
Abstract
Epidermal growth factor receptor (EGFR) promotes proliferation of cancer cells. Dominant negative EGFR (DNEGFR) can block EGFR signal pathway by competing with endogenous EGFR for ligands. However, whether EGFR is overexpressed in gastric cancer and whether DNEGFR contributes to the inhibition of gastric cancer growth are not known. In this study, with the methods of immunohistochemistry, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay, flow cytometry, Terminal deoxynucleotidyl transferase mediated deoxyuridine triphosphate nick-end labeling assay and western blotting; we demonstrate that EGFR is expressed in 29 of 60 of human gastric cancer. In addition, DNEGFR induces G0/G1 arrest by decreasing expression of phosphorylated retinoblastoma protein, phosphorylated GSK-3β, cyclin D1, and by increasing expression of p21 and p27 in human gastric cancer cell lines SGC-7901 and NCI-N87. Finally, DNEGFR induces apoptosis in these cells. Our results indicate that DNFGFR may provide promising treatment strategy for a subgroup of human gastric cancers that express EGFR.
Introduction
Gastric cancer is the fourth most common cancer worldwide and is the second leading cause of death (700,000 deaths annually). The high-risk areas (with age-standardized rate, of >20 per 100,000 in men) include East Asia (China and Japan), Eastern Europe, and parts of Central and South America. 1 In spite of surgery, chemotherapy and radiotherapy, the prognosis of gastric cancer is still poor, with the overall 5-year survival rate ranging from 5% to 15%. 2
The disorder of cell proliferation and apoptosis induced by the activation of oncogenes and the inactivation of antioncogenes are two basic molecular biological events of tumorigenesis and tumor progression. 3 Epidermal growth factor receptor (EGFR) gene is a member of the ErbB family oncogenes. 4 EGFR protein is a 170-kDa protein consisting of an extracellular ligand-binding domain, a transmembrane lipophilic segment, and an intracellular tyrosine kinase domain. 5 Following stimulation by natural ligands, such as EGF or transforming growth factor-α, the EGFR changes in conformation, allowing it to form dimers with other oligomers. 6,7 Dimerization results in the activation of intracellular tyrosine kinase and protein phosphorylation and initiates signal transduction cascades which promote the cell division, invasion, and angiogenesis. 8 –10 The dominant negative EGFR (DNEGFR) is a truncated receptor without the tyrosine kinase domain, that competes with endogenous EGFR for natural ligands, leading to the inhibition of receptor activation. 11,12 In our previous study, we constructed a pEGFPN1-DNEGFR vector which carried a fusion gene encoding EGFR with a cytoplasmic-domain substituted by enhanced green fluorescent protein (EGFP). 13 The fusion protein was designated DNEGFR-EGFP, which provided a sensitive and convenient marker for direct monitoring of DNEGFR in cells. 14
It has been reported that DNEGFR enhances radiosensitivity of malignant glioma cell. 11 However, whether EGFR contributes to tumorigenesis and tumor progression of gastric cancer is unclear. Furthermore, whether DNEGFR has antiproliferative and antiapoptotic effects on human gastric cancer cells remains unanswered. In this study, we examined EGFR expression level in primary gastric cancer specimens and non-neoplastic gastric tissues, and analyzed its relationship with clinicopathological parameters. We also validated the effects of DNEGFR on cell cycle profile and apoptosis in two human gastric cancer cell lines, and investigated the underlying mechanisms.
Materials and Methods
Collection of tissue specimens and EGFR immunohistochemistry
A total of 60 primary gastric cancer tissues and their corresponding nonmalignant gastric tissues were collected randomly from Department of Gastrointestinal Surgery, the First Affiliated Hospital of Chongqing Medical University (Chongqing, China). Relevant clinicopathological parameters were collected simultaneously. Tissue specimens were routinely fixed in 10% formalin, embedded in paraffin. Tissue specimens were sectioned (5 μm thick). The tissue sections were placed on glass slides, and heated at 58°C for 2 hours to let them be firmly attached to glass slides.
The tissue sections were deparaffinized and rehydrated according to standard histological techniques. After being covered with Protease XXV solution (Thermo Fisher Scientific, Rockford, IL) and digested for 10 minutes at 37°C for antigen retrieval, the tissue sections were incubated in 3% H2O2 to block endogenous peroxidase activity. The primary monoclonal mouse anti-human EGFR antibody (1:100 dilution; Thermo Fisher Scientific) was applied to tissue sections, and the sections were further incubated for 1 hour in a humidified chamber at 37°C. The sections were washed thrice with phosphate buffered saline (PBS) and incubated in Polymer Helper (Zhongshan, Beijing, China) for 20 minutes. After washing, the sections were incubated in a polyperoxidase-anti-mouse IgG (Zhongshan) for 30 minutes at room temperature. At last, the sections was developed in 3-3′-diamino-benzidinetetrahydro chloride (DAB; Sigma-Aldrich, St. Louis, MO) solution, counterstained with haematoxylin (Sigma-Aldrich), dehydrated, and mounted in neutral balsam. Negative controls were included with the primary antibody being substituted with PBS, and positive controls were included using EGFR-positive breast cancer tissue sections. EGFR expression was evaluated according to a previously-described method. 15 All these procedures were approved by the Ethics Committee of Human Experimentation in Chongqing, and in accordance with the Helsinki Declaration of 1975.
Cell lines and cell culture
Human gastric cancer cells SGC-7901 were obtained from the American Type Culture Collection (Manassas, VA); cells NCI-N87 were purchased from Shanghai Institute of cells, Chinese Academy of Sciences (Shanghai, China). Cells were grown in RPMI-1640 medium (Invitrogen, Carlsbad, CA) containing 10% fetal bovine serum (Invitrogen), incubated at 37°C in a humidified atmosphere 5% CO2/95% air.
Transfection of plasmids
The vector pEGFPN1-DNEGFR carrying a fusion gene encoding DNEGFR-EGFR was constructed in our previous work. 13 Mediated by Lipofectamine 2000 (Invitrogen), SGC-7901, and NCI-N87 cells were transfected with the plasmids pEGFP-N1 (Clontech, Mountain View, CA) or pEGFPN1-DNEGFR according to the manufacture's protocol. Cells stably transfected with vectors were selected with G418 (SGC-7901 cells: 350 μg/mL, NCI-N87 cells: 500 μg/mL; Invitrogen) 48 hours after transfection, and individual clones were isolated, expanded, and maintained with G418 (SGC-7901 cells: 100 μg/mL, NCI-N87 cells: 250 μg/mL) for analysis. The cells were grouped as US or UN (untreated SGC-7901 or NCI-N87 cells), ES or EN (EGFP-expressing SGC-7901 or NCI-N87 cells, which were stably transfected with pEGFP-N1), and DS or DN (DNEGFR-expressing SGC-7901 or NCI-N87 cells, which were stably transfected with pEGFPN1-DNEGFR).
MTT assay
Log-phase cells were seeded in 96-well plates (5×103 cells/well) for 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay. Each group was analyzed at indicated times from 1 to 3 day in triplicate by the following method: cells were incubated with MTT solution (5 mg/mL, 20 μL/well; Sigma-Aldrich, St. Louis, MO) at 37°C for 4 hours, and following the removal of MTT solution. The formazan crystals were solubilized in 150 μL of dimethylsulphoxide (DMSO; Sigma-Aldrich). Subsequently, the absorbance (A) value was measured at a wavelength of 570 nm on an ELISA plate reader (Bio-Rad Laboratories, Hercules, CA) with DMSO as the blank. The growth curve of cells was plotted with time (day) as x-axis and A value as y-axis.
Flow cytometry
Cells were detached with 0.1% trypsin/0.02% EDTA (Sigma-Aldrich), washed by PBS thrice, fixed with cold 70% ethanol at 4°C overnight, then washed and resuspended in PBS containing RNase A (Sigma-Aldrich) and stained by propidium iodide (PI; Sigma-Aldrich) at room temperature for 30 minutes, and then PI-stained cells were detected on a flow cytometer (BD Biosciences, San Jose, CA). Cell cycle distribution was analyzed using the Modifit-3 program (BD Biosciences).
TUNEL assay
Apoptosis index (AI) was assessed by Terminal deoxynucleotidyl transferase mediated deoxyuridine triphosphate nick-end labeling (TUNEL) assay. Log-phase cells were cultured in 24-well plates (1.5×104 cells/well) for 48 hours. For detection of apoptosis, TUNEL assay was performed using the in situ cell death detection kit (Roche Applied Science, Nonnenwald, Germany) according to the manufacture's protocol. In brief, after fixation and permeabilisation, cells were incubated with TUNEL reaction mixture at 37°C for 60 minutes in a humidified atmosphere in the dark, and then incubated with Converter-POD in a humidified chamber at 37°C for 30 minutes. Subsequently, cells were developed in DAB (Sigma-Aldrich) solution, counterstained with haematoxylin (Sigma-Aldrich). At last, stained cells were analyzed under light microscope. For each sample, five randomly selected fields at ×200 magnification were evaluated, and the AI was calculated according to the formula: 100×the number of positive cells/the number of all visual cells. 16
Western blot analysis
Western blot analysis was used to investigate EGFR, Phospho-EGFR (Tyr 1173), retinoblastoma protein (Rb), Phospho-Rb (Ser 780), p21, p27, cyclin D1, and pGSK-3β (Ser 9) at the protein level. Cells were washed once with ice-cold PBS, then lysed in lysis buffer [50 mM Tris (pH, 7.5), 100 mM NaCl, 1 mM EDTA, 0.5% NP40, 0.5% Triton X-100, 2.5 mM sodium orthovanadate, 10 μL/mL protease inhibitor cocktail, and 1 mM phenylmethylsulfonyl fluoride] for 20 minutes on ice and centrifuged. The protein concentration of the clarified lysate was quantified by BCA protein assay kit (Thermo Fisher Scientific, Rockford, IL). Protein samples (50 μg) were subjected to sodium dodecyl sulfate/10% polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Bio-Rad Laboratories). Blots were blocked in Phosphate Buffered Saline plus Tween 20 (PBST) with 1% bovine serum albumin, incubated with the primary antibodies to Phospho-Rb (Ser 780), pGSK-3β (Ser 9) (Phospho-Rb:dilution 1:1000, pGSK-3β: dilution 1:1200; Cell Signaling Technology, Danvers, MA) cyclin D1, p27 (dilution 1:100; Santa Cruz Biotechnology, Santa Cruz, CA), p21, GAPDH, EGFR (C-20), p-EGFR (Tyr 1173), Rb (C-15) (dilution 1:200; Santa Cruz Biotechnology) at 37°C for 1 hour, and washed and incubated with a goat anti-rabbit antibody conjugated horseradish peroxidase (dilution 1:10000; Santa Cruz Biotechnology) at 37°C for 1 hour. Specific antibody-labeled proteins on the membranes were detected using Pierce electrochemiluminescence (ECL) Substrate (Thermo Fisher Scientific) and visualized on Gel Doc XR System (Bio-Rad Laboratories) according to the manufacturer's instructions. Each experiment was repeated three times. Densitometry was utilized to further assess each lane.
Statistical analysis
Comparison of EGFR expression between gastric cancer tissues and their corresponding nonmalignant gastric tissues, correlation between EGFR expression in gastric cancer tissues, and clinicopathological parameters were investigated with the chi-square test. For other statistical comparison, statistical differences were evaluated by one-way analysis of variance followed by least significance difference (LSD) multiple comparison tests. All statistical tests were performed with SAS Software (SAS Institute, Inc, Cary, NC). p<0.05 was used to indicate statistical significance, and p values were given for two-sided.
Results
Overexpression of EGFR in human gastric cancer
EGFR expression level was tested in 60 gastric cancer tissues and their corresponding nonmalignant gastric tissues. A diffuse and intense plasmalemmal and cytoplasmic staining pattern for EGFR was detected in gastric cancer tissues, whereas the corresponding nonmalignant gastric tissues showed weak or no staining (Fig. 1). 29 (48.3%) gastric cancer tissues demonstrated enhanced expression of EGFR protein, 4 (6.7%) nonmalignant gastric tissue demonstrated weak expression of EGFR protein (p<0.05), suggesting its involvement in tumorigenesis of gastric cancer.
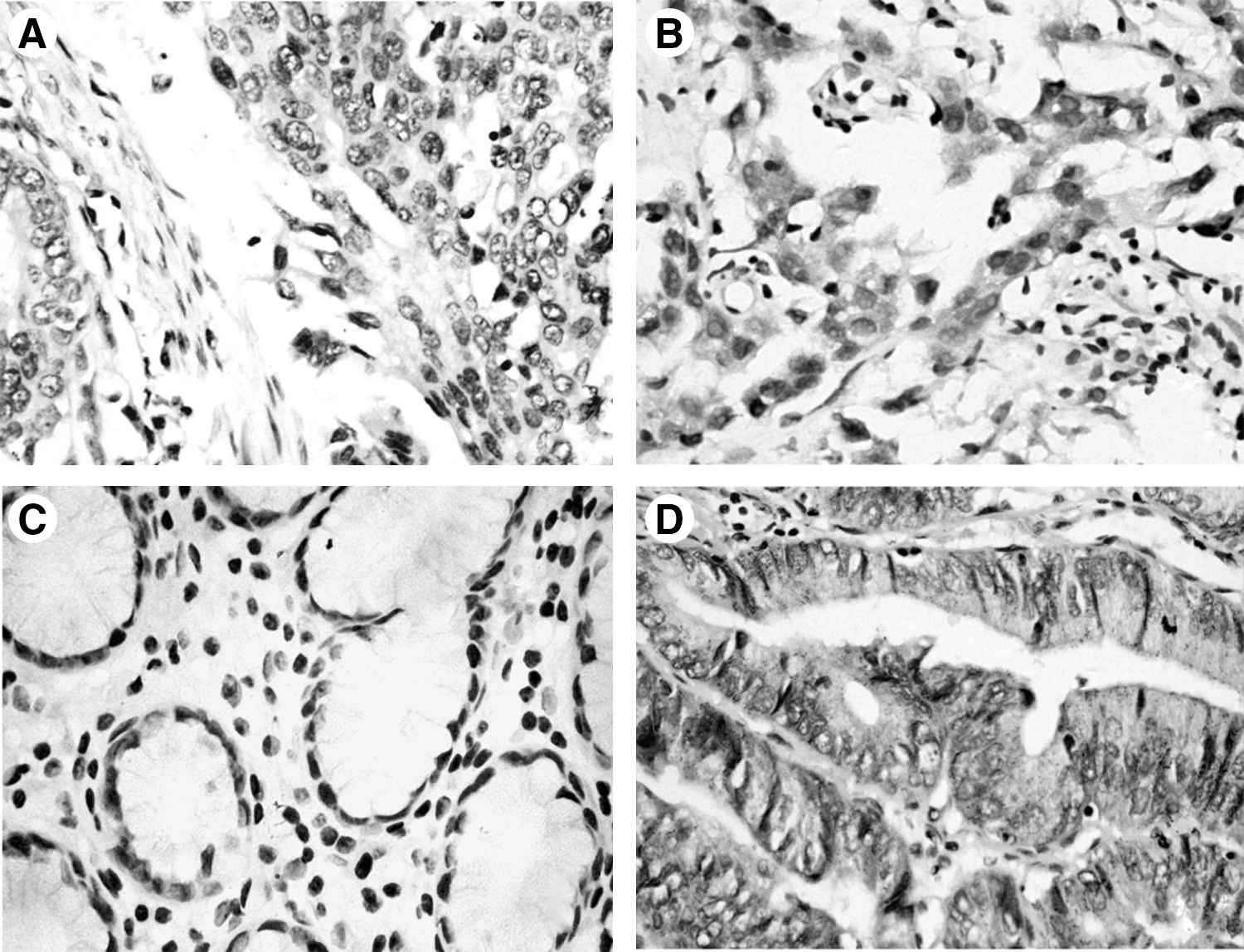
Overexpression of epidermal growth factor receptor (EGFR) in human gastric cancer tissue.
The relationship between the expression of EGFR in gastric cancer tissues and clinicopathological parameters was showed in Table 1. The positive rate of EGFR staining in gastric cancer tissues did not correlate with gender (p>0.05), age (p>0.05), tumor size (p>0.05), tumor location (p>0.05), differentiation (p>0.05). A significant correlation was seen for depth of invasion, where the positive rate of T3 and T4 was higher than that of T1 and T2 (p<0.05). A significant correlation was also seen for lymph node metastasis, where the positive rate with lymph node metastasis was higher than that without lymph node metastasis (p<0.05). Additionally, there was a significant correlation of EGFR positive rate and tumor-node-metastasis (TNM) stage: The positive rate in TNM stage III and IV was higher than that in TNM stage I and II (P<0.05). Our results were consistent with the previous reports, 17,18 suggesting its involvement in tumor progression of gastric cancer.
DNEGFR decreased phosphorylation of EGFR
As showed in Figure 2, the phosphorylation of EGFR at Tyr 1173 in groups DS and DN decreased, compared with groups US and ES, groups UN and EN, respectively (p<0.05); while no difference was observed in the expression of total EGFR protein. DNEGFR-EGFP was a fusion protein of DNEGFR and EGFP, acted as a dominant negative receptor, competed with endogenous EGFR for EGF in complete medium, resulted in the inhibition of intracellular tyrosine kinase and protein phosphorylation, leading to EGFR signaling pathway blockade at last. We named the EGFR signaling pathway blockade strategy as dominant negative strategy targeting EGFR.
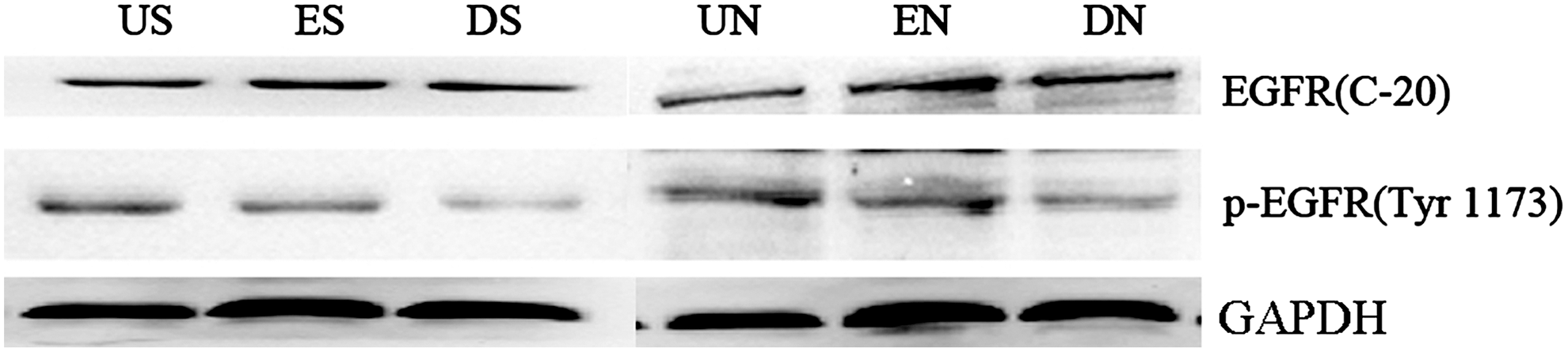
Dominant negative EGFR (DNEGFR) decreased phosphorylation of EGFR. Western blot analysis of total EGFR and the phosphorylation of EGFR at Tyr 1173 was performed in SGC-7901 and NCI-N87 cells. US or UN (untreated SGC-7901 or NCI-N87 cells), ES or EN (SGC-7901 or NCI-N87 cells, stably transfected with pEGFP-N1), and DS or DN (SGC-7901 or NCI-N87 cells, stably transfected with pEGFPN1-DNEGFR).
DNEGFR inhibited cell growth
The effect of DNEGFR on the growth of SGC-7901 and NCI-N87 cells was evaluated using the MTT assay. The MTT assay indicated that DNEGFR inhibited the growth of cells in vitro (p<0.05) (Fig. 3). The growth of cells in vitro is the overall effects of cell cycle and apoptosis. However, whether DNEGFR inhibits the growth of cells in vitro by leading to proliferation inhibition or/and inducing apoptosis is unknown. To explore the reason for cell growth inhibition, cell proliferation and apoptosis were further tested.
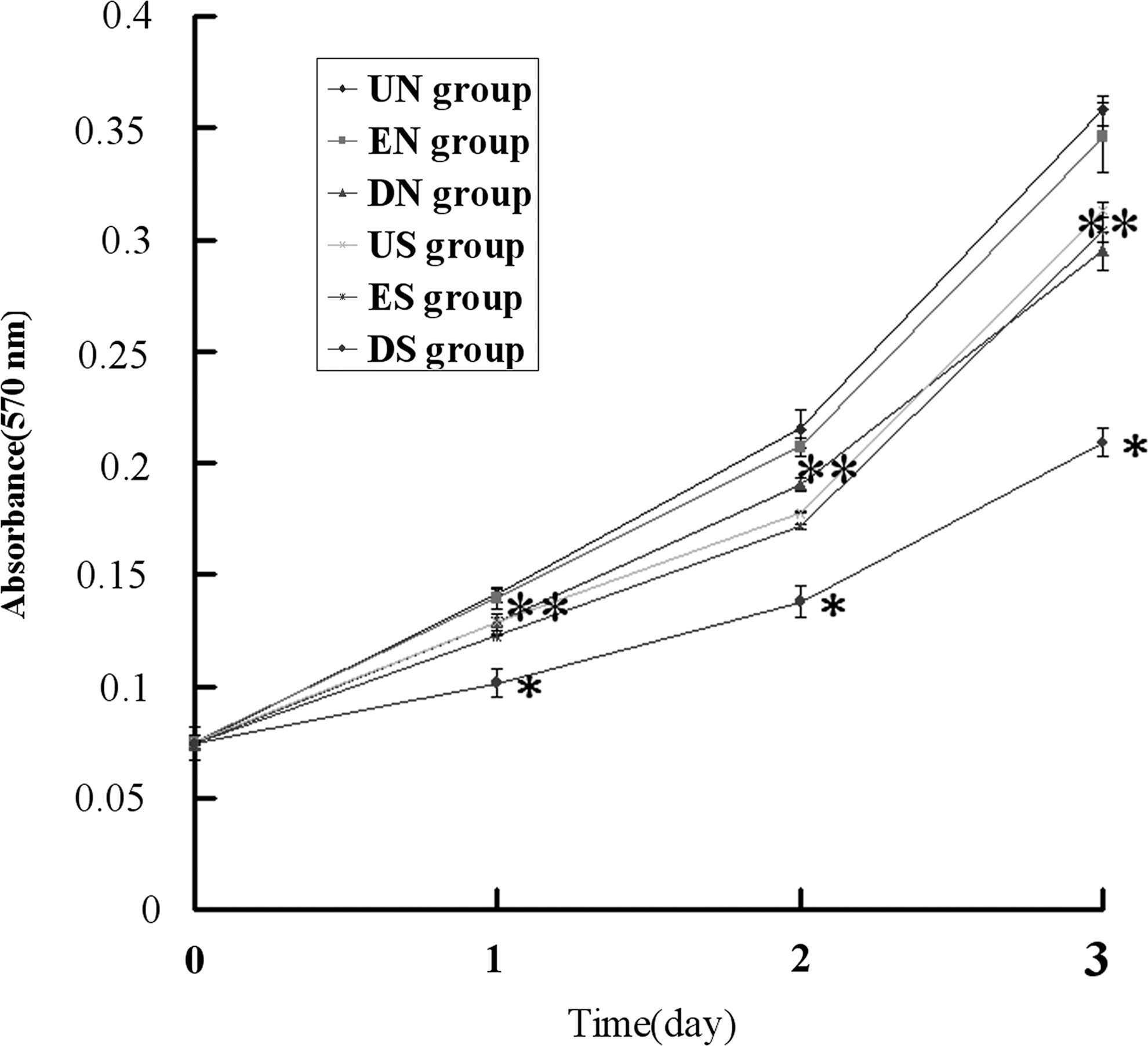
DNEGFR inhibited cell growth in vitro. Growth curves of cells plotted by the MTT assay. Each group was analyzed every day in triplicate for 24–72 hours. Each data point was the mean±SD from three independent experiments; bars, SD. *p<0.05 compared with US and ES groups, **p<0.05 compared with UN and EN groups.
DNEGFR led to G0/G1 arrest
Cell proliferation is regulated by cell cycle, and G0/G1-S transition is a critical checkpoint that acts as a switch dedicating the cell to DNA replication. 19 To test the effect of DNEGFR on cell cycle progression of SGC-7901 and NCI-N87 cells, cell cycle assay by PI staining and flow cytometry was performed. The G0/G1 phase fractions in US and ES groups were 50.03%±2.01%,49.61%±0.49%, respectively, and increased to 70.88%±0.85% in DS group; the S phase fractions in US and ES groups were 43.63%±1.26%, 43.63%±0.64%, respectively, and decreased to 21.58%±1.40% in DS group (p<0.05) (Fig. 4A and B). The G0/G1 phase fractions in UN and EN groups were 47.90%±2.52%, 48.79%±4.12%, respectively, and increased to 69.27%±4.52% in DN group; the S phase fractions in UN and EN groups were 41.77%±3.08%, 42.76%±3.38%, respectively, and decreased to 21.88%±2.40% in DN group (p<0.05) (Fig. 4A and B). These results indicate that DNEGFR prevents cell cycle progression through G0/G1 to S phase in SGC-7901 and NCI-N87 cells, leading to G0/G1 arrest.
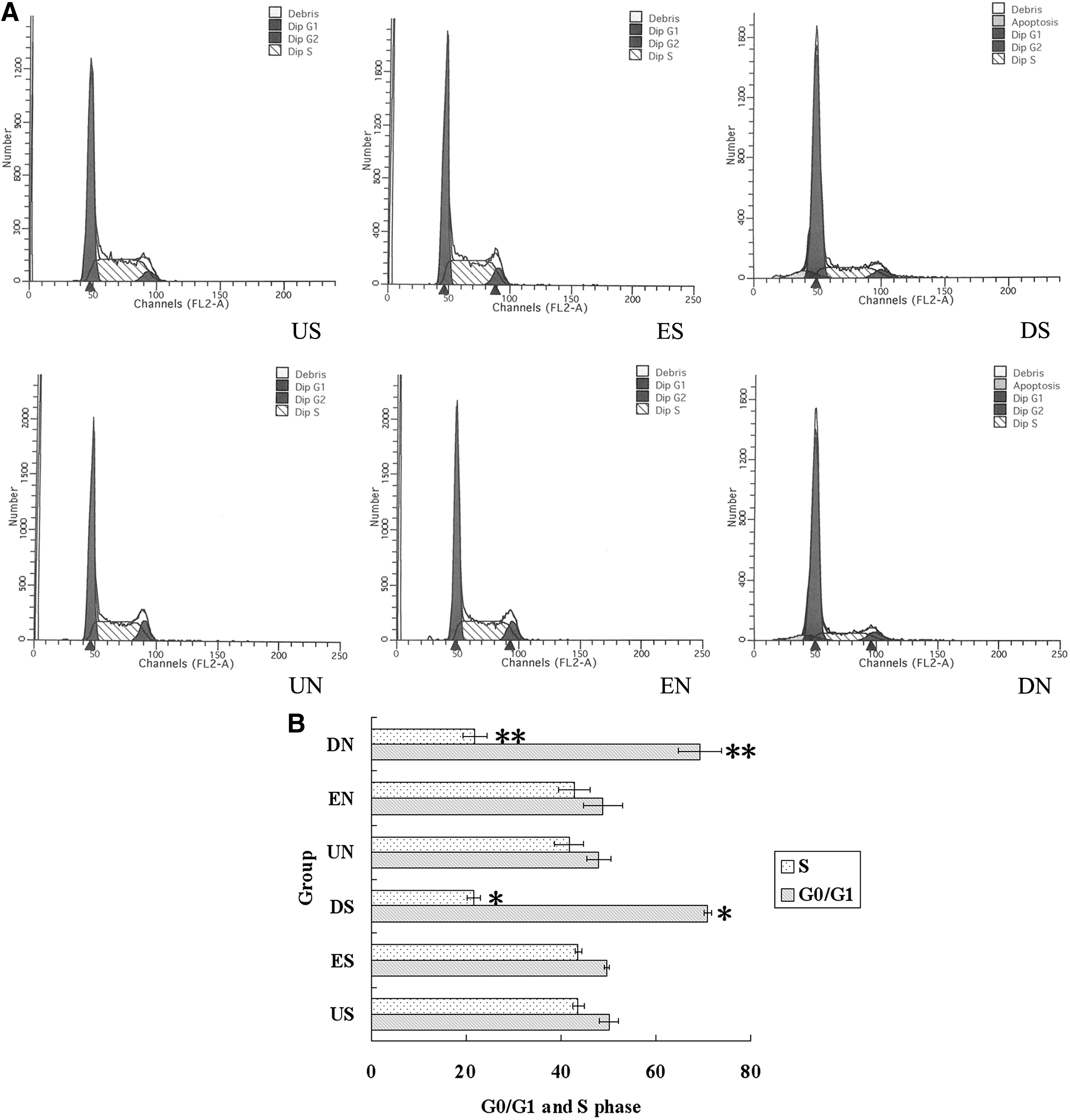
DNEGFR induced G0/G1 arrest.
DNEGFR decreased phosphorylation of Rb
The final common pathway governs G0/G1-S transition is the cyclin-dependent kinases (CDKs) induced hyperphosphorylation of the Rb. 20,21 Hypophosphorylation of Rb is active, inhibits transcriptional activity of transcription factors by binding to them. Hyperphosphorylation of Rb leads to the release of transcription factors, especially E2F, which induce the synthesis of relative proteins required for the G0/G1-S transition. 22 Therefore, we examined the effect of DNEGFR on the phosphorylation of Rb at Ser 780. As showed in Fig 5, phosphorylation of Rb in groups DS and DN decreased notably, compared with groups US and ES, groups UN and EN, respectively (p<0.05); while no difference was observed in the expression of total Rb protein. These results indicate that DNEGFR decreases the phosphorylation of Rb at Ser 780, which results in G0/G1 arrest in SGC-7901 and NCI-N87 cells.
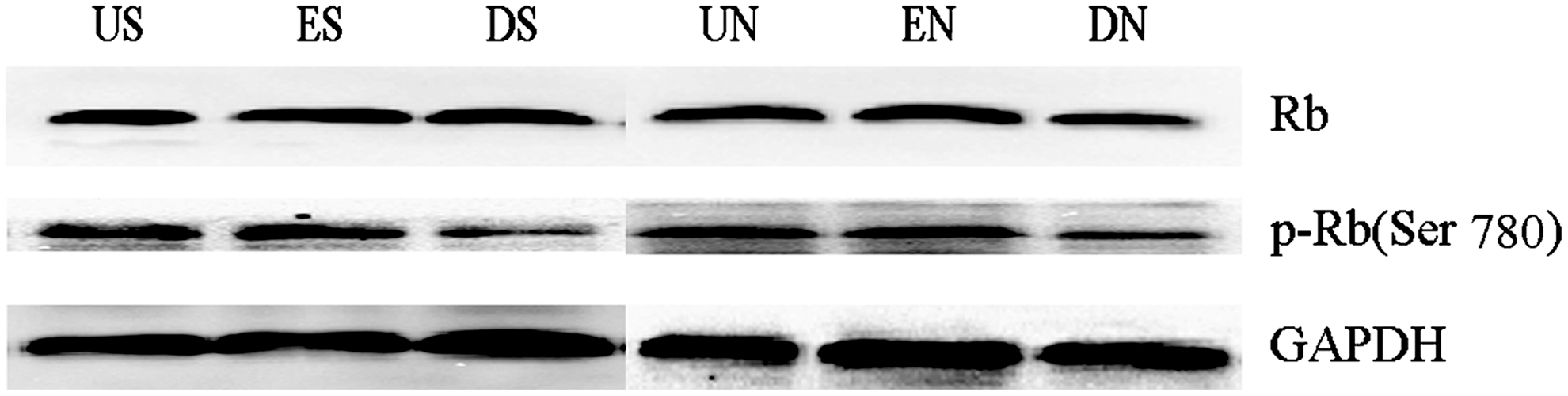
DNEGFR decreased phosphorylation of retinoblastoma protein (Rb). Western blot analysis of total Rb and the phosphorylation of Rb at Ser 780 was performed in SGC-7901 and NCI-N87 cells.
DNEGFR increased p21 and p27
Rb phosphorylation and subsequent cell cycle progression likely requires first phosphorylation by cyclin D1-CDK4/6. 21,23 –25 p21 and p27 are two critical CDK inhibitors, play an important role in cell cycle. They associate with and inhibit cyclin/CDK holoenzymes containing either CDK4 or CDK6. 26,27 We postulated that DNEGFR decreased Rb phosphorylation by up-regulating the expression of p21 and p27, and therefore tested p21 and p27 at protein level. As illustrated in Figure 6, p21 and p27 in groups DS and DN increased notably, compared with groups US and ES, groups UN and EN respectively (p<0.05). We think that the upregulations of p21 and p27 are partly responsible for the reduction of Rb phosphorylation in SGC-7901 and NCI-N87 cells.
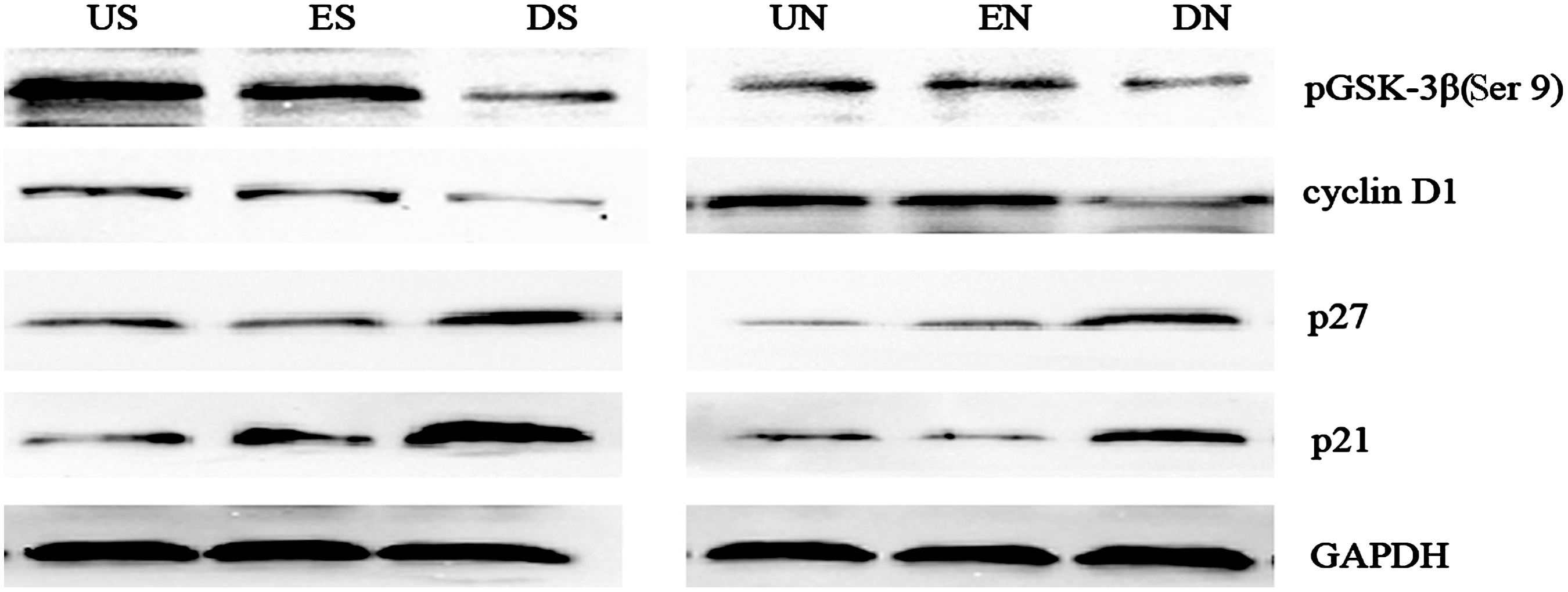
DNEGFR decreased p21 and p27, and decreased cyclin D1 by inhibiting the phosphorylation of pGSK-3β at Ser 9.
DNEGFR decreased cyclin D1 by inhibiting phosphorylation of pGSK-3β (Ser 9)
Cyclin D1 is a key regulator of cell-cycle progression, particularly at the transition from G0/G1 to the S phase. 28,29 Cyclin D1 overexpression was found in parts of gastric cancer tissue, reflected the severity of gastric cancer, and it played an essential role in proliferation and tumorigenicity of human gastric cancer cells. 30,31 Rb phosphorylation and subsequent cell cycle progression likely requires first phosphorylation by cyclin D1-CDK4/6. 21,23 –25 We postulated that DNEGFR partly decreased Rb phosphorylation by downregulating the expression of cyclin D1, and therefore, tested cyclin D1 at protein level. Cyclin D1 in groups DS and DN decreased notably, compared with groups US and ES, groups UN and EN, respectively (Fig. 6) (p<0.05). These results indicate that, besides the upregulations of p21 and p27, the downregulation of cyclin D1 are partly responsible for the reduction of Rb phosphorylation in SGC-7901 and NCI-N87 cells.
Cyclin D1 is partly under the control of pGSK-3β. pGSK-3β is activated by phosphorylating at Ser 9, 32 and active pGSK-3β phosphorylates cyclin D1 and leads to the degradation of cyclin D1 in the end. To further elucidate the mechanisms of cyclin D1 reduction, we examined the effect of DNEGFR on phosphorylated pGSK-3β at Ser 9. The phosphorylation of pGSK-3β at Ser 9 in groups DS and DN decreased notably, compared with groups US and ES, groups UN and EN, respectively (Fig. 6) (p<0.05). On the basis of these results, we speculate that DNEGFR results in local EGF withdrawal around cell membrane by competing with endogenous EGFR for EGF, and EGF withdrawal activates pGSK-3β by phosphorylating pGSK-3β at Ser 9, leading to cyclin D1 reduction at last.
DNEGFR induced apoptosis
As illustrated in Figure 7A, cells with brown nuclei are apoptotic cells, cells with blue nuclei are negative cells. The AIs in US and ES groups were 2.3%±0.6%,3.0%±1.0%, respectively, and increased to 10.3%±1.2% in DS group; the AIs in UN and EN groups were 1.3%±0.3%,2.0%±0.6%, respectively, and increased to 8.7%±0.6% in DN group (Fig. 7B) (p<0.05). These results indicate that DNEGFR leads to apoptosis in SGC-7901 and NCI-N87 cells. It was reported that pGSK-3β had proapoptotic effect. Since we proved that DNEGFR activated pGSK-3β in SGC-7901 and NCI-N87 cells, we think that pGSK-3β activated by DNEGFR is at least in part responsible for apoptosis in SGC-7901 and NCI-N87 cells. Whereas the concrete molecular mechanisms for the pro-apoptotic effect of pGSK-3β are still unknown, further studies are needed to elucidate the mechanisms.

DNEGFR induced apoptosis. Apoptotic index was assessed by Terminal deoxynucleotidyl transferase mediated deoxyuridine triphosphate nick-end labeling (TUNEL) assay.
Discussion
In this study, firstly, EGFR expression level was tested in gastric cancer tissues and their corresponding nonmalignant gastric tissues, the results suggest EGFR plays an important role in tumorigenesis and tumor progression of gastric cancer. Therefore, EGFR signaling pathway blockade was a promising therapeutic approach for a subgroup of gastric cancer overexpressing EGFR. Among EGFR signaling pathway blockade strategies, monoclonal antibodies and tyrosine kinase inhibitors targeting EGFR have been widely accepted as cancer therapy strategies in clinical practice 33 –36 ; however, their relative side effects hinder the application of them to some patients. As a type of EGFR signaling pathway blockade strategies, antisense technology targeting EGFR is a hotspot in cancer biotherapy. 37,38 Whereas, due to its drawbacks, such as off-target effect of small interfering RNA, instability and potential toxicity of antisense oligonucleotides in vivo, have limited the translation of this technology to the clinical setting. 39 –41 Antisense technology that aims at mRNA, 42,43 doesn't inhibit the function of protein that has been synthesized. Compared with antisense technology, dominant negative strategy targeting EGFR aims at EGFR protein inhibit the function of EGFR protein that has been synthesized, which is an advantage of it. Till now, there are few reports about dominant negative strategy targeting EGFR in gastric cancer biotherapy.
Therefore, secondly, multiple cellular and molecular approaches were used to evaluate the effects of DNEGFR on proliferation and apoptosis in two EGFR-overexpressing human gastric cancer cell lines, SGC-7901 and NCI-N87, and to elucidate the possible mechanisms behind them. MTT assay shows DNEGFR inhibits the growth of the two cell lines. Our study reveals that DNEGFR inhibits cell growth by leading to G0/G1 arrest and inducing apoptosis. Western blot analysis discloses that DNEGFR decreases cyclin D1 by inhibiting the phosphorylation of pGSK-3β (Ser 9), and by increasing p21 and p27 expression, leading to the reduction of phosphorylation of Rb, which result in G0/G1 arrest; and the down-regulation of phosphorylation of pGSK-3β (Ser 9) is at least partly responsible for apoptosis in the two cell lines.
One major limitation of this study is that the effects of DNEGFR were just evaluated in two EGFR-overexpressing gastric cancer cell lines, and the conclusions based on two cell lines may not be generally applicable to all human gastric cancer cell lines. Moreover, several open questions remain to be answered in future research. For these human gastric cancer cell lines coexpressing EGFR and other ErbB family receptors, what is the function of DNEGFR? As the most common rearranged form of EGFR, EGFR type III variant (EGFRvIII) has been identified in nonsmall cell lung cancer, glioblastomas, and breast tumors. 32,44,45 Is EGFRvIII expressed in human gastric cancer tissue and current human gastric cancer cell lines? If so, what are the effects of DNEGFR on EGFRvIII-expressing human gastric cancer cell lines?
In summary, EGFR is overexpressed in a subgroup of human gastric cancer. In EGFR expressing cell lines of human gastric cancer, DNEGFR inhibits growth by inducing cell cycle arrest and apoptosis. These results suggest DNFGFR may provide promising treatment strategy for a subgroup of human gastric cancers that express EGFR.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (No.30972872). The authors thank Professor Weixue Tang (Department of Pathophysiology, Chongqing Medical University) for her excellent technical assistance.
Disclosure Statement
No competing financial interests exist.