
Abstract
Purpose:
Lung cancer is a leading cause of cancer death worldwide. Efficacy of radiation therapy on lung cancer is hindered by many factors. Among these, both cancer stem-like side population (SP) and main population (MP) cells may contribute to tumorigenesis and resistance to radiation therapy. However, the detailed mechanism responsible for this effect remains unknown.
Materials and Methods:
The SP and MP cells were obtained from lewis lung carcinoma cells and analyzed the DNA dye (Hoechst 33342) method and flow cytometry. The levels of ABCG2 and CD133 markers were examined by reverse transcription polymerase chain reaction, Western blot, and immunofluorescence. The effects of ionizing radiation (IR) on the growth and apoptosis of SP and MP cells were determined by 3-(4, 5-dimethylthiazol-2-y)-2, 5-diphenylterazolium bromide (MTT), colony formation, and apoptosis assays. Mitochondrial membrane potential and intracellular reactive oxygen species production were measured by flow cytometry. Finally, the expression of Bax, Bcl-xL, Bcl-2, activated caspase-3 and caspase-9 proteins were examined by Western Blot.
Results:
IR decreased proliferation, increased apoptosis and mitochondria damage in MP, but not in SP cells. Protein levels of Bcl-2 and Bcl-xl were decreased, while Bax expression was increased in MP cells following IR exposure. In addition, increased activation of caspase-3 and caspase-9 were detected after IR exposure in MP cells, but not in SP cells.
Conclusions:
Our results show that IR decreases proliferation, increases apoptosis, and induces mitochondria damage in MP cells, but not in SP cells, through increased Bax and decreased Bcl-2 and Bcl-xl protein expression. This protein expression pattern induces activation of caspase-3 and caspase-9. This study suggests that IR exposure targets MP cells through a mitochondrial apoptosis pathway. However, more work is required to further confirm these results using in vivo xenograft models. More importantly, further studies are warranted to elucidate the radioresistant mechanisms of SP cells.
Introduction
Lung cancer continues to be the most commonly diagnosed cancer, as well as the leading cause of cancer death in the world. Lung cancer accounts for 13% (1.6 million) of the total cancer deaths in the year 2010. 1 In the last decade, radiochemotherapy, such as ionizing radiation (IR), has been successfully applied in clinical arena for the treatment of lung cancer. Irradiation induces death of tumor cells by causing DNA damage. 2,3 However, the efficacy of radiation therapy is hindered by many factors in lung cancer cells, such as the targeting of normal cells, activation of cell survival pathways, inhibition of tumor suppressor genes, and blockade of DNA repair signaling pathways. 4,5
Recent studies suggested that cancer stem cells (CSCs) may be responsible for tumorigenesis and contribute to the resistance of cancer cells to therapeutic interventions. 6,7 CSCs, a unique subpopulation in cancer cells, are regarded as possessing the ability to initiate tumor growth and sustain self-renewal. Dick and Bonnet. 8 first demonstrated the existence of CSCs in human acute myeloid leukemia more than a decade ago. Since then, numerous studies have provided evidences for the existence of CSCs in solid tumors, such as brain, 9 breast, 10 and prostate tumors. 11 The most valuable technique to isolate and identify CSCs is dye exclusion. Stem cells have an ability to exclude fluorescent DNA-binding dye (Hoechst 33342). These cells are resistant to Hoechst 33342 staining due to the ABCG2 (BCRP1) transporter. 12 Other researchers have also designated a small subset of cells as “side population (SP)” cells, which are a minor population of cells that has been identified in a variety of cancers and have many CSC-like properties. These properties include self-renewal potential, tumor-forming capacity, and a chemoresistant phenotype. 13 –15 Interestingly, CD133, a surface marker, has been shown to be expressed in SP cells isolated from many cancers, including lung, 15 hepatocellular, 16 colorectal, 17 prostate, 18 and pancreatic adenocarcinomas. 19
The release of mitochondria cytochrome c has been regarded as an initiating event in the apoptotic process, and can trigger activation of caspase-3. 20 –22 A series of studies have identified reactive oxygen species (ROS)-induced apoptosis in various cancer cells, including hepatoma cells and HeLa cells. 23,24 Further, Yang et al. 25 found ROS could affect the mitochondrial membrane potential (MMP) in human melanoma cells. The mitochondrial cell death pathway is regulated by the ratio of pro- to antiapoptotic proteins, including members of the Bcl-2 family. Among these family members, Bax plays a key role in the loss of mitochondrial transmembrane potential. 26
To our knowledge, there are no studies on the effects of IR on lewis lung carcinoma (LLC) SP cells and main population (MP) cells. This in vitro study is focused on the induction of apoptosis by IR in LLC cells, and explores the associated molecular signaling pathways. We isolated SP and MP cells from LLC cells to identify their radiosensitivity. We observed that IR induced apoptosis in MP cells, which was accompanied by a loss of MMP. Our findings have revealed differential sensitivity of lung carcinoma stem-like SP and MP cells to IR. This finding may be useful in improving the clinical effectiveness of radiotherapy for the treatment of malignant tumors.
Materials and Methods
Cell culture and SP cell analysis
LLC cells were obtained from American Type Culture Collection (Bethesda, MD) and grown in RPMI-1640 medium (Hyclone, Logan, UT) supplemented with 10% fetal bovine serum and antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin). Cells were maintained in a humidified cell incubator with 5% CO2 at 37°C. LLC cells were suspended at 1×106 cells/mL and then incubated at 37°C for 60 minutes with 5 μg/mL Hoechst 33342 (Sigma Chemicals, St Louis, MO). The control cells were cultured in the presence of 500 μM verapamil (Sigma). After incubation, 1 μg/mL propidium iodide (PI) was added to identify dead cells. Analysis and sorting of the SP cells was performed using a FACS Vantage SE cytometer (Becton Dickinson, San Jose, CA). Hoechst 33342 was excited using a UV laser at 350 nm and fluorescence emission was measured at both 402–446 nm for Hoechst blue and 640 nm for Hoechst red.
Reverse transcription polymerase chain reaction
Total RNA was isolated from MP and SP cells using an RNeasy Mini Kit (Biomed, Beijing, China). cDNA was reverse transcribed with 1 μg of total RNA using a TaKaRa Reverse Transcription Kit (TaKaRa Dalian, Dalian, China) and was amplified using the following primers. CD133 primers were 5′- ACCGACTGAGACCCAACATC -3′ (sense) and 5′- GGTGCTGTTCATGTTCTCCA -3′ (antisense). ABCG2 primers were 5′- AGCTGCAAGGAAAGATCCAA -3′ (sense) and 5′- TCCAGACACACCACGGATAA -3′ (antisense). GAPDH primers were 5′- AGAAGGCTGGGGCTCATTTG-3′ (sense) and 5′- AGGGGCCATCCACAGTCTTC -3′ (antisense) and used as an internal control. The polymerase chain reaction (PCR) products were electrophoresed on a 1.5% agarose gel, and visualized by ethidium bromide staining under a UV imaging system (UVP, LLC, Upland, CA).
Radiation treatment
The LLC, MP, and SP cells were irradiated with a series of radiation doses (2, 4, 6, 8, 10 or 12 Gy) for 30 minutes. The RS 2000 Biological irradiator (Rad Source Technologies, Boca Raton, FL) X-ray unit was operated at 150 kv/50 mA, and dose rate of 2 Gy/min was used for radiation exposure.
Cell growth inhibition assays
Cell sensitivity to radiation was determined by the 3-(4, 5-dimethylthiazol-2-y)-2, 5-diphenylterazolium bromide (MTT) method. The cells described above were seeded in quadruplex in 96-well dishes at a density of 1500 cells/well. After 24 hours, the cells were treated with 0.5 mg/mL MTT for 4 hours and lysed with dimethyl sulfoxide. Absorbance rates were measured at 550–560 nm using a microplate reader (Bio-Rad, Hercules, CA). The surviving fraction and the IC50 value were determined. We used the IC50 dose to treat MP and SP cells in the following experiments.
Colony formation assay
For the colony formation assay, cells were seeded at 200 cells/well in 12-well tissue culture plates. The plates were incubated for 3 weeks in a humidified incubator at 37°C. Three weeks after seeding, colonies were stained with 0.05% crystal violet containing 50% methanol, and counted. The colonies were counted in four to five random fields for each of the duplicate samples using a microscope at 100× magnification.
Cell apoptosis assay
Apoptosis was determined using an apoptosis detection kit (KeyGEN, Nanjing, China). Briefly, cells were collected, washed twice in ice-cold PBS, and then resuspended in binding buffer at a density of 1×106 cells/mL. The treated cells were incubated with fluorescein-labeled Annexin V and PI for 20 minutes. Following the labeling reaction, the expression of Annexin at the cell surface was analyzed by a FACS Calibur machine (Model FACSC 420, Baltimore, MD). Data were analyzed using Cell Quest software from Becton Dickinson.
Determination of MMP
MMP was analyzed using the fluorescent dye 5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolycarbocyanine iodide (JC-1) following the manufacturer's protocol (KeyGEN). Briefly, cells were plated in six-well culture plate. After treatment with IR for 24 hours, cells were washed twice with PBS, harvested and incubated with 20 nM JC-1 for 30 minutes in the dark. MMP was then analyzed using the FACS Calibur machine, as described above.
Analysis of intracellular ROS production
Cells were dispensed in 10 cm culture dishes at a density of 1×106 cells/well. After 24 hours incubation, the cells were treated with IR under the conditions described above. The cells were then incubated with 2′,7′-dichlorodihydrofluorescein diacetate H(2) (DCF-DA, 10 μM) at 37°C for 15 minutes to assess ROS-mediated oxidation of DCF-DA to the fluorescent compound DCF. Next, cells were collected, and the pellets were suspended in 1 mL PBS. Signals were detected with a PNA content (FL1 fluorescence) signal detector (525 nm) using the FACS Calibur machine. 27
Immunofluorescence
Cells were fixed with 4% paraformaldehyde for 15–20 minutes, followed by washing twice in PBS at room temperature for 5 minutes, and permeabilized in PBS containing 2% Triton X-100 for 30 minutes. Nonspecific binding sites were blocked with 3% BSA in PBS for 1 hour. The primary monoclonal antibodies, diluted in 3% BSA/PBS, were applied overnight at 4°C. The cells were washed twice with PBS, and then exposed to the secondary antibody diluted at 1:100 in 3% BSA/PBS for 1 hour. In all cases, cells were counterstained with PI to evaluate nuclear condensation. For every coverslip, the cells were observed and photographed in 5 random fields using an Olympus CX71 fluorescence microscope (Olympus, Tokyo, Japan).
Western blot analysis
Cells were harvested, washed twice with PBS, lysed on ice for 30 minutes in 100 μL lysis buffer [120 mM NaCl, 40 mM Tris (pH=8.0), 0.1% NP 40], and then centrifuged at 13,000 g for 15 minutes. The supernatants were collected from the lysates and the protein concentration was determined. Aliquots of the lysates (90 μg of protein) were boiled for 5 minutes and electrophoresed using a 10% sodium dodecysulfate-polyacrylamide gel. The blots in the gels were transferred onto nitrocellulose membranes (Bio-Rad), which were then incubated with primary antibodies. Anti-Bax (sc-7480, 1:500), anti-Bcl-xL (sc-8392, 1:500), anti-Bcl-2 (sc-783, 1:500), anti-phospho-Bcl-2 (Ser 87) (sc-16323, 1:500), anti-caspase-3 (sc-136219, 1:500), and anti-β-actin (sc-103656, 1:1000) antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Anti-ABCG2 (4477, 1:200), anticaspase-9 (9504, 1:200), and anti-Bad (9292, 1:500) antibodies were purchased from Cell Signaling Technology (Beverly, MA). Anti-CD133 (MAB4310, 1:200) was purchased from Millipore Corporation (Billerica, MA). The nitrocellulose membranes were further incubated with secondary immunoglobulin-G-horseradish peroxidase conjugates. Immunostaining was detected using an enhanced chemiluminescence system (Amersham Biosciences; Westborough, MA).
Statistical analysis
Values are presented as mean and standard deviation (SD) for these experiments (mean±SD). Statistical significance was calculated with a Student's t-test. p-values less than 0.05 were considered significant. Each result in this work is representative of at least three separate experiments.
Results
SP fraction in LLC
LLC cells were labeled with Hoechst 33342 and analyzed by flow cytometry. We found that the SP cell fraction comprised 0.37% of the total cells, and that this population disappeared following treatment with the selective ABCG2 transporter inhibitor, verapamil (Fig. 1A). In addition, we showed that CSC-specific markers, ABCG2 and CD133 mRNA were significantly increased in the SP cells compared with that in MP cells (p<0.05, Fig. 1B). The expression of ABCG2 and CD133 proteins in SP cells were significantly higher than that in the MP cells (p<0.05, Fig. 1C). Immunofluorescence results demonstrated that ABCG2 and CD133 were localized in the membrane of the SP cells (Fig. 1D).
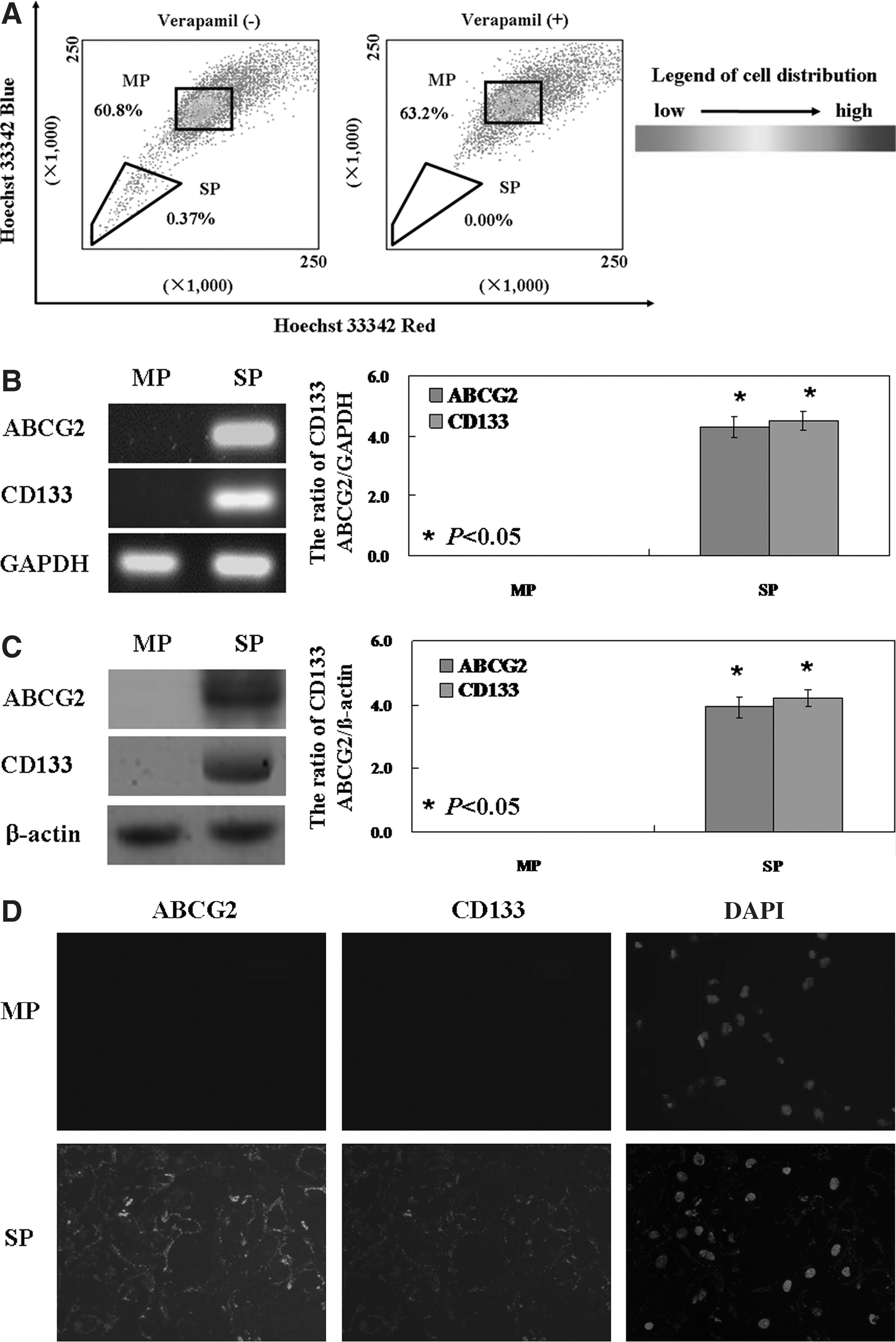
Identification of side population (SP) and main population (MP) cells in lewis lung carcinoma (LLC) cell lines. LLC (1×106 cells/mL) were cultured at 37°C for 60 minutes with 5 μg/mL Hoechst 33342 (Sigma Chemicals, St Louis, MO). The control cells were cultured in the presence of 500 μM verapamil (Sigma). After incubation, 1 μg/mL propidium iodide (PI) was added. Analysis and sorting were performed on FACS Vantage SE (
IR exposure inhibits proliferation and induces apoptosis in MP, but not in SP cells
As shown in Figure 2A, IR induced a reduction of cell viability in a dose-dependent manner in the total LLC cell population and in MP cells, as determined by the MTT assay. The IC50 value for LLC cells was 8.16 Gy. Of note, the MP and SP cells showed a significant difference in the sensitivity to IR. The SP cells showed greater IR-resistance than the MP cells, exhibiting a cell survival rate of 99.1%±0.4% following 8 Gy radiation, compared with 57.3%±0.5% survival of MP cells treated with the same radiation dose (p<0.05, Fig. 2A). Results of the colony formation assay also found that the proliferative ratio of SP cells with IR treatment is significantly higher compared with MP cells (p<0.05, Fig. 2B). In addition to the inhibitory effect on proliferation, IR also induced apoptosis in the MP cells. Annexin V/PI analysis showed that apoptotic ratio of MP cells exposed to 8 Gy radiation was 22.9%±0.43%, whereas no change was observed in the SP cells at the same dose exposure (p<0.05, Fig. 2C).
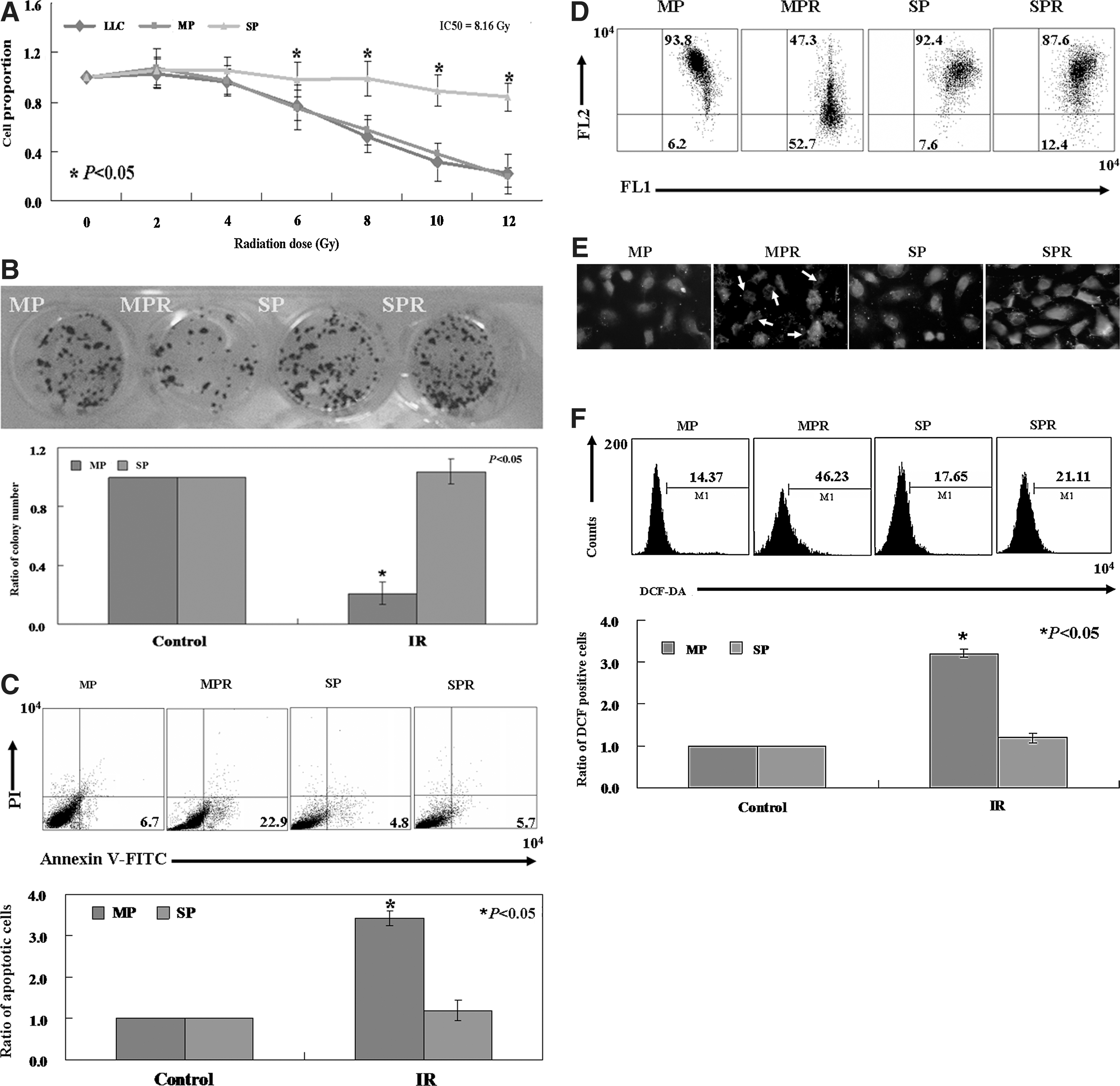
The effects of ionizing radiation (IR) on LLC SP and MP cells. (
To assess the role of mitochondria in IR-induced apoptosis
JC-1 is a potentiometric dye that exhibits membrane potential-dependent depletion as mitochondria transition from polarized to depolarized. The loss of membrane potential is indicated by the fluorescence emission shift from red to green. As shown in Figure 2D, the ratio of red/green in the MP cells (6.2% green, 93.8% red) was reversed following IR treatment (52.7% green, 47.3% red). However, there was no change in membrane potential of the SP cells. The results indicated that IR-induced apoptosis was associated with downregulation of MMP in the MP cells. Here we found that the level of cytochrome c was increased significantly in the cytosol of the MP cells, as assessed by immunofluorescence after IR treatment (Fig. 2E). We used the fluorescent dye DCF-DA to measure ROS content in MP and SP cells following IR. As shown in Figure 2F, IR directly induced an increase in fluorescence intensity in the MP cells (46.2%) as compared with untreated cells (14.4%) (p<0.05). We did not detect a change in fluorescent signal in SP cells exposed to the same radiation dose.
The mechanism(s) of IR-induced apoptosis in the MP cells
To identify whether IR-induced apoptosis in MP cells was mediated through the mitochondrial apoptotic pathway, we detected protein expression of Bax, Bcl-2, and Bcl-xl, which are members of the Bcl-2 family, using Western blot analysis. As shown in Figure 3, we found a decrease in Bcl-2 and Bcl-xl protein and an increase in Bax protein levels in MP cells after IR exposure (p<0.05). In previous studies, Bcl-2 has been shown to be inactivated via phosphorylation. 28 We noticed that the phosphorylation of Bcl-2 was increased in MP cells treated with IR (p<0.05, Fig. 3). It is well known that the caspase family plays a critical role in the apoptotic process. 29 Along with changes in cytochrome c, caspase-3 was activated by the cleavage of procaspase-3 into the active form. As shown in the left panel of Figure 3, active caspase-3 protein was significantly increased in MP cells following radiation treatment. As a downstream marker of active caspase-3, the cleavage of procaspase-9 was also increased (Fig. 3).

Effects of IR on mitochondrial apoptosis related proteins. Cellular protein was isolated from MP and SP cells treated with IR for 24 hours. Afterward, Western blot analysis was performed to detect Bcl-xl, Bcl-2, Bax, phosphor-Bcl-2, activated caspase 9, and caspase 3 proteins using the respective specific antibodies (right panel). The bar graph in the left panel shows Bcl-2/Bax and Bcl-xl/Bax expression ratios in MP and SP cells. * indicates a significant difference between the two cell types (p<0.05).
Discussion
ATP binding cassette transporters (ABCG) that can pump out Hoechst 33342 have been found in a relatively rare subgroup of many cancer cells. 30 Cancer cells possessing this property were named SP cells. In this study, we isolated SP cells (0.37% of the total cell population) from LLC cells. Consistent with previous studies, SP cells isolated from other cancer cells make up no more than 5% of the total cells. 31 By reverse transcription polymerase chain reaction (RT-PCR), Western blot, and immunofluorescence analysis, we identified the expression of ABCG2 in SP cells, but not in MP cells, providing further support that we have successfully isolated SP cells. While studies have reported that CD133 may not be increased in some types of CSC-like SP cells, 32 CD133 has been reported as a surface marker in SP cells in many types of cancers. 15 –19 Consistent with this, we found that CD133 was expressed in LLC SP cells, which suggested the isolated SP population cells.
We found that IR induced apoptosis in MP cells, but not in SP cells. Interestingly, loss of MMP is associated with IR-induced apoptosis in MP cells. We suspected that IR induced apoptosis via the mitochondrial-dependent caspase signaling pathway. The disruption of MMP has been reported to be affected by ROS. 33 ROS is formed by byproducts of the normal cellular metabolism of oxygen, such as singlet oxygen molecules and free radicals. 21 If the balance between ROS levels and the antioxidant level is disrupted, oxidative stress could be generated. An oxidative environment can lead to cellular structure damage, signal transduction changes, and cell death. 22 Consistent with this; we showed that the intracellular ROS level was increased in MP cells following IR exposure. The release of cytochrome c from the mitochondria into the cytosol, which is often a consequence of decreased MMP, is considered to be a significant initiator in the apoptosis process. 20 Our results showed that the cytochrome c level was increased significantly in the cytosolic fraction of MP cells treated with IR.
Previous studies have demonstrated that cytochrome c can bind to apoptotic protease activating factor 1, which is an activator of caspase-9 and caspase-3. 34 Consistent with our results, activated caspase-9 was increased in MP cells, along with the generation of cytochrome c. As a consequence of caspase-9 activation, procaspase-3 is cleaved into activated caspase-3. Expression changes in Bax and Bcl-2 are relevant to the mitochondria-dependent apoptotic pathway. 35,36 According to a previous study, Bcl-2 and Bcl-xl are two antiapoptotic proteins, and Bax is a pro-apoptotic protein. 37 The ratio of Bcl-2/Bax is critical for the induction of apoptosis. The balance of proapoptotic and antiapoptotic members controls the sensitivity of cells to apoptosis. 38 In line with the earlier findings, we observed reductions in both the Bcl-2/Bax and Bcl-xl/Bax expression ratios in MP cells treated with IR. Consistent with Adams' results, 28 we also found that p-Bcl-2, an inactivated form of Bcl-2, was increased significantly. Together, our results suggested that IR promotes apoptosis via a mitochondria-dependent caspase signaling pathway that included reduction of Bcl-2 and Bcl-xl proteins, and an induction in Bax protein and increased activation of caspase-3 and caspase-9. More studies are needed to further confirm this using the inhibitor blockade or silencing directed toward target proteins within the caspase signaling pathway listed above.
Radiation resistance of CSC-like SP cells is one possible reason of therapeutic resistance to IR. 39,40 In our studies, IR showed no effects on SP cells isolated from LLC cells. SP cells exhibited lower radiosensitivity than MP cells. High radiation resistance of SP cells in solid tumors has been demonstrated in breast cancer 37 and glioma tumors. 41 The reason for this resistance has remained unclear. Maynard et al. 42 showed that expression of DNA repair-DSBs genes were hyperstimulated in human embryonic stem cells after IR treatment. One recent study from López et al. 43 found increased clonogenic survival in human cervical cancer-initiating cells-enriched populations in which the homologous recombination mechanism and nonhomologous end joining mechanism repair pathways were activated with IR treatment. Another study demonstrated that the noncanonical Wnt signaling was involved in murine mesenchymal stem cells senescence induced by radiation exposure. 44 However, additional studies are required to elucidate the detailed mechanisms of LCC SP cell radioresistence.
SP cells are regarded as the root of cancer origin and recurrence. 6,7 Thus, new therapeutic approaches targeting these malignant cells have become a topic of ongoing research. For example, Notch signaling is highly active in SP cells compared with that of non-SP cells derived from nasopharyngeal carcinoma (NPC) cells. Therefore, Notch inhibition could reduce the function of SP cells, and may be a promising clinical approach in CSC-targeting therapy for NPC. 45 Apatinib, an oral multitargeted tyrosine kinase inhibitor, enhanced the efficacy of conventional anticancer drugs in the SP cells and ABCB1-overexpressing leukemia cells in vitro and in vivo. 46 In addition, Targeting miRNAs, such as miR-21, provides a new strategy for cancer therapy by impairing resistance to chemotherapy in CSC SP cells. 47
In summary, our results show that IR induces apoptosis via a mitochondria-dependent caspase signaling pathway that includes reduction of Bcl-2 and Bcl-xl proteins, and induction of Bax protein and increased activation of caspase-3 and caspase-9. However, the same IR dose has no effect on SP cells, suggesting that SP and MP cells display distinct radiosensitivity (Fig. 4). Future studies are needed to elucidate the mechanism by which SP cells resist radiotherapy, thereby looking for potential agents that can increase the response of LLC SP cells to radiation therapy.
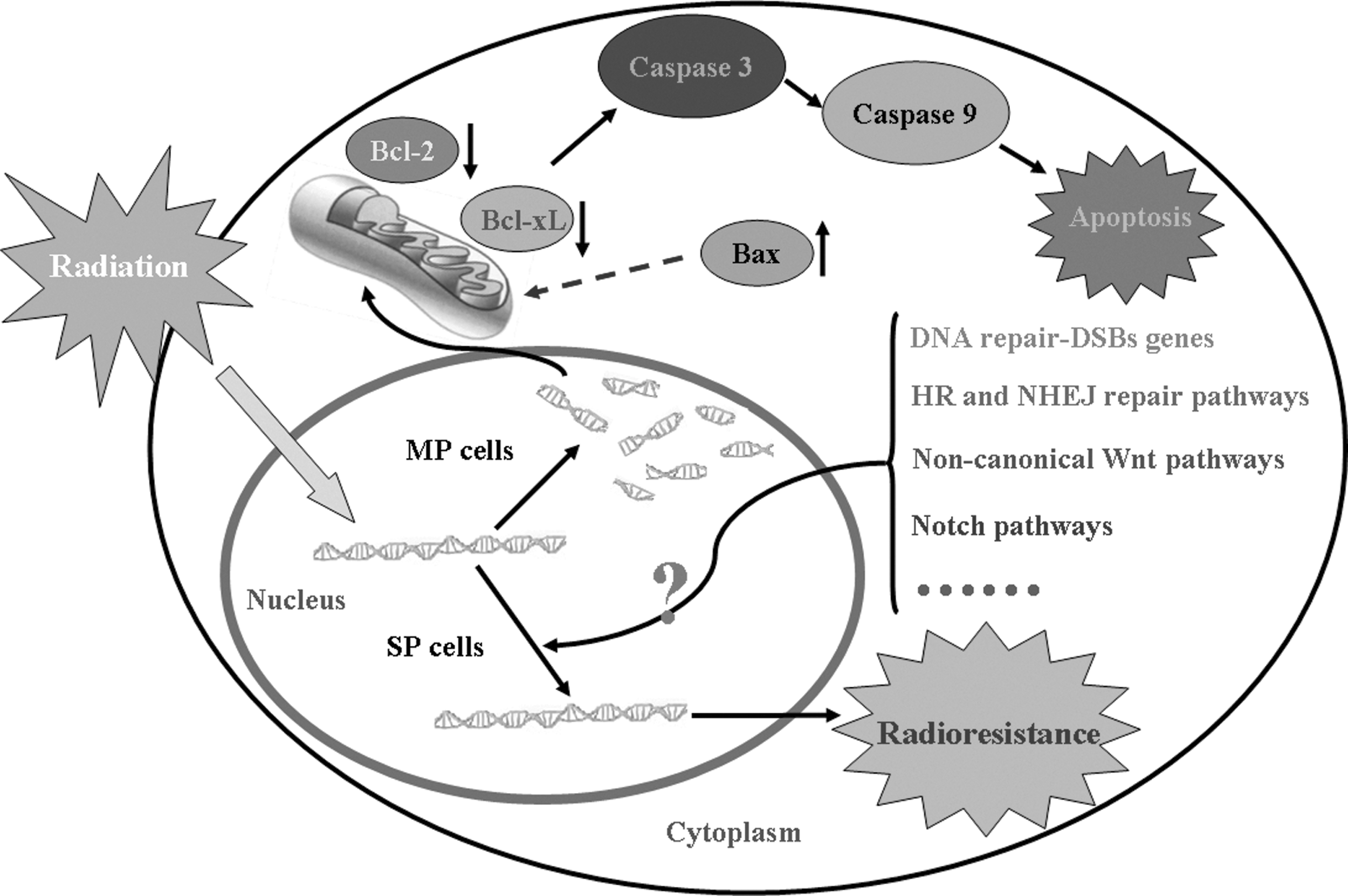
The proposed model of the apoptotic signaling pathways induced by IR in LLC MP and SP cells. The diagram shows IR induces apoptosis via a mitochondria-dependent caspase signaling pathway that includes reduction of Bcl-2 and Bcl-xl proteins, and an induction of Bax protein and increased activation of caspase-3 and caspase-9. However, the same dose of IR has no effect on the SP cells, suggesting that SP and MP cells have distinct radiosensitivity. Possible radioresistant mechanisms of SP cells are also proposed in the model. HR, homologous recombination mechanism; NHEJ, nonhomologous end joining mechanism.
Footnotes
Acknowledgments
This study was supported by grants from the Shenyang Outstanding Talent Foundation of China; the Shenyang Science and Technology (F11-264-1-10); the Natural Scientific Foundation of China (No. 81172371); and the Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports and Technology of Japan (23659958).
Authors' Contributions
P.X. performed the RT-PCR, luciferase assay, cultured cell lines, and performed the bioinformatics analyses. W.G., Z.N., and S.C. performed the western blot, Colony formation assay, and FCM. J.W. helped draft and revise the manuscript, and performed some of the imaging. Y.T. and H.Z. conceived the idea, drafted the manuscript, and supervised the experiments. All authors read and approved the final manuscript.
Disclosure Statement
The authors declare no conflicts of interest.