
Abstract
The efficacy of 131I-metaiodobenzylguanidine (MIBG) therapy relies on norepinephrine transporter (NET) function. The ionic make-up of the extracellular fluid critically controls neuronal cell activity and can also affect substrate transport. In this study, we explored the effect of treatment with elevated KCl concentration on MIBG uptake in SK-N-SH neuroblastoma cells. KCl stimulation caused a rapid increase of 131I-MIBG uptake in a manner that was calcium-dependent and accompanied by activation of calcium/calmodulin-dependent protein kinase (CaMK)II. The effect was completely abolished by KN93, an inhibitor of CaMKI, II, and IV. STO609, a selective inhibitor of CaMK kinase required for activation of CaMKI and IV, but not CaMKII, only modestly attenuated the response. The KCl effect was also completely abrogated by ML7, a selective inhibitor of myosin light chain kinase (MLCK). This restricted form of CaMK activates myosin, which is required for vesicle trafficking. Saturation kinetic analysis revealed KCl stimulation to increase maximal transport velocity without affecting substrate affinity. In conclusion, KCl stimulation rapidly upregulates NET function through the CaMK pathway via activation of CaMKII and MLCK. These findings allow a better understanding of how NET function is acutely modulated by the ionic environment, which in turn may ultimately help improve the efficacy of 131I-MIBG therapy.
Introduction
Neuroblastoma is the most common extracranial solid tumor in children, and it often presents as a progressive disease with poor outcome despite aggressive multimodal therapy. 1,2 For patients with inoperable or disseminated neuroblastoma, one of the most actively employed treatment approaches is targeted radiotherapy using 131I-labeled metaiodobenzylguanidine (MIBG). 1 –5 MIBG is a guanethidine-derived analogue of norepinephrine (NE) that accumulates in cells through the NE transporter (NET). As neuroblastomas derive from primordial neural crest cells, over 90% of the tumors express NET. 1,2 Therefore, systemically administered 131I-MIBG can be specifically taken up by tumor cells to cause β-ray irradiation-induced cell death. Despite the attractiveness of this strategy, however, only about 30% of refractory or relapsed neuroblastoma patients respond to 131I-MIBG therapy under current treatment protocols. 2,3
The efficacy of 131I-MIBG therapy could be improved by augmenting the magnitude of radiotracer uptake, which is solely determined by the activity of membrane-localized NET. 6,7 As such, a better understanding of physiologic conditions that influence the expression and/or functional state of this transporter could help devise methods to improve 131I-MIBG therapy. 8,9 A characteristic crucial to the function of neuron cells is the ionic make-up within the cells and in the surrounding extracellular fluid. In NET-expressing cells, the ionic environment can also influence NE transport capacity. 10 –12 Among extracellular fluid ions, the potassium ion has particularly great bearing in neuronal function. As such, fluctuations in extracellular potassium concentration can accelerate neuron activity and lead to their depolarization. 13,14 In a previous study on a pheochromocytoma cell line, brief exposure to elevated KCl concentration was shown to increase cellular NE uptake. 15 A better understanding of how KCl concentration modulates NET function and recognition of the molecular mechanism responsible for this response could provide insight useful for developing strategies to enhance 131I-MIBG therapy for neuroblastoma. NET activity is largely modulated through specific intracellular signaling cascades, and candidate pathways for its acute regulation include protein kinase C (PKC), 16 mitogen activated protein kinase (MAPK), 17 phosphatidyl inositol-3 kinase (PI3K), and Ca2+/calmodulin-dependent protein kinase (CaMK). 18
In this study, we thus investigated the acute effects of KCl stimulation on 131I-MIBG uptake in a human neuroblastoma cell model and further explored the intracellular signaling mechanisms that mediate this response.
Materials and Methods
Cells and reagents
Human neuroblastoma SK-N-SH cells (American Type Culture Collection) were maintained at 37°C and 5% CO2 in minimum essential media supplemented with 10% fetal bovine serum, 2 mM glutamine, 100 mU/L penicillin, and 100 mg/L streptomycin. Experiments were performed when cell confluence reached 80% at 2 to 3 days after seeding.
131I-MIBG with a specific activity of 1178 Ci/mmol was obtained from the Korea Atomic Energy Research Institute (Korea). All other reagents including desipramine hydrochloride and unlabeled MIBG were from Sigma.
131I-MIBG uptake measurement
Uptake experiments were performed on 12-well or 24-well plates containing 0.4×106 cells in 1 mL or 0.2×106 cells in 0.5 mL of buffer, respectively. Cells washed twice with 1 mL Ca2+-free Krebs-Ringer (KR) buffer were preincubated for 5 minutes in KR buffer containing 130 mM NaCl, 0.25 mM MgCl, 0.7 mM Na2HPO4, 1.5 mM NaH2PO4, 10 mM D-glucose, and 2.2 mM CaCl2. KCl stimulation was performed by exposure to KR buffer containing 5 to 40 mM of KCl with equimolar amounts of NaCl removed to maintain osmolarity. Control cells were treated with normal KR buffer. A separate group of cells were tested for KCl effect when calcium was chelated by 1 mM ethyleneglycol tetraacetic acid (EGTA).
Cells were incubated at 37°C in 5% CO2 for 30 minutes with 37 kBq of 131I-MIBG added to each well. Cells were then rapidly washed twice with ice-cold phosphate buffered saline (PBS) and measured for cell associated radioactivity on a γ-counter (Wallace). Nonspecific uptake was measured in the presence of 50 μM of the selective NET inhibitor desipramine, and specific uptake was calculated by subtracting nonspecific radioactivity from total counts.
Kinase inhibition experiments
Involvement of candidate intracellular signaling pathways was assessed by addition of specific inhibitors or activators to the cells 30 minutes prior to treatment with KCl. This included inhibitors of the PKC pathway (staurosporine, 1 μM), PI3K pathway (LY294002, 30 μM), and MAP kinase pathway (PD98059, 25 μM). The PKC activator phorbol 12-myristate 13-acetate (PMA, 1 μM) was also evaluated. The role played by the CAMK pathway was investigated with the CAMK II inhibitor KN93 (10 μM) and the CAMK kinase inhibitor STO609 (5 μM). The potential role of myosin light chain kinase (MLCK) was evaluated by the specific inhibitor ML-7 (5-iodonaphthalene-1-sulfonyl homopiperazine, 10 μM). Any effect of the treatments on cell viability was visually assessed under a light microscopy.
Saturation kinetic analysis of MIBG transport
For saturation binding assays, 0.2×106 cells in 0.5 mL KR buffer were incubated for 30 minutes with graded concentrations of 131I-MIBG ranging from 0 to 2 μM following 5 minutes treatment with 40 mM KCl or vehicle. 131I-MIBG uptake was measured as described above, and nonspecific uptake was determined with excess desipramine for each 131I-MIBG concentration. Saturation transport curves were drawn by nonlinear fitting and the maximal transport velocity (Vmax) and half-maximal transport (Km) was calculated by GraphPad Prism® V. 3.02 software (GraphPad Software, Inc.).
Western blotting of phosphorylated CaMKII
Western blotting was performed using 100 mm plates containing 7.9×106 cells in 10 mL media. Cells were washed with cold PBS and solubilized for 20 minutes at −20°C with 50 μL of protein extraction solution (Pro-prep™) containing 0.1 μM aprotinin, 1 μM leupeptin, 1 μM pepstatin A, 1 mM phenylmethylsulfonyl fluoride, and 1 mM ethylendiamine tetraacetic acid. Supernatant obtained by 15 minutes centrifugation at 15,000 g at 4°C, was measured for protein concentration, and protein was separated by sodium dodecyl sulfate polyacrylamide gel (12%) electrophoresis. The protein was transferred to a polyvinylidene fluoride membrane (Bio-Rad), which was then incubated with a primary phospho-CaMKII (Thr286) antibody (1:1000 dilution), followed by an antirabbit HRP-conjugated secondary antibody (Santa Cruz Biotechnology; 1:1000 dilution). Immunoreactive protein was detected by an enhanced chemiluminescence detection system, and protein band intensities were measured using a GS-800™ calibrated densitometer and Quantity One® software (Bio-Rad).
Statistical analysis
All cell uptake experiments were performed in triplicates and repeated at least twice. Results were expressed as mean±SD. Statistical comparison of uptake level between groups was performed by Student t-tests. p-values of less than 0.05 were considered statistically significant.
Results
KCl enhances SK-N-SH cell MIBG uptake in dose- and time-dependent manners
Our experimental conditions were such that baseline SK-N-SH cells took up 2 to 3% of the added 131I-MIBG (specific activity of 1178 Ci/mmol) in 30 minutes. The level of nonspecific uptake as determined by 100 μM desipramine was ∼17%.
When SK-N-SH cells were stimulated with 40 mM KCl for 5 minutes, 131I-MIBG uptake was significantly elevated to 140.0%±4.2% of untreated cells. When 5 minutes KCl stimulation was repeated twice, uptake further increased to 177.8%±7.2% of controls (Fig. 1A). The response was rapid with uptake plateauing around 179% of controls by 10 minutes treatment with 40 mM of KCl (Fig. 1B). Augmented 131I-MIBG uptake by KCl occurred in a concentration-dependent manner with uptake increasing to 137.1%±12.9%, 145.2%±5.7%, 159.7%±6.3%, and 169.2%±4.5% of controls by 5, 10, 20, and 40 mM, respectively (Fig. 1C). Based on these results, 5 minutes treatment with 40 mM KCl was used for further stimulation experiments.
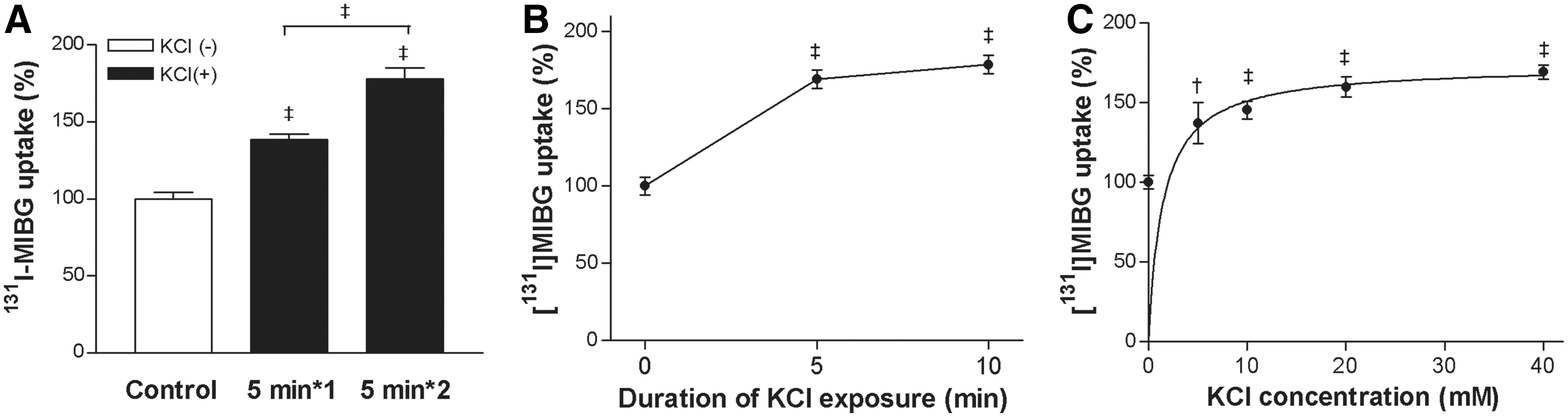
Effect of KCl on SK-N-SH cell MIBG uptake.
Central role of the calmodulin signaling on KCl-stimulated MIBG uptake
Chelation of calcium with EGTA caused a 27.4%±8.2% lowering of baseline 131I-MIBG uptake (Fig. 2A). EGTA also appeared to attenuate KCl-stimulated 131I-MIBG uptake when compared with controls without EGTA, although the difference was not significant when each group was compared to their respective controls (Fig. 2A). Treatment with the CaMK inhibitor KN93 (10 μM) completely abolished the KCl effect, reducing MIBG uptake to levels even lower than controls (54.3%±4.4%). Baseline MIBG uptake level was also reduced by KN93 (53.1%±3.0% of controls; Fig. 2B). The selective CaMK kinase inhibitor STO609 (5 μM) partially blocked KCl-stimulated 131I-MIBG uptake without influencing baseline uptake level (Fig. 2C).

Central role of calcium and the CaM pathway on the KCl effect.
Western blots displayed increased phosphorylated CaMKII (pCaMKII) level in cells stimulated with KCl, but this was blocked by KN93 (Fig. 2D).
PKC, PI3K, and MAPK signaling are only partially involved in the KCl effect
Stimulation of PKC activity with 1 μM PMA caused only a small decline of baseline and KCl-induced 131I-MIBG uptake (Fig. 3A). Blocking of the PKC pathway with 1 μM staurosporine also had little or no effect on baseline or KCl-stimulated 131I-MIBG uptake (Fig. 3B).

Partial Roles of PKC, PI3K, and MAPK on the KCl Effect.
Treatment of cells with the specific PI3K inhibitor LY294002 (30 μM) partly suppressed the ability of KCl to stimulate 131I-MIBG uptake from 171.0%±6.1% to 127.7%±5.1% of controls (Fig. 3C). The MAPK inhibitor PD98059 (25 μM) also resulted in lower levels of KCl-stimulated (124.6%±11.7% of controls) and baseline 131I-MIBG uptake (Fig. 3D).
Effect of MLCK on KCl-stimulated MIBG uptake
The potential role of MLCK, a restricted form of CaMK, on KCl-stimulated 131I-MIBG uptake was examined with a selective inhibitor. As a result, 10 μM ML7 completely abrogated the ability of KCl to enhance 131I-MIBG uptake (14.8%±0.8% of controls), and also dramatically reduced baseline uptake (11.9%±2.4% of controls; (Fig. 4A).
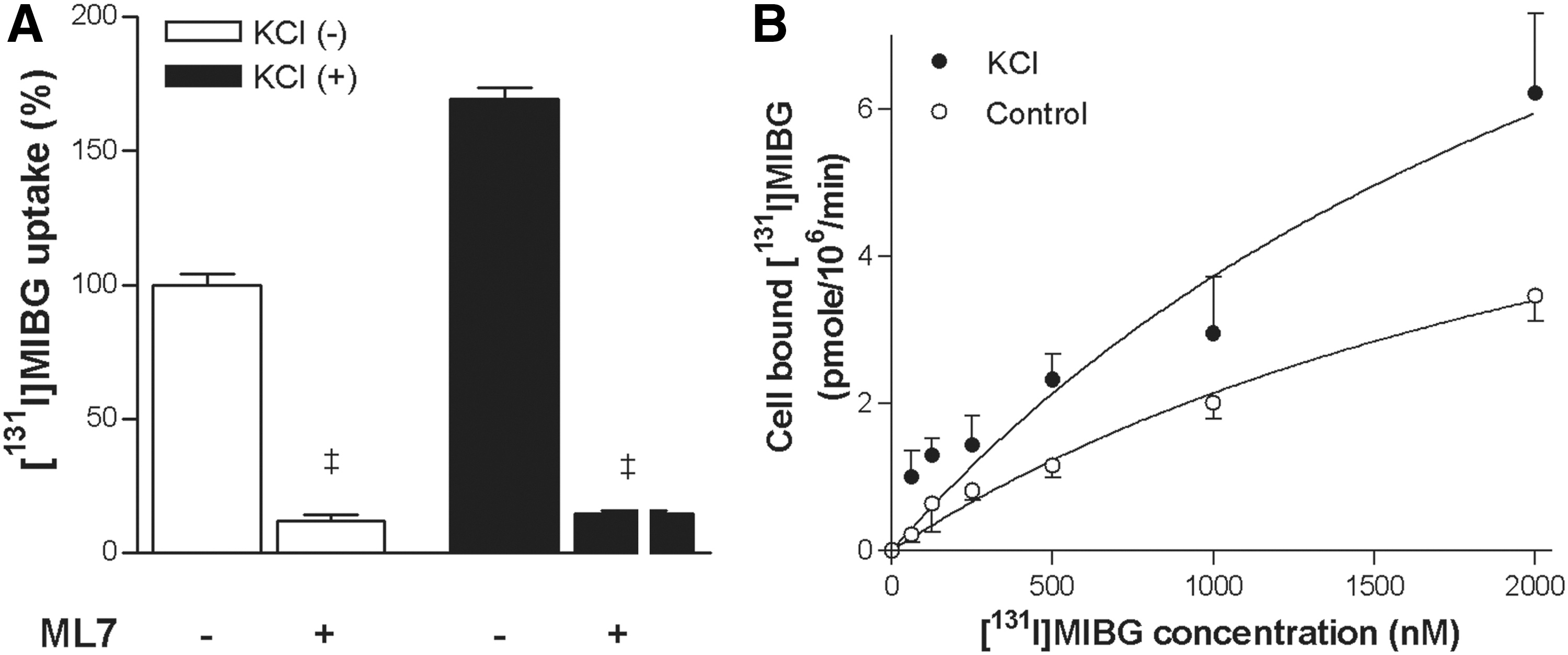
Role of MLCK and saturation transport kinetics.
Effect of KCl stimulation on MIBG transport kinetics
Saturation kinetic analysis by incubation with increasing concentrations of 131I-MIBG yielded divergent saturation binding curves for control and KCl-stimulated cells. In the basal state, the Vmax was 8.2±1.8 pmol/106-cells/min and the Km value was 2.8±0.9 μM. When cells were stimulated with KCl, there was a substantial increase in Vmax to 14.7±5.4 pmol/106-cells/min, whereas the Km value was not significantly affected (2.9±1.6 μM; Fig. 4B). These results suggest that KCl stimulation increases SN-N-SH cell 131I-MIBG uptake by augmenting maximum NE transport capacity.
Discussion
NE uptake by its cognate transporter is substantially influenced by the ionic environment, as exemplified by the absolute requirement of Na+ and Cl− for its substrate binding. 10,11 Also, an appropriate balance of K+ ions across the cell membrane is essential for proper NET function. It is suggested, for instance, that a negative membrane potential created by high extracellular K+ concentration can contribute to the driving force for NE transport. 14
This study demonstrates that brief stimulation of SK-N-SH neuroblastoma cells with KCl dose-dependently upregulates 131I-MIBG uptake in an acute fashion that occurs within a matter of minutes. The function of monoamine transporters including NET are finely tuned by intracellular signaling mechanisms, most commonly through phosphorylative activation by protein kinases. 6,7 We therefore tested the involvement of candidate intracellular signaling pathways to further dissect the mechanism for the KCl effect. The most thoroughly investigated signaling cascade related to monoamine transporter function is the PKC pathway. Studies with various cell types have shown PMA-induced PKC activation to suppress the reuptake of neurotransmitter monoamines including NE, presumably through sequestration of transporters away from the plasma membrane. 16 Other major intracellular signaling systems implicated in the regulation of monoamine transport capacity include PI3K and MAPK pathways. 17 In our results, neither activation nor inhibition of PKC activity had any noteworthy influence on KCl-stimulated 131I-MIBG uptake. Further, selective inhibitors of PI3K and MAPK pathways caused only modest blocking of the KCl effect.
Another important intracellular signaling system implicated in monoamine transporter function is the calcium-dependent calmodulin pathway. 18 –21 As calcium is maintained at extremely low concentrations in resting cells, it can function as a pervasive second messenger when mobilized via specific stimuli. KCl has been proposed as a calcium-sensitizing stimulus that can induce membrane depolarization via calcium entry through voltage-operated channels. 19 Several lines of evidence indicate that calcium signaling can alter the transport kinetics of monoamine neurotransmitters such as dopamine and serotonin. 20,21 In our experiments, the ability of KCl to enhance 131I-MIBG uptake was efficiently suppressed by the chelator EGTA, pointing to calcium dependence of the response. Depolarization of neuroblastoma cells by high K+ also stimulates MIBG release. Although this was suggested to be mediated by the NET working in reverse mode, it was observed to mainly occur in a calcium-independent fashion. 22
Calcium signaling in cells is predominantly mediated through calmodulin, a multifunctional calcium-binding messenger protein that transduces various biological processes by target protein interaction. 23,24 A key target for the calcium/calmodulin complex is a family of structurally related serine/threonine-specific protein kinases called CaMKs. Among these, CaMKII is present in high concentrations in neuronal synapses and plays pivotal roles in transcription factor regulation and neurotransmitter secretion. In our study, SK-N-SH cells stimulated with KCl displayed an increase in phosphorylated CaMKII level, and KN93 completely blocked the ability of KCl to activate CaMKII and enhance 131I-MIBG uptake. A previous study by Mandela and Ordway in pheochromocytoma PC12 cells observed a similar finding of increased NE uptake by KCl stimulation that was inhibited by KN93. 15 Although KN93 is a well-known CaMKII inhibitor, this small molecule is now recognized to also suppress CaMKI and CaMKIV activity. Therefore, the possibility that the KCl effect is mediated by these kinases cannot be excluded by use of KN93 alone. A major difference in these kinases is that interaction with calcium/CaM is sufficient for CaMKII activation, whereas CaMKI and CaMKIV activation also requires CaMK kinase-mediated phosphorylation. We therefore dissected this issue by blocking CaMK kinase activity with the selective inhibitor STO609. 23 As a result, CaMK kinase inhibition only modestly attenuated the KCl effect, indicating that CaMKII, rather than CaMKI or CaMKIV, is the main contributor.
In addition to the multifunctional forms, MLCK is a restricted type of CaMK that mediates cellular processes linked to the biological functions of its sole substrate, the regulatory light chain of myosin. 25 This motor protein is known to mediate actin-based rapid transport of intracellular organelles, 26 including glucose transporter containing vesicles. 27,28 Further, inhibition of MLCK has been shown to block synaptic vesicle pool mobilization in response to calcium signaling. 29 In our study, KCl-stimulated 131I-MIBG uptake was completely abrogated by the selective inhibitor ML7, demonstrating dependence on MLCK activity. Our kinetic analysis further showed KCl to significantly increase maximum transport velocity without affecting substrate affinity of NETs. This finding is consistent with a previous study where Ca2+-stimulated NE uptake was accompanied by elevations in Vmax and increased desipramine binding. 17 Taken together, our findings suggest that KCl stimulation may act through MLCK to increase the surface density of functional NET.
In conclusion, brief stimulation with KCl rapidly enhances 131I-MIBG uptake of SK-H-SH neuroblastoma cells. This response is mediated by the Ca2+/calmodulin pathway, through the combined activity of CaMKII and MLCK, which act to upregulate the functional capacity of surface NET. These findings may allow a better understanding of how NET function is acutely modulated by the ionic environment, and may ultimately help improve the efficacy of 131I-MIBG therapy.
Footnotes
Acknowledgments
This work was supported by Samsung grant (#C-A9-225-3) and Konkuk University.
Disclosure Statement
No competing financial interests exist.