
Abstract
Endostar was approved for the treatment of cancer as an antiangiogenic agent with limited therapeutic effects used alone in cancer treatment. Tetrandrine (TET) has a variety of nontumor-related effects and anticancer effects, including antiangiogenic effects. This study was designed to explore the interaction of Endostar and TET. Antiproliferative effects of TET combined with Endostar on human umbilical vein endothelial cells (HUVECs) and the human colon cancer cell line LoVo were evaluated by the 3-(4,5-dimethylthiazol-2-y)-2,5-diphenylterazolium bromide (MTT) assays. The effects on HUVEC migration and tube formation of TET plus Endostar were observed by the transwell test and tube formation assay. Refer to the mechanisms of the cell proliferation inhibition effect caused by the two drugs: apoptosis assay and cell cycle analysis of HUVECs were analyzed by Annexin V-fluorescein isothiocyanate (FITC) and propidium iodide (PI) staining, followed by flow cytometry. The combination of TET and Endostar had a synergistic effect on the antiproliferation of HUVECs and LoVo cells. Further, all these antiangiogenic effects such as inhibition of cell migration, suppression of tube formation, induction of cell apoptosis, and cell cycle arrest were enhanced when HUVECs were treated with TET combined with Endostar. Thus, we identified that there was a synergistic antiangiogenic effect in vitro when combining TET with Endostar.
Introduction
Antiangiogenic therapy, directly targeting tumor vascular endothelial cells, is now recognized as the fourth cancer treatment in addition to surgery, chemotherapy, and radiotherapy. 1 As of yet, several conclusive antiangiogenic drugs such as Avastin, Sorafenib, and Sunitinib have been approved by the U.S. Food and Drug Administration. Endostar, a novel recombinant human endostatin, was approved by the Chinese State Food and Drug Administration in 2005 for the treatment of nonsmall-cell lung cancer as an antiangiogenic agent. The mechanisms of antiangiogenic efficacy for Endostar have been proved by previous reports both in vitro and in vivo. 2 –4 Endostar is well tolerated with minor toxicity in clinical use, but clinical application of Endostar revealed a weak antitumor activity when used alone in cancer treatment. 5 –7 Thus, Endostar is always applied together with other chemotherapeutics, or it is used in the second or later line of cancer treatment for patients with poor status.
Tetrandrine (TET) is a bisbenzyl isoquinoline alkaloid isolated from the root of Stephania tetrandra S. Moore, which has a variety of effects and has been widely used in the traditional Chinese medicine for several decades. Recent investigations have reported that TET exhibited anticancer effects, including antiproliferation and apoptosis induction effects, on cancer cells, 8 –10 reversing the multidrug resistance of cancer, 11 antiangiogenic effects 12 –15 and synergistic effects with chemotherapeutics. 16 –18 For antiangiogenesis, TET was reported to suppress vascular endothelial growth factor (VEGF) expression in glioma cells and induce cytotoxic effects on human umbilical vein endothelial cells (HUVECs). An in vivo Matrigel plug assay revealed that TET could suppress the angiogenesis that was induced by basic fibroblast growth factor (b-FGF), and TET could also suppress tumor growth and angiogenesis of gliomas in rats. 15 However, few were known about the interaction effect of TET and Endostar on the antiangiogenic effect. In this study, we studied the antiangiogenic activity of TET alone and combined it with Endostar on the model of HUVECs.
Materials and Methods
Cell culture and chemicals
HUVECs were isolated from freshly obtained human umbilical cords as described previously. 19 The collected HUVECs were maintained in the endothelial cell growth medium (EGM-2; Cambrex Corporation). The EGM-2 is composed with an endothelial cell basic medium, endothelial cell growth supplement, and 10% fetal bovine serum (FBS). LoVo cells were grown in Roswell Park Memorial Institute (RPMI)-1640 supplemented with 10% FBS (Gibco Corporation). Both of the cell lines were grown in a 37°C humidified growth chamber containing 5% CO2 in air. TET (molecular formula C38H42N2O6) was obtained as a powder with a purity of >98% from Jiangxi Yibo Pharmaceutical Development Company. It was dissolved in 0.1 N hydrochloric acid and diluted with distilled water (the pH value was adjusted to 6.6–6.8 with 1 N sodium hydroxide).
Cell proliferation assay
The HUVECs and LoVo cells were seeded at 4000 cells/well and 2000 cells/well in 96-well plat-bottom plates, respectively. A complete medium containing different concentrations of TET and/or Endostar was added to the wells after the cells were seeded for 24 hours. The viability of cells was determined by the 3-(4,5-dimethylthiazol-2-y)-2,5-diphenylterazolium bromide (MTT) (Gibco Corporation) assay after another 48 hours of incubation. The absorbance was read at 490/630 nm. The half maximal inhibitory concentration (IC50) was defined as the concentration that required for 50% inhibition of cell growth.
Cell migration assay
Before the cell migration assay, we designed an MTT assay to evaluate the HUVEC viability after drug incubation for 12 hours. Then, the migration assay was performed using transwell-permeable supports (Corning Incorporated), which consist of upper and lower compartments separated by a porous membrane (8 μm) as described previously. 20 HUVECs was precultured with a low-serum medium (0.5% FBS) for 12 hours to induce a quiescent state. Transwell inserts were coated with 100 μL fibronectin (10 μg/mL), and then EGM-2 supplemented with 10% FBS (0.6 mL) was added to the lower well. After that, HUVECs were seeded (2×105 cells/well, 50 μL) in the upper wells with the low-serum (0.5% FBS) medium, and the medium was added with different concentrations of TET or TET combined with Endostar. The cells incubated with the low-serum (0.5% FBS) medium were used as the negative control. After additional 12-hour incubation, the medium was removed; the inserts were washed three times with phosphate-buffered saline; and the cells adhering to the filter were fixed with 10% methanol for 30 minutes and stained with 1% crystal violet for 20 minutes. Cells on the lower side of the porous membrane were counted in five separate and random microscopic fields, and we take the means±standard deviations (SDs) as the migration number.
Tube formation assay of HUVECs
Formed tubes of endothelial cells sandwiched between fibrinogen gels 21 were observed to evaluate the differentiation ability of HUVECs. The fibrinogen solution (250 μL, 1.0 mg/mL) was placed into each well of a 24-well culture plate, and 5 μL of human thrombin (12.5 U/mL) was added to the fibrinogen solution. After 5 minutes at 37°C in the incubator, the gels were polymerized. HUVECs (1×105 cells/mL, 250 μL) were seeded onto each fibrinogen gel. After about 1-hour incubation at 37°C, the apical surface of the fibrinogen gel polymerized. EGM-2 (1 mL) was added on the top of each fibrinogen gel, and was replaced with the fresh medium every 24 hours in the 72-hour incubation. The medium containing TET alone or TET plus Endostar was added on the top of the fibrinogen gels. Moreover, wells receiving the fresh medium were used as the control. After 24-hour treatment, lengths of tubes were measured in five separate and random microscopic fields, and the means±SDs were taken as the migration number.
Cell apoptosis assay and cell cycle analysis
The cell apoptosis rates were measured according to the manufacturer's instructions (BioVision Incorporated). Briefly, cells were plated in a 60-mm culture dish and treated with drugs for 48 hours and were collected and tested by flow cytometry with Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) staining. Cells distributed in a different phase of cell cycle were quantified by PI staining, and the percentage of cells in different phases was determined using Modfit LT software (BD Biosciences).
Statistical analysis
The Webb coefficient method was applied to determine the interaction between the two drugs. 22 The predicted value (c) was calculated according to the equation c=a×b/100, where a and b indicate survival values obtained when cells were treated with single agents. Synergism in drug interaction was indicated by observed survival of ≤70% of the predicted survival. Data were analyzed using SPSS 13.0 software. Student's t analysis was used to compare the difference between two groups. All values are calculated as means±SDs. The accepted level of significance was P<0.05.
Results
Inhibitory effect of TET and/or Endostar on cell proliferation
HUVECs and LoVo cells were treated with different doses of TET alone and in combination with Endostar as described above. As shown in Figure 1A, TET (2, 4, and 8 μg/mL) inhibited the proliferation of LoVo cells and HUVECs in a dose-dependent manner, with the IC50s of 59.31 μg/mL for LoVo cells and 4.27 μg/mL for HUVECs. The IC50 on endothelial cells is lower than that needed for a toxicity on LoVo cells (P=0.024). The differential toxicity of TET on HUVECs and LoVo indicated that endothelial cells were the main targets of TET. However, Endostar alone only showed a weak inhibitory effect both on tumor cells and HUVECs (data not shown). According to the assessment of the Webb coefficient method described above, the combination of 2, 4, and 8 μg/mL TET with 10 μg/mL Endostar showed a synergistic inhibitory activity, with the IC50s of 8.96 μg/mL for LoVo cells and 2.69 μg/mL for HUVECs (P=0.028) (Fig. 1B).
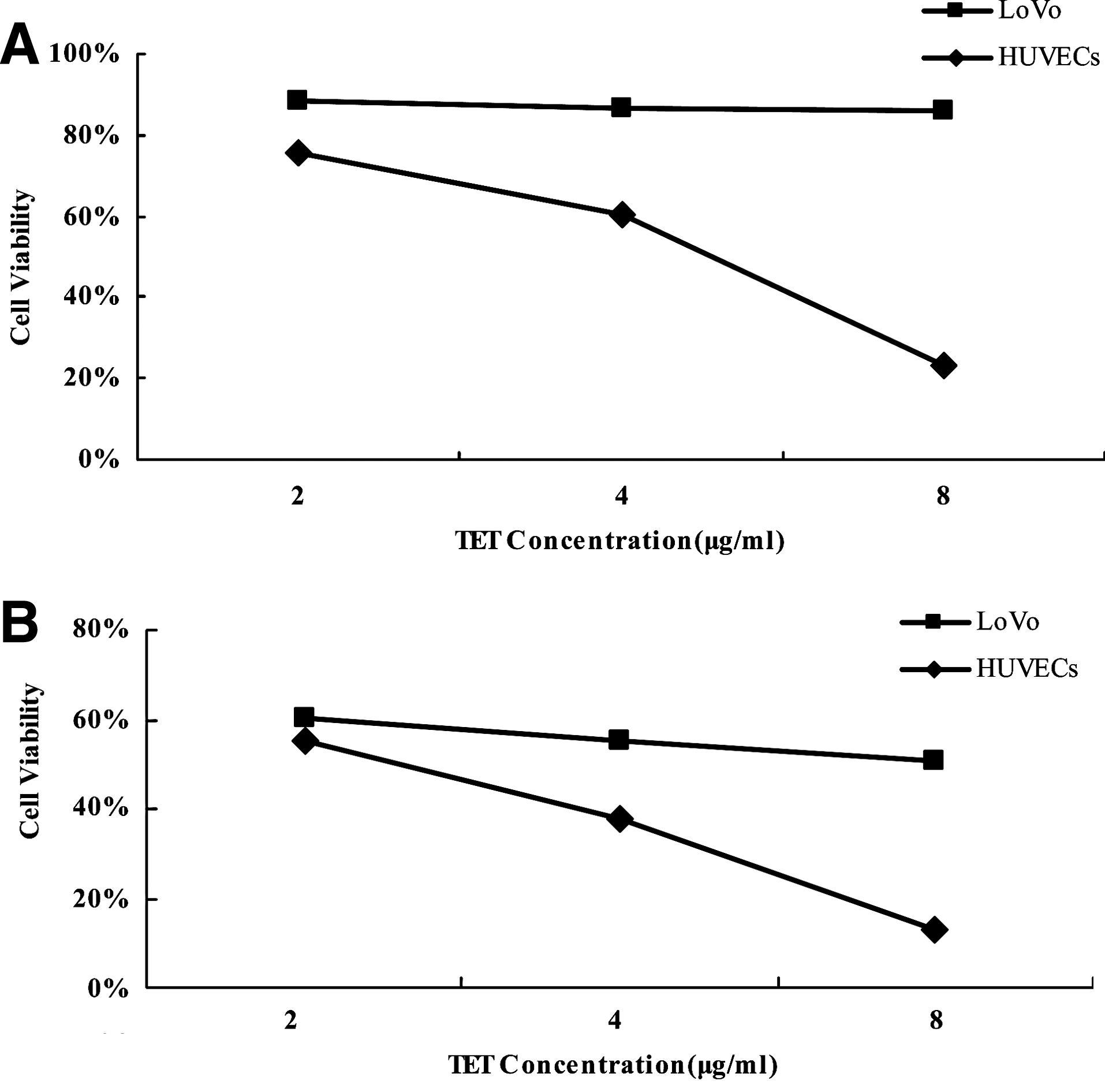
Antiproliferation assay in vitro of human umbilical vein endothelial cells (HUVECs) and LoVo cells after treatment with
Inhibitory effect of HUVEC migration by TET and/or Endostar
HUVECs that were treated with 2, 4, and 8 μg/mL TET for 12 hours demonstrated the cell viability as 99%, 96.3%, and 90.9%, respectively. In the cell migration assay, TET significantly decreased the number of migrating cells at the concentrations of 2, 4, and 8 μg/mL (P<0.001) compared with the control (Fig. 2). The relative high survival rate of HUVECs treated with TET for 12 hours indicated that the cytotoxicity of TET almost had no effect on the number of migrated HUVECs, whereas there was no significant difference between 10 μg/mL Endostar-treated group and the control group (P=0.831). According to the Webb coefficient, 2, 4, and 8 μg/mL TET combined with 10 μg/mL Endostar caused a dose-dependent decrease in migrating cells compared with the corresponding groups treated with TET alone (P=0.001, 0.003, and 0.003, respectively).
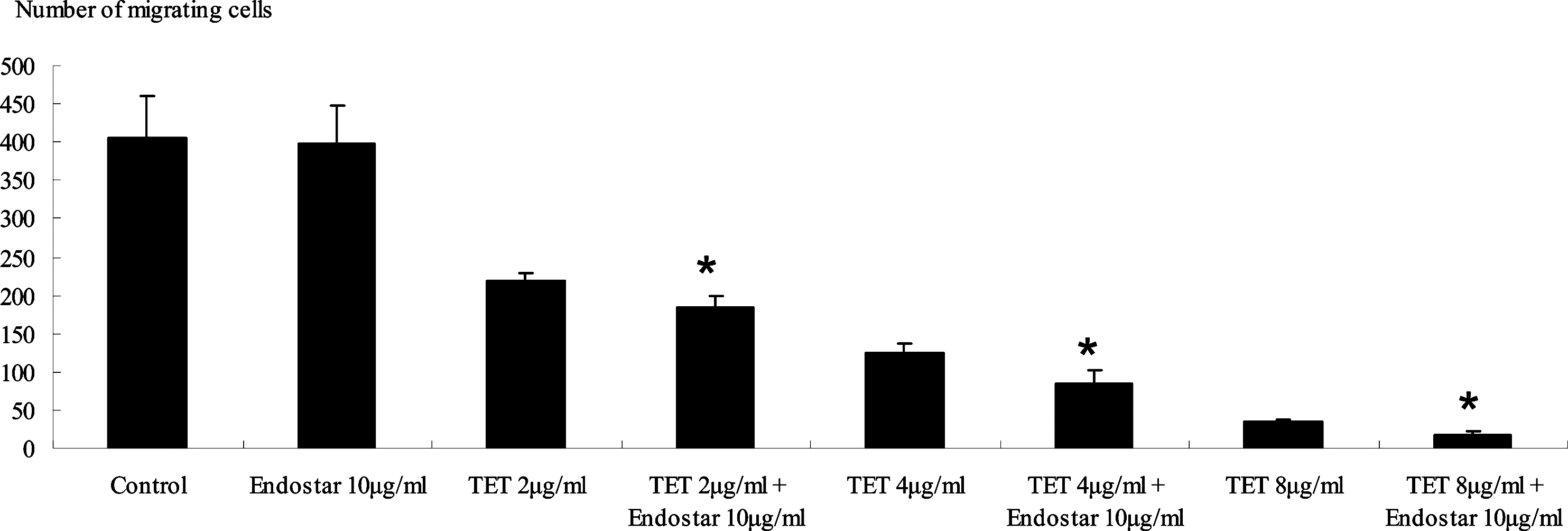
The effect of inhibiting HUVEC migration was measured by the number of migration through the transwell-permeable supports after treated with different groups. Treatment without any drug served as a control. *Significant differences versus the corresponding TET treated alone, P<0.01.
TET and/or Endostar inhibits tube formation of HUVECs
The ability of the antiangiogenic activity of TET alone or TET combined with Endostar was examined in a tube formation assay. We performed dose–response experiments to assess the potency of the antiangiogenic effects of TET (2, 4, and 8 μg/mL) and/or 10 μg/mL Endostar (Fig. 3). TET and Endostar alone significantly reduced the length of the tubes. Moreover, according to the Webb coefficient, TET (2, 4, and 8 μg/mL) combined with 10 μg/mL Endostar led to a synergistic inhibitory effect of HUVEC tube formation comparing to the corresponding group with TET-treated alone (P<0.001, P<0.001, and P=0.002, respectively).

Tube formation of HUVECs was inhibited by different groups. Results represented as the length of tubes. HUVECs treated without any drug served as a negative control. *Significant differences versus the corresponding TET treated alone, P<0.01; and # significant differences versus the control, P<0.01.
TET and/or Endostar induces apoptosis on HUVECs
To investigate the mechanisms of the antiproliferative effects of TET and/or Endostar in HUVECs, we examined the apoptosis rates of HUVECs induced by the two drugs. HUVECs in culture flasks were treated with 8 μg/mL TET alone, 10 μg/mL Endostar, or 8 μg/mL TET combined with 10 μg/mL Endostar. After 48 hours, the cells were collected and determined by double staining with Annexin V-FITC/PI, followed by flow cytometric analysis. As shown in Figure 4, the apoptotic rate in 8 μg/mL TET-treated group was higher (20.8%) than the control (7.1%), and the combination of both drugs showed more prominent activity (60.4%), whereas the apoptotic rate in the Endostar-treated-alone group was 18.8%.

The apoptotic ratio induced by TET and/or combined with Endostar on HUVECs after treatment for 24 hours as measured by flow cytometric analysis. HUVEC treatment without any drug served as a negative control.
TET and/or Endostar change cell cycle on HUVECs
Flow cytometric analysis was performed on HUVECs after treated with different treatments to assess the cell cycle status. As shown in Figure 5, cell cycle analysis showed a noticeable increase in the G0/G1 phase and a progressive decrease in the G2/M and S phase when cells were treated with 8 μg/mL TET alone, whereas 10 μg/mL Endostar alone showed a minor increase effect in the G2/M phase and the S phase. However, there was no synergistic effect on cell cycle when HUVECs were treated with the combination of the two drugs.
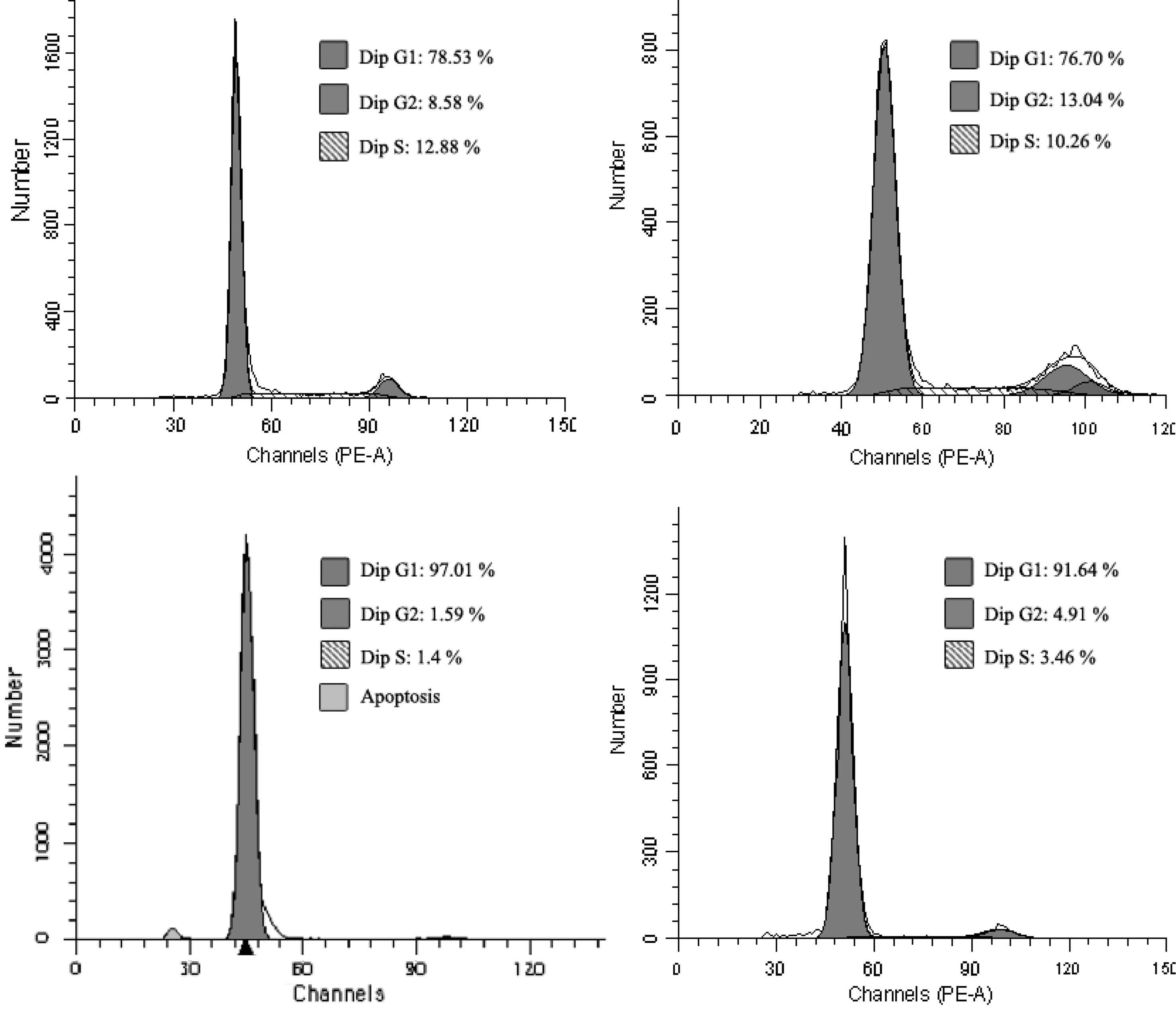
Cell cycle analysis of HUVECs treated with TET and/or combined with Endostar for 24 hours. HUVEC treatment without any drug served as a negative control.
Discussion and Conclusions
Traditional chemotherapy as a major treatment for the malignant tumor has two major problems at all times: acquired drug resistance and nonselective toxicity. Antiangiogenesis therapy avoids drug resistance and toxicity of common therapeutics due to its special mechanism. However, antiangiogenic therapy-related side effects such as severe hemorrhage may also limit its clinical application. Combined treatment could work on different pathways and reduce the dose of each drug, which can overcome the shortcomings of a single treatment. Actually, many reports have shown that Endostar has more apparent antitumor and antiangiogenic effects when combined with another drug than the single drug. 23 –26 In this study, we have shown for the first time that TET combined with Endostar inhibits the proliferation, migration, and tube formation of HUVECs, thus demonstrating its antiangiogenic activity in vitro. Further, we explored the possible mechanism of cytotoxicity on HUVECs using the cell apoptosis assay and cell cycle analysis.
The proliferation of vascular endothelial cells is a crucial fundament for the formation of the vessels. In the present study, we found that TET specifically inhibited proliferation of HUVECs more efficiently than on LoVo cells, indicating an obvious different cytotoxicity on HUVECs and tumor cells. Endostar had no significant inhibitory effect on HUVEC proliferation. Interestingly, we investigated that there was a synergistic antiproliferative effect when cells treated with TET combined with Endostar, and the possible mechanism may due to the synergism in the induction of apoptosis.
Apoptosis and cell cycle examination provided a primary understanding of the mechanism on the HUVEC proliferation inhibition. Previous studies reported that TET induced apoptosis in human hepatoblastoma cells via caspase-dependent Bid cleavage and cytochrome c release, 27 and another report demonstrated that TET-induced apoptosis is mediated by activation of caspases and PKC-delta in U937 cells. 28 Recently, it has been reported that TET-induced apoptosis in rat primary hepatocytes is initiated from the mitochondria: caspases and the endonuclease G pathway. 29 Although no significant proliferation inhibitive effect was observed for Endostar on HUVECs, a slight apoptosis was induced by Endostar in our experiments. Reports showed that Endostar could induce apoptotic effects on HUVECs through activation of caspase-3. 4 It is high likely that TET combined with Endostar induces apoptosis in HUVECs via a caspase-dependent pathway. However, the detailed molecular mechanism needs to be verified by further experiments.
In the current study, HUVECs treated with 8 μg/mL TET alone or TET in combination with Endostar induced the cell cycle arrest in the G0/G1 phase, but Endostar only induced a minor increase in the G2/M phase. Combination of TET and Endostar showed the same trend as in the TET group, which indicated that TET exerted a more important effect on the cell cycle. On the other hand, cell cycle arrest may not be the mechanism of synergism of TET and Endostar as the antiangiogenesis effect.
Endothelial cell migration and tube formation are the two crucial steps in the tumor-associated angiogenesis. Cell migration was inhibited in our study after 12 hours of treatment when no significant proliferation inhibition was observed. It suggested that the migration inhibition effect may be not via cytotoxicity. Tube formation assay provides the most complete assessment in vitro of endothelial cell function as a semiquantitative method, and inhibitory effects of tube formation are all observed when cells were treated by different concentrations of TET and Endostar. When TET combined Endostar, a synergistic inhibition of cell migration and tube formation of HUVECs were observed.
The activated, proliferative, invasive endothelial cells that form cords and tubes were thought to recapitulate early capillary development. TET alone or combined with Endostar may have an antiangiogenic effect on HUVECs in terms of cell proliferation, migration, and tube formation. Thus, TET could be used in antitumor treatment as an antiangiogenic agent alone or combined with Endostar. However, a systematic study that contains in vivo examination and mechanism exploration should be carried out. Moreover, well-designed clinical trials are also in plan to confirm the role of TET alone and combined with Endostar in the antiangiogenic treatment for cancer patients.
Footnotes
Acknowledgments
This work was supported in part by the Key Foundation of Health Department of Nanjing, China (ZKX09017), and the Foundation of Health Department of Jiangsu Province, China (H201035).
Disclosure Statement
None declared.