
Abstract
Colorectal cancer is an aggressive malignancy with a high mortality rate; however, effective therapies are currently lacking. Cancer-targeting gene–virotherapy (CTGVT) has been proposed to be a promising strategy for cancer therapy. The purpose of this study was to investigate the antitumor activity of the oncolytic adenovirus harboring Lipocalin-2 (ZD55-Lipocalin-2, an example of CTGVT) in colorectal cancer. ZD55-Lipocalin-2 was generated by deleting E1B55-KD and inserting the Lipocalin-2 gene. Its cytopathic effects and cell growth inhibition were detected in vitro, and antitumor effects were examined in a nude mouse model of human colorectal cancer xenografts. Results showed that ZD55-Lipocalin-2 significantly inhibited the colorectal cancer growth by selective cytolysis, inducing apoptosis and decreasing the microvessel density in tumors. The anticancer potential of ZD55-Lipocalin-2 showed stronger than that of the isolated Lipocalin-2 gene therapy or isolated ZD55 oncolytic adenovirus therapy. ZD55-Lipocalin-2 may serve as a potential anticancer agent for colorectal cancer treatment.
Introduction
Colorectal cancer is the third most commonly diagnosed malignancy among humans. About 1 million new colorectal cancer cases are diagnosed worldwide each year, and ∼492,000 1,2 of these result in death. Especially, in the more developed regions in China, colorectal cancer has become more common in recent years and has become a substantial cancer burden. 3 Unfortunately, no ideal therapeutic options are currently available. Radical surgery is the main treatment for this disease, but this is not an option in about 50% of cases. Moreover, the incidence of metastasis and postoperative recurrence is high, so it would be valuable to develop a novel, more effective therapeutic approach for treatment of this aggressive disease.
Oncolytic virus therapy represents an attractive platform for the development of novel cancer treatments. An increasing number of oncolytic viruses have been demonstrated to kill cancer cells, and the efficacy and safety of oncolytic adenoviruses have been confirmed in experimental and clinical trials. 4,5 Cancer-targeting gene–virotherapy (CTGVT) based on oncolytic viruses represents a promising new approach in cancer therapy and has shown synthesized antitumor. In CTGVT, the therapeutic anticancer gene is inserted into conditionally replicative adenoviruses. The resulting antitumor effect is much greater than that of a single oncolytic adenovirus, but its safety is similar to that of its corresponding oncolytic adenovirus.
The replication of oncolytic adenoviruses that express therapeutic anticancer genes can be used to limit cancer growth. The release of viral progeny from infected tumor cells provides a potential to amplify the oncolytic effects and encourage its lateral spread through solid tumors. At the same time, the antitumor function of the gene within the oncolytic adenovirus will also be amplified dramatically. Certainly, CTGVT has shown a great potential in the field of cancer therapeutics, and it has even been shown to completely eliminate xenografts in nude mouse models. 6
Lipocalin-2, known as neutrophil gelatinase-associated lipocalin, is a small 25-kDa protein. 7 The principal characteristic of Lipocalin-2 is its ability to capture iron particles and the siderophores transporting them to the inner cell after interacting with its receptor (24p3R). Some studies have demonstrated that Lipocalin-2 has antitumor and antimetastatic effects in some neoplasias, such as cancer from the colon, ovary, and pancreas, due to its inhibition of the proneoplastic protein hypoxia-inducible factor 1, FA-Kinase phosphorylation, and vascular endothelial growth factor (VEGF) synthesis. 8 –12 Apoptosis induced by Lipocalin-2 in cells was also reported, 13,14 but not in colorectal cancer cells. Moreover, there is no direct evidence showing therapeutic effects of Lipocalin-2 in colorectal cancer. Thus, to investigate whether the combination of an oncolytic adenovirus and Lipocalin-2 will exhibit the ability to kill colorectal cancer cells, the novel oncolytic adenovirus ZD55-Lipocalin-2 was established, where the E1B55 gene was deleted, and the Lipocalin-2 expression cassette was inserted into ZD55. 15 Its antitumor efficacy was explored both in vitro and in vivo. Our results showed that ZD55-Lipocalin-2 could effectively induce apoptosis of tumor cells both in vitro and in vivo, and suggested that it might be a potential therapeutic approach for the treatment of colorectal cancer.
Materials and Methods
Plasmids and recombinant adenovirus reconstruction
The Lipocalin-2 gene was amplified by polymerase chain reaction (PCR) from a cDNA library (Clontech Corporation). It was cloned into plasmid pcDNA3.1/Myc-his(+)A. The Lipocalin-2 coding sequence was obtained from pcDNA3.1/Myc-his(+)A by digestion with Hind III and Xho I. The sequence was then subcloned into plasmid pCA13 to form pCA13-Lipocalin-2. pZD55-Lipocalin-2 was constructed by inserting the whole Lipocalin-2 expression cassette digested from pCA13-Lipocalin-2 into the corresponding site of pZD55. All recombinant plasmids were confirmed by restrictive enzyme digestion, PCR, and DNA sequencing. The replication-defective adenovirus Ad-Lipocalin-2 and oncolytic adenovirus ZD55-Lipocalin-2 were produced in HEK293 cells by homologous recombinations between pCA13-Lipocalin-2 or pZD55-Lipocalin-2 and the adenovirus-packaging plasmid pBHGE3 (Microbix Biosystems), respectively. Viruses were purified by cesium chloride-gradient ultracentrifugation. Virus titers were measured by a plaque assay on HEK293 cells. ZD55-EGFP, ZD55-IL-24, and ZD55 are all E1B55-kD-deleted oncolytic adenoviruses. ZD55-EGFP and ZD55-IL-24, based on ZD55, were generated by inserting the EGFP gene and the IL-24 gene, respectively. ZD55-EGFP, ZD55-IL-24, and ZD55 viruses were kept in our laboratory. Lipocalin-2 primers were as follows: sense-5′ CTATAAGCTTATCATGGCCCTAGGTCTCCT 3′ and antisense-5′ TCTAGAATTCAGCTCCCTCAATGGT GTTC 3′. The PCR program for Lipocalin-2 is as follows: 94°C 5 minutes, 30 cycles (94°C 30 seconds, 53°C 45 seconds, and 72°C 1min), and 72°C 10 minutes.
Cell lines and culture conditions
The human lung fibroblast cell line MRC5 and human colorectal cell lines Sw620, Ht29, and Hct116 were purchased from the Shanghai Cell Collection. The HEK293 cell line was obtained from Microbix Biosystems. Cells were cultured in DMEM supplemented with 10% fetal bovine serum, 4 mM glutamine, 50 units/mL penicillin, and 50 μg/mL streptomycin.
Western blot analysis
Cells were harvested and resuspended in a lysis buffer. After centrifugation at 12,000 g for 15 minutes at 4°C, the supernatant containing the cytoplasmic fraction was used to detect E1A, E1B55, and Lipocalin-2 proteins. Proteins were separated by SDS-PAGE and transferred to polyvinylidene difluoride membranes (Millipore). The membranes were blocked with 5% nonfat dry milk and then incubated with corresponding primary antibodies (E1A and E1B55 from Santa Cruz and Lipocalin-2 from Abcam) and secondary antibodies. An enhanced chemiluminescence detection kit (Pierce Biotechnology) was used for immunodetection.
Cytopathic effect assay
The cytopathic effects (CPEs) were observed by crystal violet staining. After adenovirus infection for 5 days, the culture medium was removed, and the cells were washed twice with phosphate-buffered saline (PBS). Then, 500 μL of 0.5% crystal violet solution was added to each well of a 24-well plate, and cells were incubated for 10 mins before being washed with water.
Cell viability assay
Cytotoxicity was checked by the MTT [3-(4, 5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay. Sw620 cells were plated into a 96-well microplate and infected with various adenoviruses at different multiplicities of infection (MOIs). At different time points, the medium was removed, and a fresh medium containing MTT (0.5 mg/mL) was added to each well. After 4 hours of incubation at 37°C, 100 μL of lysing buffer (50% SDS and 50% 2,4-dinitrofluorobenzene) was added. Absorbance was read at 595 nm with a Bio-Rad microplate reader model.
Tumor xenografts in nude mice
Human colorectal cancer xenografts were established on 5-week-old nude mice (Shanghai Experimental Animal Center, Shanghai, China) by subcutaneous inoculation of 1.5×107 Sw620 cells into the right flank of each mouse. When tumors reached about 200 mm3, the xenograft mice were randomly divided into four groups: ZD55-Lipocalin-2, Ad-Lipocalin-2, ZD55-EGFP, or PBS. Adenoviruses and PBS (5×108 PFU/dose, totally four doses with 2×109 PFU) were injected intratumorally according to the corresponding group. The tumor volume was monitored every 2–3 days using a venire caliper (V=length×width2/2).
Histopathological and immunohistochemical study
Tumor xenografts in nude mice were dissected and fixed in 10% formalin, embedded with paraffin, and then cut into 5-μm-thick sections. All sections were baked, deparaffinized, and heated in a citrate buffer. The sections were incubated with a blocking serum at room temperature for 30 minutes. Immunohistochemistry was performed with anti-VEGF antibody (Santa Cruz). The microvessel density (MVD) was assessed according to Weidner's method. 16 Tissue sections stained without the primary antibody served as negative controls. The slides were also stained with hematoxylin and eosin.
Apoptosis checking by terminal deoxynucleotidyl transferase dUTP nick-end labeling
The terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assay was used to examine apoptosis and carried out according to the manufacturer's procedures using an in situ cell apoptosis detection kit (KGA).
Statistical analysis
Statistical analysis was carried out with SPSS 13.0 for windows. All continuous data were presented as mean±standard deviation. Categorical variables were compared by the chi-square test or Fisher's exact test. The independent-sample t-test or analysis of variance (ANOVA) was used to compare the means of groups. The Kaplan–Meier method was used to estimate survival, and the survival difference was compared by the log-rank test; p-values<0.05 were considered statistically significant.
Results
Verification of the pZD55-Lipocalin-2 plasmid and oncolytic adenovirus ZD55-Lipocalin-2
Adenovirus E1A plays a key role in viral replication. E1A-mediated host cell apoptosis is antagonized by adenovirus E1B-55 kDa. 17 If E1B-55 kDa is deleted, the adenovirus will survive in tumor cells, but not in normal cells. The inserted anticancer genes can be highly expressed in ZD55 vector-deleted E1B-55kDa. 18 Our results showed that 1300-bp gene fragments were obtained from pZD55-Lipocalin-2 digested with BglII single-restriction enzyme (Fig. 1A). About 700-bp fragments were amplified by PCR using Lipocalin-2 primers and pZD55-Lipocalin-2 as a template (Fig. 1B). This information indicates that the Lipocalin-2 gene was successfully cloned into ZD55.
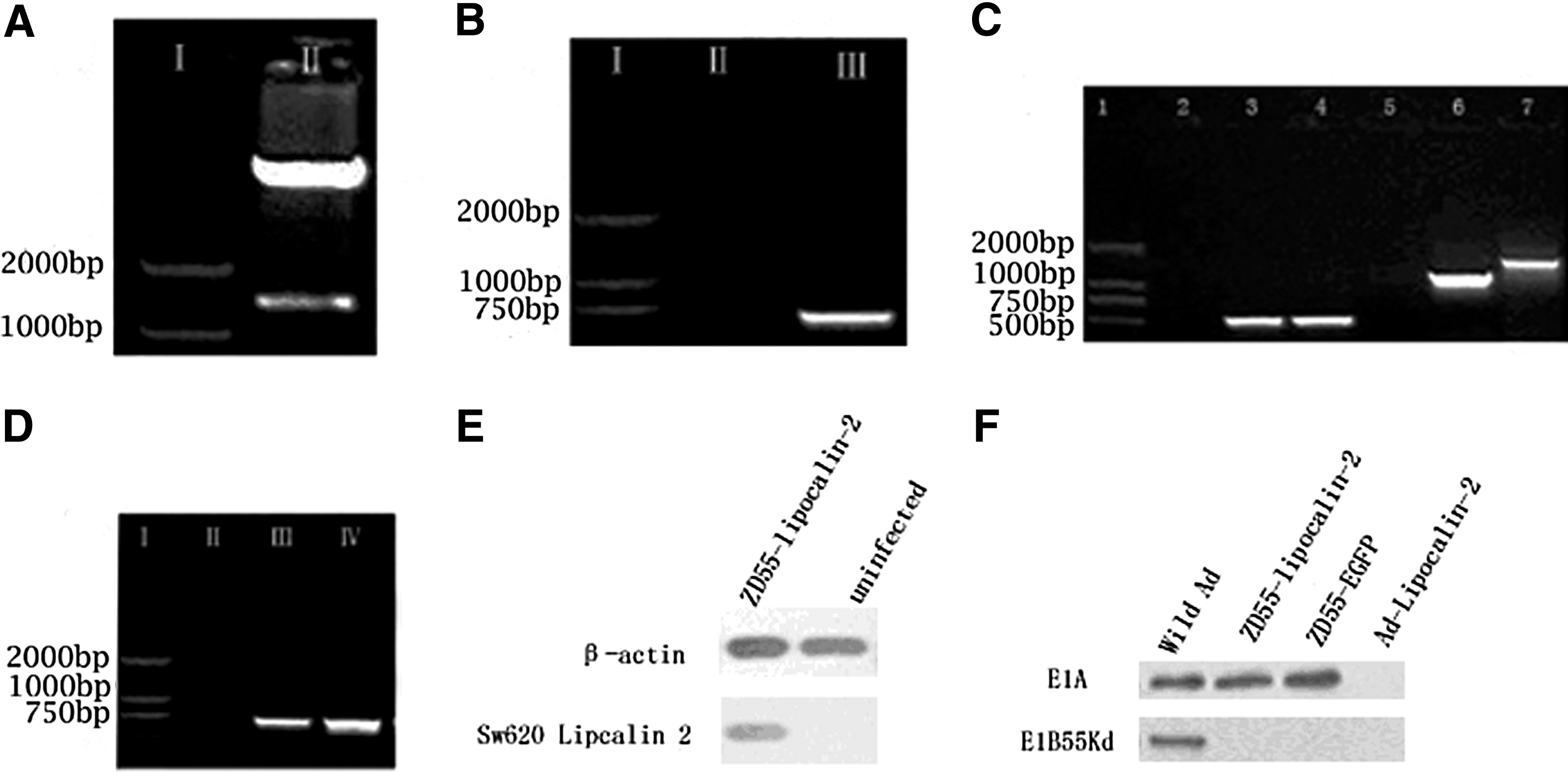
Confirming plasmid pZD55-Lipocalin-2 and adenovirus ZD55-Lipocalin-2.
Wild-type adenovirus contamination is a major issue during ZD55-Lipocalin-2 virus reconstruction. However, the ZD55 sense (5′ AGAGCCCATGGAACCCGAGA 3′) and antisense primers 15 (5′ CATCGTACCTCAGCACCTTCCA 3′) can be used to solve this problem. If ZD55-Lipocalin-2 is contaminated by wild-type adenoviruses, the 1153-bp fragments will be amplified using this pair of primers. In contrast, the 1450-bp fragments are amplified in conditions without wild-type adenovirus contamination. 15 Results from PCR showed that 1450-bp products were detected using the ZD55 sense and ZD55 antisense primers and the ZD55-Lipocalin-2 template (Fig. 1C). Using primers for the Lipocalin-2 gene, Lipocalin-2 was amplified from the ZD55-Lipocalin-2 template (Fig. 1D). Western blot also demonstrated that high expression of Lipocalin-2 protein was detected in Sw620 cells infected by ZD55-Lipocalin-2 (Fig. 1E). ZD55-Lipocalin-2 expressed E1A protein, but failed to express E1B-55 kDa protein. In contrast, wild-type adenovirus expressed both proteins. E1-deleted replication-defective Ad-Lipocalin-2 did not express either of them (Fig. 1F). These results suggest that ZD55-Lipocalin-2 was successfully established.
CPEs and growth inhibition of ZD55-Lipocalin-2
To detect the tumor-specific CPEs of ZD55-Lipocalin-2, two p53-mutated cell lines (Sw620 and Ht29), one wild-type p53 tumor cell line (Hct116) and one wild-type p53 normal fibroblast cell line (MRC5), were infected with ZD55-Lipocalin-2, ZD55-IL-24, a mixture of ZD55-IL-24 and ZD55-Lipocalin-2, and ZD55-EGFP at different MOIs. Significant CPEs were observed in tumor cell lines infected with ZD55-Lipocalin-2. The killing effect of ZD55-Lipocalin-2 was higher than that of ZD55-EGFP in vitro. ZD55-IL-24 acted as a positive control, which showed strong antitumor effects in colorectal cancer. 17 ZD55-Lipocalin-2 had similar killing effects to ZD55-IL-24 on Hct116 cells, but was stronger on Sw620. In addition, the killing effect of ZD55-Lipocalin-2 was stronger in Sw620 or Hct116 than in Ht29 colorectal tumor cells. Moreover, ZD55-Lipocalin-2 caused limited cell death in MRC5 cells (Fig. 2A). The viability of Sw620 cells was significantly decreased after infection with ZD55-Lipocalin-2, whose growth inhibition was both dose- and time-dependent. ZD55-Lipocalin-2 produced the strongest growth inhibition in Sw620 among three agents (Fig. 2B).
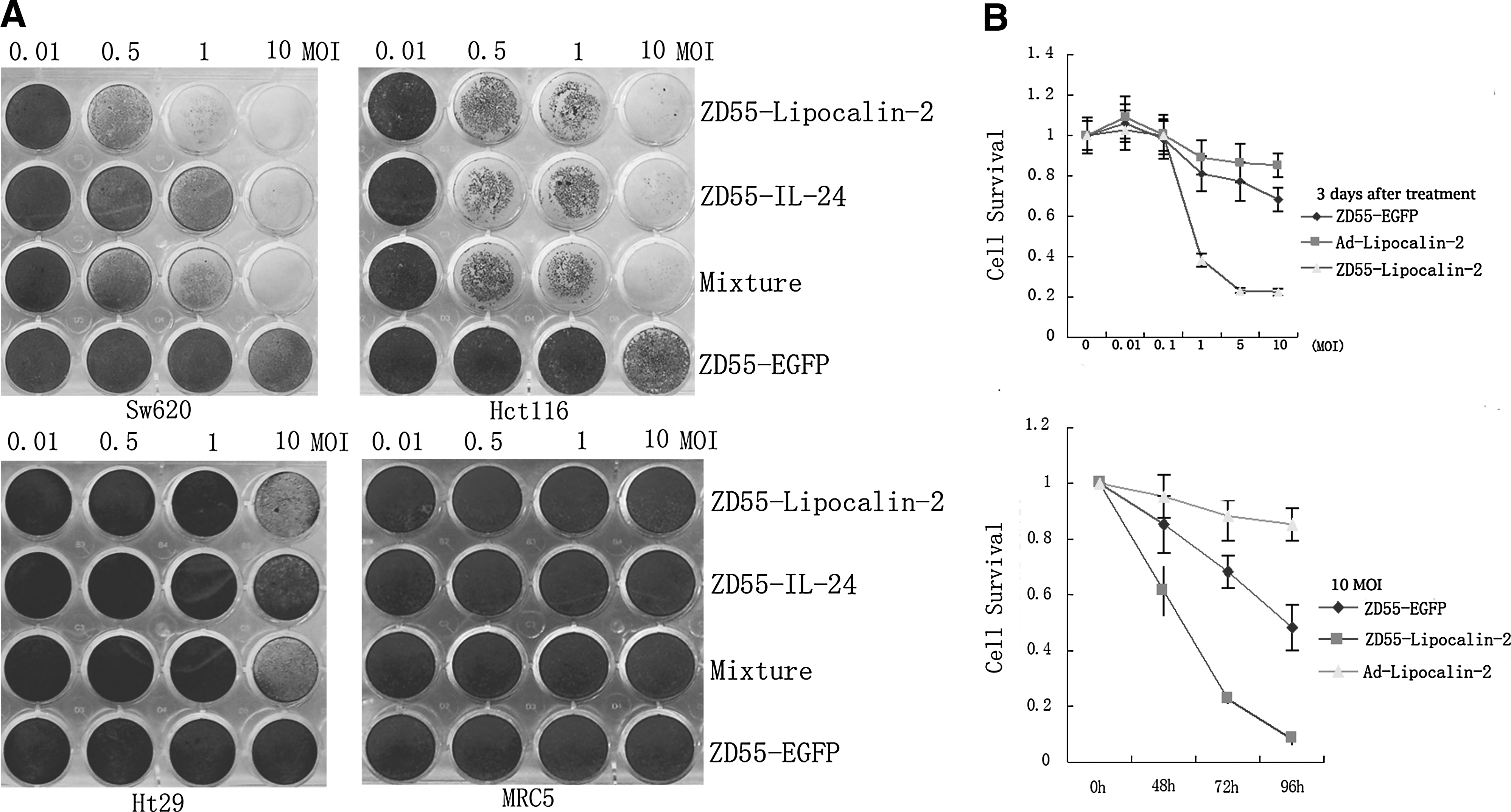
The cytopathic effects and growth inhibition of ZD55-Lipocalin-2.
Antitumor efficacy of ZD55-Lipocalin-2 in tumor xenografts
To investigate the antitumor efficacy of ZD55-Lipocalin-2 in vivo, nude mouse tumor xenografts (average tumor volume, [194.48±3.92 mm3]) were randomly divided into 4 groups. There was no difference for the baseline tumor volume between the groups by one-way ANOVA (p=0.372). Tumors treated with ZD55-Lipocalin-2 exhibited more growth inhibition compared with those treated with PBS (p<0.001), Ad-Lipocalin-2 (p<0.001), or ZD55-EGFP (p<0.001). Tumor growth inhibition in the group Ad-Lipocalin-2 was similar to that in the group ZD55-EGFP (p=0.118), but both Ad-Lipocalin-2 and ZD55-EGFP showed a significant antitumor efficacy compared with PBS (p<0.001) (Table 1 and Fig. 3A). After a 30-day post-treatment interval, all tumors were removed from the nude mice. The weight of each tumor and the average inhibition rate were evaluated (Table 2). The weight of tumors treated with ZD55-Lipocalin-2 was significantly less than that of the tumors treated with PBS (p<0.01). The tumor weight of group ZD55-EGFP or group Ad-Lipocalin-2 was also less than that of group PBS (p<0.05), but there was no significant difference between group ZD55-EGFP and group Ad-Lipocalin-2 (p=0.551). The tumor growth inhibition rate in group ZD55-Lipocalin-2 was higher than that in any of the other three groups. In the survival analysis, the failure event was defined as the tumor volume more than 600 mm3. Prognosis of group ZD55-Lipocalin-2 was the best among four groups. By pairwise comparisons for survival analysis, there was a difference between group ZD55-Lipocalin-2 and any of the other three groups; no difference was observed between group ZD55-EGFP and group Ad-lipocalin-2 (Fig. 3B).
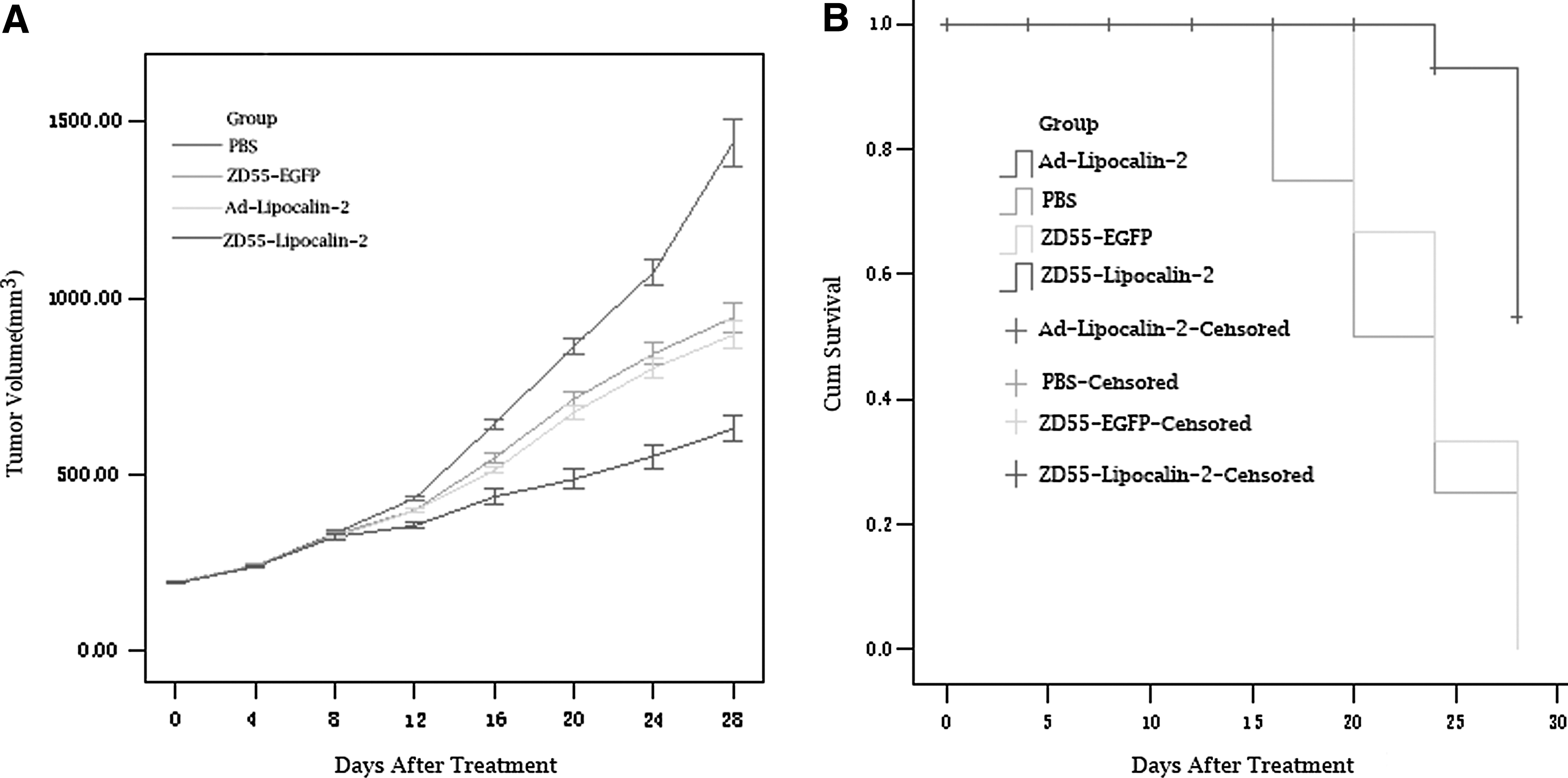
Antitumor efficacy of ZD55-Lipocalin-2 in nude mice.
P<0.05 (compared with group PBS).
P<0.01 (compared with group PBS)
RTV=post-therapeutic tumor volume/pretherapeutic tumor volume.
PBS, phosphate-buffered saline; RTV, relative tumor volume.
p<0.05 (compared with group PBS).
p<0.01 (compared with group PBS).
AIR: The average inhibition rate=[(average tumor weight of control group − average tumor weight of experimental group)/average tumor weight of control group]×100%.
Apoptosis induced by ZD55-Lipocalin-2 in vivo
Apoptosis in situ was detected by TUNEL. Positive cells were those with brown particles in the nucleus (Fig. 4). The apoptosis index (AI) was counted (AI=[number of positive cells/1000 cancer cells]×100%). The results are shown in Table 3. The AI of group ZD55-Lipocalin-2 was significantly higher than that of any of the other three groups (p<0.01). The difference of the AI between group ZD55-EGFP and group Ad-lipocalin-2 was not significant (p=0.665).
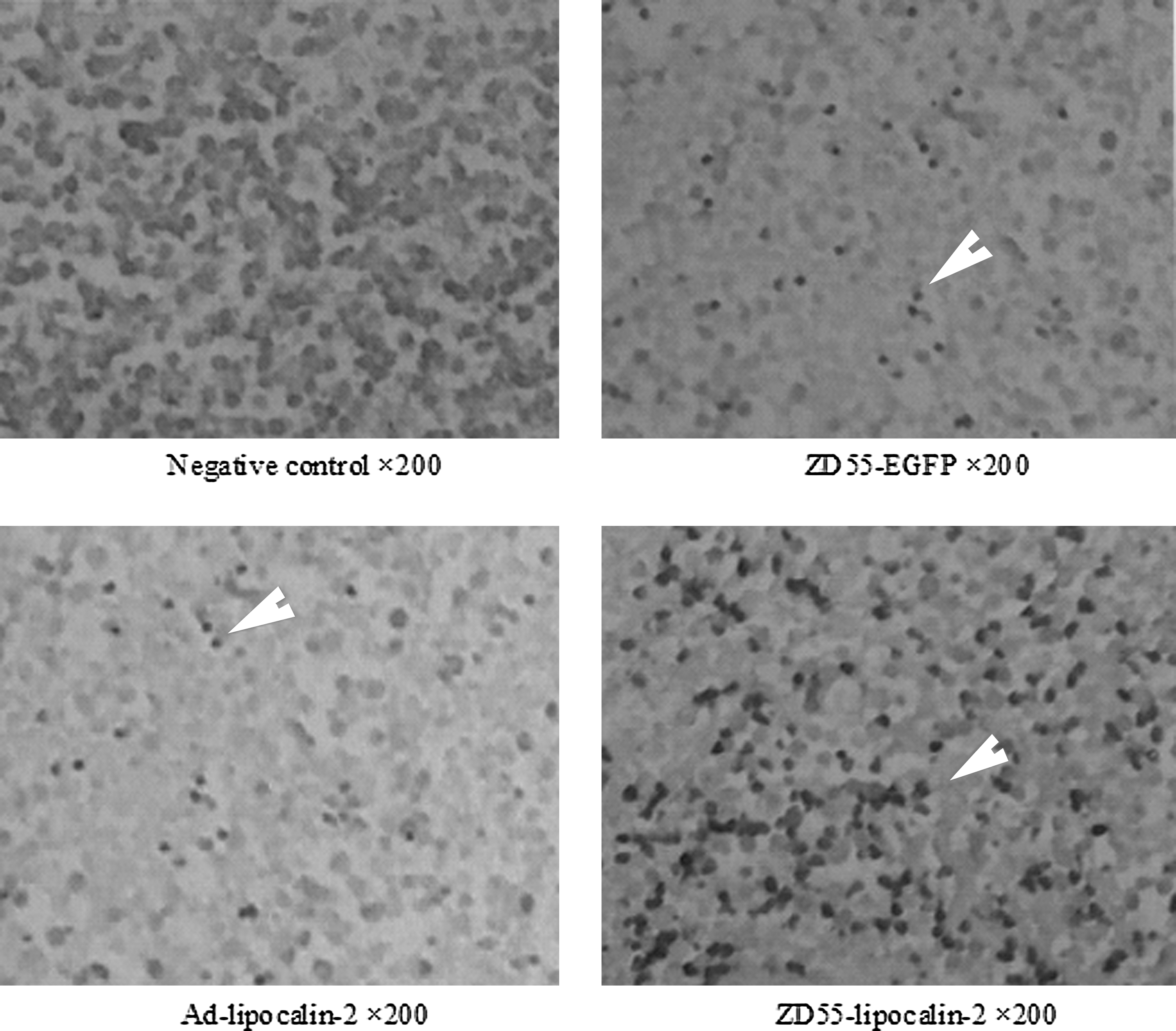
Terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) staining. Apoptotic cancer cells with brown particles (arrow indicated) in group ZD55-Lipocalin-2 were significantly more prevalent than in any of the other three groups. The details of the apoptosis index are shown in Table 3.
p>0.05 (compared with group ZD55-EGFP).
p<0.05 (compared with group ZD55-EGFP).
AI=(number of apoptotic cells/1000 cancer cells)×100%.
AI, apoptosis index.
ZD55-Lipocalin-2 suppresses angiogenesis in colorectal cancer
To confirm whether ZD55-Lipocalin-2 can inhibit tumor angiogenesis, VEGF expression and the MVD were detected by immunohistochemistry in tissue specimens. The expression of VEGF in group ZD55-Lipocalin-2 was lower than that in any of the other two groups. MVD of group ZD55-Lipocalin-2 was the lowest among the three groups (p<0.01). No significant difference was observed between group ZD55-EGFP and group Ad-Lipocalin-2 (p=0.171) (Table 4 and Fig. 5).

Microvessel density (MVD). There was more cancer cell necrosis in group ZD55-Lipocalin-2 than in group ZD55-EGFP or group Ad-lipocalin-2 in hematoxylin and eosin stain. The expression of vascular endothelial growth factor (VEGF) and the MVD in group ZD55-Lipocalin-2 were lower than that in Group Ad-lipocalin-2 or group ZD55-EGFP (arrow indicated).
p>0.05 (compared with group ZD55-EGFP).
p<0.05 (Compared with group ZD55-EGFP).
Discussion
Oncolytic viruses represent a promising platform for novel cancer therapy. The combination of oncolytic viruses with existing drugs show particular promise and have exhibited a much greater ability to kill cancer cells. 19 –22 Oncolytic viruses exert an intrinsic anticancer activity mainly from the lysis of cancer cells through its own selective replication. CTGVT integrates a single oncolytic virus with an anticancer gene to generate stronger antitumor efficacies. ZD55-Lipocalin-2 (an example of CTGVT) shows well the combined antitumor effects from ZD55 oncolytic adenoviruses and Lipocalin-2. It inhibited the tumor cell growth in vitro and in vivo.
Our results indicated that the viability of Sw620 significantly decreased after treatment with ZD55-Lipocalin-2. The growth inhibition of ZD55-Lipocalin-2 was stronger than that of ZD55-EGFP or Ad-lipocalin-2. In crystal violet staining, one positive control therapeutic agent, ZD55-IL-24, 17 was used, which had been reported to be effectively therapeutic in colorectal cancer. Tumor-specific CPEs of ZD55-Lipocalin-2 were similar to those of ZD55-IL-24 in Hct116, but were stronger in Sw620 and Ht29 cells. The antitumor efficacy of ZD55-Lipocalin-2 was greater in Sw620 or Hct116 than in Ht29. Sw620 and Hct116 tumor cells had negative expression of Lipocalin-2, whereas Ht29 cells showed high Lipocalin-2 expression, 12 so the antitumor efficacy of ZD55-Lipocalin-2 might be associated with Lipocalin-2 expression in tumor cells. Sw620 tumor xenografts were used to detect the antitumor activity of ZD55-Lipocalin-2 in vivo, and the growth curve showed that ZD55-Lipocalin-2 had the strongest ability to inhibit tumor growth among the four groups. The tumor growth inhibition rate in group ZD55-Lipocalin-2 (40.36%) was significantly higher than that in group ZD55-EGFP (16.37%) or the single-gene therapy group Ad-Lipocalin-2 (18.91%). The mean survival of group ZD55-Lipocalin-2 was longer than that of group ZD55-EGFP or group Ad-Lipocalin-2.
Why does ZD55-Lipocalin-2 has a better therapeutic effect than Ad-Lipocalin-2 or ZD55-EGFP? Because when ZD55-Lipocalin-2 infected colorectal cancer cells, it proliferated in tumor cells, lysed the tumor cells, released progeny virus, and continued to infect the adjacent tumor cells. At the same time, Lipocalin-2 was highly expressed in tumor cells accompanying its replication; thus, the antitumor effects were magnified and overlapped. It was reported that Lipocalin-2 activated the intracellular JAK/STAT pathway, inducing apoptosis, 23 but we still did not know whether it induced apoptosis in colorectal cancer cells. Thus, TUNEL was used to detect apoptosis in Sw620 tumor xenografts. We found that ZD55-Lipocalin-2 markedly induced Sw620 cell apoptosis. Moreover, it produced more necrosis in tumors detected by the hematoxylin and eosin stain. The AI in group ZD55-Lipocalin-2 was higher than that in group ZD55-EGFP (3.73-fold) or group Ad-Lipocalin-2 (4.00-fold). In addition, Tong et al. 24 found that Lipocalin-2 protein inhibited angiogenesis partly by blocking VEGF production in pancreatic cancer cells. In our study, expression of VEGF and MVD in group ZD55-Lipocalin-2 was significantly lower than that in either of the other two groups. Therefore, Lipocalin-2 also inhibited angiogenesis in colorectal cancer.
As of yet, 40% of colorectal cancers encountered in the clinic include the K-ras mutations that are resistant to chemotherapy. 2 In malignant RAS-transformed cells, Lipocalin-2 induced an epithelial phenotype, blocking their capacity for growth, invasiveness, and metastasization. 11,25 In our study, Sw620 and Hct116 cells that included the K-ras mutations were sensitive to ZD55-Lipocalin-2, especially Sw620 cells in vivo. As such, ZD55-Lipocalin-2 may be a promising agent for treating colorectal cancer that is resistant to chemotherapy.
In summary, we developed a novel antitumor agent for colorectal cancer that can induce apoptosis, lyse tumor cells, and suppress tumor angiogenesis. The synergetic anticancer potential of ZD55-Lipocalin-2 is greater than that of the gene therapy (Ad-Lipocalin-2) or oncolytic adenovirus therapy (ZD55-EGFP) alone. Therefore, ZD55-Lipocalin-2 might have a therapeutic potential for the treatment of colorectal cancer, especially for cancer cells with K-ras mutations.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (81001007 and 81272533), the Program for Young Excellent Talents in Tongji University (2008KJ060), and Youth Fund of the Shanghai 10th People's Hospital (10RQ105). Thanks to Aaron Leppin (Mayo Clinic) for language editing.
Disclosure Statement
The authors declare no potential conflict of interests.