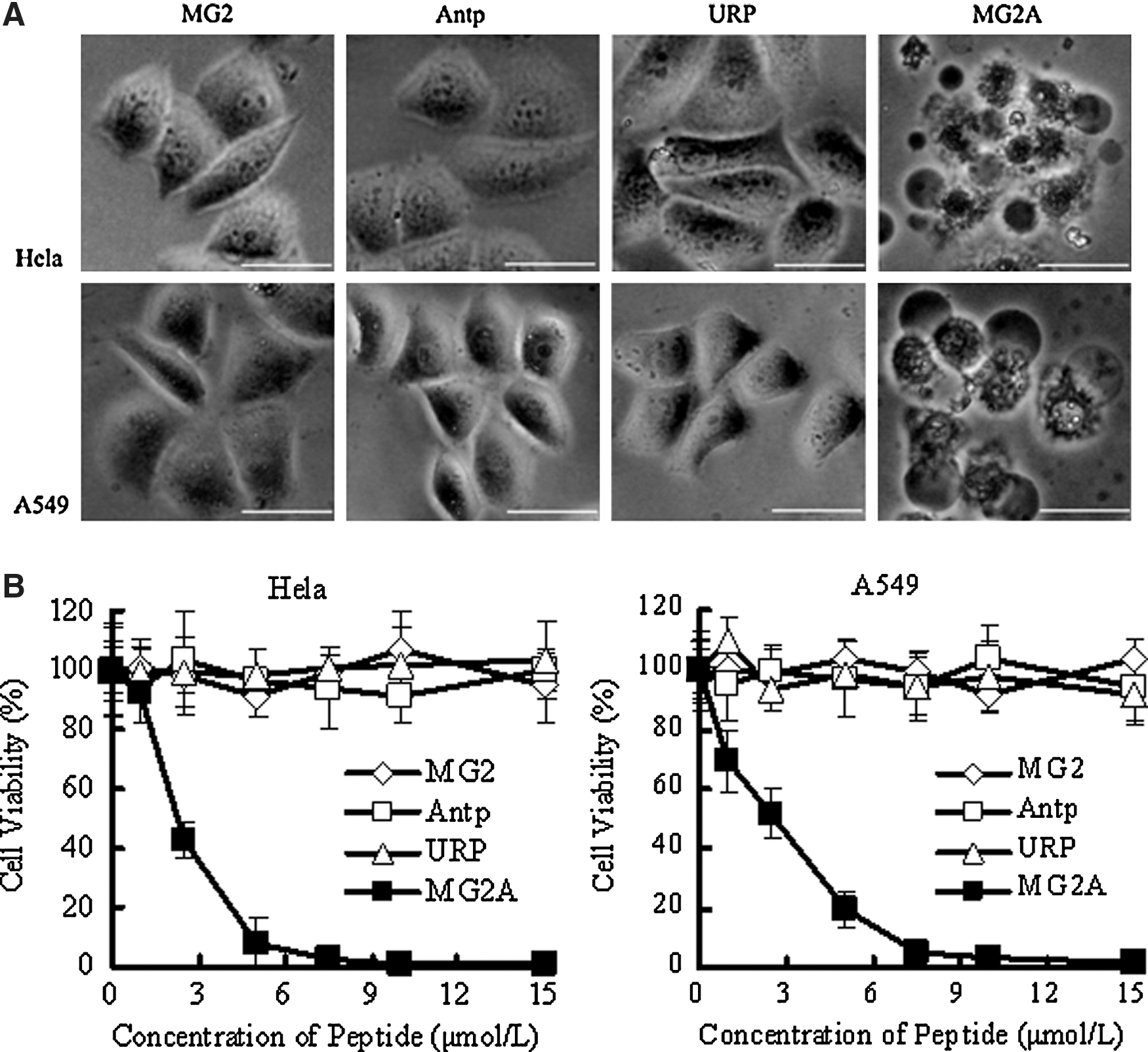
Antp enhances cytotoxicity of MG2 in tumor cells.
Selective cytotoxicity of MG2A to tumor cells
Assessment of the cytotoxicity of MG2A to various tumor and normal cells using CCK-8 determined that more than 80% of HeLa, A549, A375, and C6 tumor cells were dead after treatment with 5 μM of MG2A; however, 70%–80% of the normal, VeroE6, and HK-2 cells were alive even after a 7.5 μM treatment (Fig. 2A). The IC50 of MG2A in normal cells was three to five times higher than that of MG2A in tumor cells (IC50 values of 2–3 μM). After treatment with 5 μM of MG2 or MG2A for 20 minutes, cells were simultaneously stained with SYTO 9 and PI. As shown in Figure 2B, almost all of the MG2-treated HeLa and Vero E6 cells were alive (green). However, in MG2A-treated cells, 90% of HeLa cells were dead (red), and 80% of VeroE6 cells were alive. The results just cited indicate that MG2A is selectively cytotoxic to tumor cells.

Selective cytotoxicity of MG2A to tumor cells.
Involvement of caspase in MG2A-induced cell death
Double staining with annexin V and PI was used to examine whether HeLa cells treated with MG2A underwent apoptosis. After treatment with 1 μM of MG2A for 15 minutes, cells that were annexin V+/PI− (indicative of apoptosis) and annexin V+/PI+ (indicative of necrosis) were observed under a fluorescence microscope (Fig. 3A). Flow cytometry analysis revealed that the number of apoptotic cells increased but then decreased as the concentration of MG2A increased; in contrast, the necrotic cells increased gradually. After incubation with 0, 5, 7, and 9 μM of MG2A, the percentage ratio of apoptotic to necrotic cells was 4.84/0.97, 24.66/14.86, 41.90/35.07, and 12.16/73.43, respectively (Fig. 3B). Changes in mitochondrial membrane potential were detected with the double staining of JC-1 and DAPI. As shown in Figure 3C, JC-1 formed aggregates and emitted red fluorescence at the mitochondria of MG2A-free HeLa cells. Granulo-mitochondria were observed clearly. However, the granulo-mitochondria disappeared after incubation with MG2A. Flow cytometry results show that before and after treatment with MG2A, the ratio of red fluorescence and green fluorescence decreased from 94/6 to 4/96 (Fig. 3D). Therefore, MG2A can induce the cells' mitochondrial membrane unpolarized. In addition, nuclei condensation and DNA fragmentation were also observed in the peptide-treated cells (Fig. 3E). The peptide-mediated cell death could be rescued by addition of pan-caspase inhibitor z-VAD-Fmk in the cells (Fig. 3F). These results demonstrate that MG2A induced caspase-dependent apoptosis. However, the addition of caspase inhibitor could not rescue all the cells, suggesting that the peptide-mediated necrosis could not be excluded.
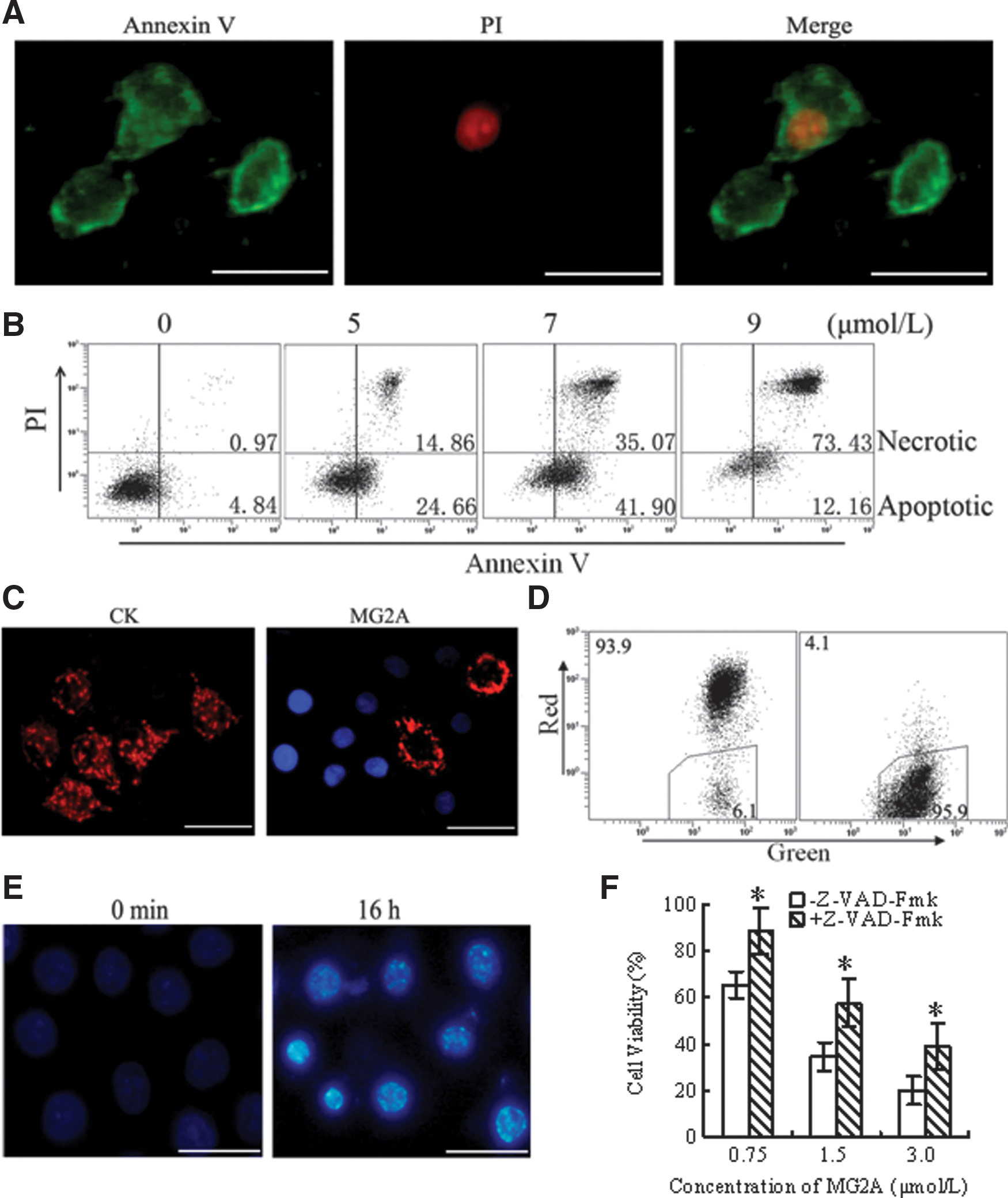
MG2A induces caspase-dependent apoptosis.
Binding to CS contributes to the selective cytotoxicity of MG2A to tumor cells
Antp can bind the CS that is overexpressed on the surface of tumor cells. 21,23 –25 Accordingly, it is possible for Antp to bind CS and mediate the selectively cytotoxicity of MG2A to tumor cells. To prove this hypothesis, we first examined and compared the amount of surface CS on tumor and normal cells. We found that in tumor cells, including HeLa, A549, and A375, the positive percentage of CS was 83.98%, 92.25%, and 99.46%, respectively; whereas in normal cells, including VeroE6 and HK-2, it was only 6.46% and 5.91% (Fig. 4A). These data indicate that CS is overexpressed on the surface of these tumor cells. We also investigated the inhibitory effect of exogenous CS and HA on the cytotoxicity of MG2A. We found that preincubation with 4 μM of MG2A and CS can inhibit 60% of cell death in HeLa cells (100 μg/mL of CS) and A549 cells (10 μg/mL of CS). However, preincubation with exogenous sodium HA had no effect on the cytotoxic activity of MG2A (Fig. 4B). These results suggest that the binding of MG2A to overexpressed CS on the surface of tumor cells contributes to its selective cytotoxicity.

Binding to the overexpressed chondroitin sulfate (CS) on tumor cell surface contributes to the selective cytotoxicity of MG2A to tumor cells.
MG2A suppresses tumor growth in vivo
The in vivo activity of MG2A was tested against HeLa cells transplanted subcutaneously in the right flank of BALB/c (nu/nu) mice. At the onset of a palpable tumor (approximate volume of 50 mm3), 15 mg/kg of MG2A was intraperitoneally injected into mice daily for a total of 5 days. The mice in the control group were injected with the equivalent volume of PBS. Rapid tumor growth in mice from the PBS group was observed as early as 7 days after the injection. However, tumor growth in the MG2A group slowed over the course of the experiment. Beginning on day 14 after injection, the average tumor volume in the MG2A group was significantly different (p<0.05) from the tumor volume in the PBS group. At the end of the experiment, the mean tumor volume in the MG2A group was 3204±882 mm3 and 5127±592 mm3 in the PBS group (Fig. 5A).
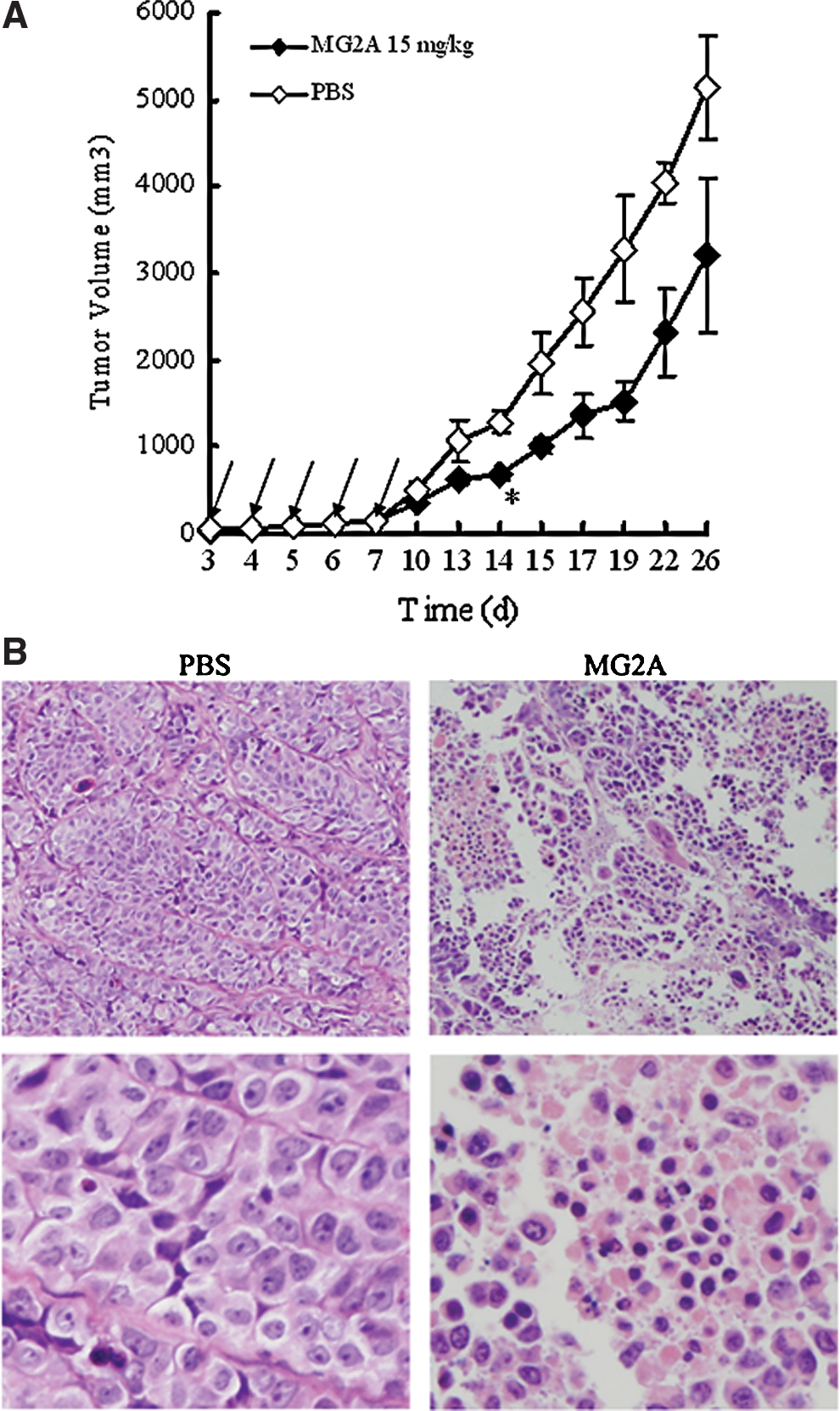
MG2A suppresses tumor growth in vivo.
Furthermore, a histological examination demonstrated that intratumorally injected MG2A caused extensive necrosis in tumor cells. As shown in Figure 5B, H&E staining shows a large number of cells with condensed and fragmented nucleus in tumor tissues at MG2A-injected sites when compared with PBS-injected sites. The results indicate that MG2A can inhibit tumor growth in vivo.
Discussion
In this study, we constructed a fusion peptide, MG2A, by conjugating MG2 to the N-terminus of Antp. We found that the IC50 of MG2A in tumor cells was at least 30 times lower than that of MG2. This change indicates that conjugation to Antp significantly enhanced the cytotoxicity of MG2 in tumor cells. Moreover, the IC50 of MG2A in normal cells was three to five times higher than that in tumor cells. In addition, it was found that the cytotoxicity of MG2A was inhibited by addition of exogenous CS, indicating that conjugated Antp enhanced the cytotoxicity of MG2 in tumor cells through the binding of CS.
CAPs are a novel strategy for the development of alternative cancer therapies as a result of their reduced induction of drug resistant tumor cells. 2 Tumor cell membranes contain negatively charged molecules, while CAPs are positively charged. The electrostatic interaction with the negatively charged molecules on the membranes enable CAPs to bind to tumor cell membranes and exert their antitumor activity. 1 We hypothesized that the cytotoxicity of MG2 in tumor cells only at high concentrations resulted from its poor membrane binding and cell entry due to weak electrostatic interactions. Accordingly, raising the surface positive charge of MG2 by altering its amino-acid composition can enhance binding affinity and antitumor activity. 13,14 In addition, the efficiency of MG2 delivery to tumor cells can also be increased through the use of some vectors. 15,16 CPPs are able to mediate the cell entry of a conjugated cargo across the cellular plasma membrane. 17,18 One of the most frequently applied CPPs, Antp, was found to be able to bind to overexpressed CS on the surface of tumor cells. 19,20 Therefore, conjugation of Antp to MG2 may also enhance the cytotoxicity of MG2 to tumor cells.
Here, we constructed the fusion peptide MG2A by coupling MG2 with Antp. As expected, MG2A showed greater antitumor activity when compared with unconjugated MG2. The IC50 of MG2A for tumor cells was at least 30 times lower than that of unconjugated MG2 (Fig. 1B). Further investigation showed that MG2A was selectively cytotoxic to tumor cells. The IC50 values of MG2A in tumor cells were approximately three to five times lower than in normal cells (Fig. 2A). Moreover, we found that the CS expression was approximately 15 times higher in tumor cells than in normal cells (Fig. 4A), and addition of exogenous CS can significantly reduce the cytotoxicity of MG2A in tumor cells (Fig. 4B). These results demonstrate that conjugation to Antp enhanced the cytotoxicity of MG2 in tumor cells, and binding of Antp to overexpressed CS on the cell surface might contribute to the selectivity of MG2A. These results are consistent with other Antp-mediated mitochondria-disrupting peptides described in our previous work. 21 Consequently, Antp might be a drug carrier candidate for CAP-targeted cancer therapy.
Similar to other CAPs, MG2 was reported to form pores in the membrane of solid and hematological tumor cells, 8,26 suggesting that MG2 might induce necrotic cell death by disrupting the cell membrane. In addition, MG2 have been found to disrupt the membranes of isolated rat liver mitochondria. 27 MG2 can trigger caspase-3 activation and cytochrome c release in HL-60 cells. 28 These data suggested that MG2 might induce apoptosis in tumor cells. In this experiment, necrotic cells (Annexin V+/PI+) were detected in MG2A-treated HeLa cells by a fluorescence microscope (Fig. 3A) and FACS analysis (Fig. 3B). These data suggest that MG2A induced necrotic cell death by a membrane-lytic reaction. In addition, we observed many apoptotic cells (Annexin V+/PI−) using microscopy (Fig. 3A) and FACS analysis (Fig. 3B). The loss of mitochondrial membrane potential (Fig. 3C, D), nuclei condensation, and DNA fragmentation (Fig. 3E) were detected. Moreover, the involvement of caspase in MG2A-induced cell death was verified by using the pan-caspase inhibitor (Fig. 3F).These results demonstrate that MG2A also induced caspase-dependent apoptosis in tumor cells. Therefore, it is possible for MG2A to induce cell death by both necrotic and apoptotic pathways.
Macromolecular biological active agents, such as genes, peptides, and proteins, are limited in their ability to permeate the plasma membrane. CPPs are known as highly efficient delivery vectors and have attracted considerable attention as a tool to introduce drugs into cells. 17,18,20 However, their delivery of cargo into cells is generally considered nonspecific, because a similar cellular uptake by tumor and normal cells has been observed. 29 Therefore, the nonspecific membrane penetration of both tumor and normal cells has extensively limited the use of CPPs in tumor-targeted therapy. Fortunately, in our previous study, we found that one of the most widely studied CPPs, Antp, has the ability to bind to CS, and that CS is overexpressed on the surface of tested tumor cells. 21 Thus, binding of Antp to overexpressed CS on the tumor cell surface may add selectivity to the delivery of cargo into tumor cells by Antp. In our experiments, we observed selective cytotoxicity of MG2A to tumor cells. More than 80% of tumor cells were dead after treatment with 5 μM of MG2A, while 80% of normal cells were still alive (Fig. 2A). However, when the concentration was increased to 20 μM, MG2A induced more than 90% cell death in normal cells. Therefore, the CS-mediated enhanced internalization of MG2A to tumor cells remained hindered by the insufficient selectivity in tumor targeting. Ideally, improved selective tumor targeting is needed for further application of CPPs in tumor-targeted therapy. Many recent studies have made some progress in improving the selectivity of CPPs. 30 –32 Strategies described in these studies might be used for selectivity improvement of MG2A peptide.
In conclusion, this study demonstrated that conjugation of MG2 with Antp can improve the cytotoxic activity of MG2A toward tumor cells. Moreover, MG2A can selectively induce apoptotic cell death in tumor cells. In addition, CS overexpression on the tumor cell surface and inhibition of the cytotoxicity of MG2A by exogenous CS suggest the increased cytotoxicity of MG2A for tumor cells resulting from the increased membrane binding and cell entry. In vivo, MG2A also displayed tumor suppression activity. These data suggest that conjugation with Antp might improve the antitumor activity of the membrane-active CAPs and might be an alternative for cancer-targeted therapy.
Footnotes
Acknowledgments
This work was supported by the Natural Science Fund of China (81072566) and China Postdoctoral Science Foundation funded project (201104645, 20100481392).
Disclosure Statement
The authors declare that there is no conflict of interest.