
Abstract
Hepatocellular carcinoma (HCC) is one of the most common malignant gastroenterological cancers over the world. α-fetoprotein (AFP) is an oncofetal protein produced during HCC development that could generate weaker and less reproducible antitumor protection, and it may serve as a target for immunotherapy. Therefore, it is imperative to enhance its immunogenicity and develop therapeutic vaccines to eliminate AFP-expressing tumors. In this study, by way of glutaraldehyde cross-linking, we constructed a potential therapeutic protein vaccine, glycoprotein 96 (gp96)/AFP. Our results demonstrated that AFP and gp96 synergistically exhibited significant increase in AFP-specific CD8+ T-cell responses and impressive cytotoxic antitumor effect against AFP-expressing tumors. Priming mice with the reconstructed vaccine, we elicited robust strong protective immunity. Our study suggests that tumor vaccine by cross-linking tumor antigen and gp96 is a promising approach to cancer therapy.
Introduction
Hepatocellular carcinoma (HCC) is one of the most common malignant gastroenterological cancers over the world and accounts for as many as 1.2 million deaths annually. 1 The highest incidence is in Southeastern and Eastern Asia, with a rate of 18.3–35.5 per 100,000 populations. 2 It is also rapidly rising in China because of hepatitis B and C infections. 1,2 Established treatments of HCC, such as chemo- and radio-therapy, usually fail to prevent the disease progress due to the proliferation of residual malignant cells that have escaped control by these methods. Although surgery and liver transplantation are the effective therapy, most patients still lose chance due to diagnosis at a late stage or underlying liver insufficiency in the setting of cirrhosis. 3 The most common causes of mortality in patients with HCC include recurrence, metastasis, and development of new primary tumors. 3 Novel therapies for HCC should be developed. A combined therapy is likely to prolong patients' lives and living quality. In recent years, immunotherapy with therapeutic vaccination, which could manipulate and enhance the host's immune system to elicit antigen-specific responses to the tumors, has widely attracted attention.
Much attention is on tumor active immunity where the purpose is to induce hosts' immune attack on tumor cells. 80% of HCC have a high expressing rate of α-fetoprotein (AFP), which may serve as a target for immunotherapy. 3,4 AFP is an oncofetal protein during HCC development which could generate weaker and less reproducible antitumor protection. Therefore, a vaccine specifically targeting AFP is particularly desirable. AFP oncogenic proteins are critical to the induction and maintenance of cellular transformation, which represent ideal target antigens for therapeutic AFP vaccine development. 5 Many therapeutic strategies to enhance the pre-existing antitumor immunity have been examined including recombinant proteins, 6 peptides, 7 plasmid DNA, 8 chimeric virus-like particles, 9,10 and viral or bacterial vectors expressing AFP proteins, 11 and adoptive transfer of tumor-specific T cells. 4,5 However, therapeutic vaccination has been restricted by inadequate antigen-specific immune responses. Therefore, ways to develop more potent immunotherapy aimed at initiating very robust antigen-specific immune responses need to be thoroughly explored.
Heat shock proteins (HSPs) have been verified to act as potent immunoadjuvant to enhance antigen-specific tumor immunity. 12 HSPs play important roles, not only in the process of protein folding, transport, and degradation, but also in directing more efficient antigen presentation to CD8+ T cells through the MHC-I pathway. 12 This central role for HSPs in therapeutic vaccine is facilitated by their dual functions in both chaperoning antigens and serving as an adjuvant. HSPs are classified into families according to their size, structure, and function, including small HSP, HSP60, HSP70, HSP90, glycoprotein 96 (gp96), and calreticulin (CRT). They have different structures that are adapted to different immunoadjuvant functions. 12,13 Specifically, HSP90 and HSP70, the most abundant cytosolic chaperones, have been proposed to transport peptides to the heterodimeric transporters associated with antigen processing (TAP1 and TAP2), whereas gp96 and CRT, the major chaperone of the lumen of the endoplasmic reticulum (ER), have been presumed to facilitate assembly of the MHC I—β-2 microglobulin—peptide complexes in the ER. 13,14 Further, HSP-based tumor vaccine strategies have been highly successful in animal models and are undergoing testing in clinical trial. 10,15,16 Therefore, an innovative approach that combines with families of HSPs will be likely to generate more potent antitumor effect.
A number of groups have shown that superior levels of T-cell immunity could be generated using a heterogeneous prime-boost strategy, in which animals are primed and boosted with a plasmid vector encoding the stimulating molecule and targeted peptides. 8 –10 In many of these vaccine models, 17,18 gp96 combined with certain antigen prime enhanced immunogenicity, presumably through processing and presenting the antigen to host antigen-presenting cells (APCs). In the present study, we investigated whether the immunogenicity of AFP could be improved by gp96 molecules and whether gp96 could synergistically enhance the potency of AFP therapeutic effect, and then the immune responses induced by gp96/AFP were further evaluated. We constructed a protein vaccine containing the molecule chaperon-gp96 and AFP protein by way of glutaraldehyde cross-linking. Our results demonstrated that AFP and gp96 synergistically exhibited significant increase in AFP-specific CD8+ T-cell responses and impressive antitumor effect against AFP-expressing tumors. Priming mice with the reconstructed vaccine, we elicited robust strong protective immunity. Therefore, cross-linking gp96/AFP protein holds promise for treating HCC through a combination of antigen-specific immunotherapy.
Materials and Methods
Mice and tumor cell lines
Six- to eight-week-old female BALB/C mice were purchased from the Experimental Animal Center at Xian Jiaotong University. All animals were maintained under specific pathogen-free conditions, and all procedures were performed according to approved protocols and in accordance with recommendations for the proper care of laboratory animals. The investigation was approved by the Ethics Committee on animal Study at Shaanxi University of Chinese Medicine (2004-4B). AFP-producing H22 mice HCC cells were kindly provided by the College of Biomedicine, Xi'an Jiaotong University and maintained in RPMI 1640 (Gibco-BRL) with 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin (Invitrogen Corp.) at 37°C under humidified atmosphere of 95% air and 5% CO2. Lewis lung carcinoma (LLC) cells were purchased from the Institute of Biochemistry and Cell Biology of Chinese Academy of Sciences and cultured in DMEM (Gibco-BRL) with 10% fetal bovine serum at 37°C in 5% CO2.
AFP, gp96 and conjugation
Mouse AFP was purchased from GenWay Biotech, Inc. Lyophilized material was resuspended in sterile distilled water at 10 mg/mL, aliquoted, and stored at −70°C until use.
Mouse gp96 expressing Escherichia coli colony was provided by Dr. Q Wang from Xi'an Jiaotong University, China. The gp96 expression vector contained an isopropyl β-D-1-thiogalactopyranoside (IPTG) inducible T7 promoter and was in frame with the C-terminal 6-His-tag (Invitrogen Corp.). E. coli cells transformed with the plasmid were grown until the OD reached 0.5, and then IPTG (1 mM final concentration) was added. After induction with IPTG at 37°C for 3 hours, cells were disrupted by sonication in a lysis buffer containing a nonionic detergent and lysozyme. Recombinant gp96 protein was purified using nickel nitriloacetic acid (Ni-NTA)-agarose (Qiagen) column. The identity of the purified protein was verified by western blotting with a polyclonal antibody (Santa Cruz Biotechnology Inc.). Protein concentration was determined using BCATM protein assay kit (Pierce Inc.).
One milligram AFP was coupled to 1 mg gp96 in the presence of 0.2% glutaraldehyde in phosphate buffer saline (PBS) for 2 hours, and then dialyzed against PBS overnight. Aliquots of each conjugate were stored at −70°C until use.
Purification and western blot analysis
To obtain the 1:1 dimer of gp96 and AFP cross-linking protein, the cross-linking protein complexes were sequentially isolated and respectively by gp96 polyclonal antibody-affinity chromatography and AFP polyclonal antibody-affinity chromatography. After the two column elutions of bounded gp96/AFP cross-linking complexes, the condensed eluted samples underwent the preparation fractionation by electrophoresis to separate the 1:1 dimer of gp96/AFP cross-linking protein according to previous methods. 19,20
Briefly, the gp96 or AFP polyclonal antibody coupling was performed using Affi-Gel 15 kit (153-6051; Bio-Rad Laboratories) according to the manufacturer's recommendations. Gp96 or AFP polyclonal antibody solubilized in Hepes buffer (0.1 M) was incubated with the gel (1 mL) under constant gentle agitation for 4 hours at 4°C. Thereafter, 100 μL of ethanolamine-HCl (1 M, pH 8.0) was added and the gel was incubated for an hour to block protein binding activity. The coupled gel was transferred to the column, washed with Hepes buffer, and equilibrated with PBS, pH 7.2. Pooled gp96/AFP complex was dialyzed against 0.15 M Tris-HCl buffer containing 0.5 M NaCl (pH 7.8) at 4°C for 2 hours and centrifuged at 10,000 g for 10 minutes. The supernatant solution was passed through the gp96 polyclonal antibody-affinity chromatography column in 0.15 M Tris-HCl buffer containing 0.5 M NaCl. The antibody-bound fraction was eluted with 0.2 M glycine-HCI buffer containing 0.5 M NaCl (pH 2.5) and adjusted to between pH 7.0 and 7.5 with 40% Tris buffer (pH 11). The eluted solution was dialyzed and centrifuged as mentioned above, passing through the AFP polyclonal antibody-affinity chromatography column in the same buffer. The final eluted solution was dialyzed at PBS (Ph 7.2) at 4°C for 2 hours.
1:1 dimer of cross-linking complex gp96/AFP purity was verified by western blotting with polyclonal antibodies Santa Cruz Biotechnology, Inc. Protein concentration was determined using BCATM protein assay kit (Pierce, Inc.).
After centrifugation, the purified protein complex was denatured with SDS sample buffer (62.5 mM Tris-HCL, pH 6.8, 25 g/L SDS, 50 mL/L β-mercaptoethanol, and 100 mL/L glycerol) at 100°C for 5 minutes. The purified protein complexes were then analyzed by 90 g/L SDS-polyacrylamide gel electrophoresis (SDS-PAGE). Proteins were transferred to nitrocellulose membrane (Bio-Rad) and detected by immunoblotting with gp96 rabbit antimouse polyclonal antibody (1:100) or AFP rabbit antimouse polyclonal antibody (1:100) at 4°C overnight, respectively. After a complete wash in PBS, the membranes were treated with horseradish peroxidase-labeled goat antirabbit antibody (1:100) for 45 minutes at 37°C. After a complete wash in PBS, the membranes were developed in 0.5 g/L freshly prepared 3, 3′-diaminobenzedine solution (DAB; Sigma Co.) for 8 minutes. Purified gp96 and AFP were used as positive controls to determine the specificity of polyclonal antibody. β-actin was used as a negative control and internal reference.
Mice immunized with gp96/AFP complex
Female BALB/C mice were randomly divided into gp96/AFP group, AFP group, gp96 group, PBS control, and empty group. Every group had 10 mice. Before injection, each group was diluted in saline to 100 μg/100 μL. Various groups of mice were subcutaneously (s.c.) injected into the left flank of mice. Priming and boosting were performed with 10 μg gp96/AFP, AFP, or gp96, whereas PBS was used as controls. A 0.3 insulin syringe with a 25-gauge 0.5-inch-long needle was used for the s.c. injections. Mice were boosted s.c. with above proteins twice at 2 weeks intervals after the first priming. Two weeks after last immunization, splenocytes were harvested and diluted to different concentrations.
ELISPOT assay
The ELISPOT was used to measure the frequency of specific T cells producing the cytokine IFN-γ in splenocytes harvested from the immunized mice. BD ELISPOT Plates (BD PharMingen) were coated with 5 μg/mL rat antimouse IFN-γ antibody in 100 μL of PBS. After overnight incubation at 4°C, the wells were washed and blocked with RPMI-1640 culture medium containing 10% fetal bovine serum. 1×106 splenocytes were added to the ELISPOT plate wells along with 5 μg/mL of AFP containing 10% fetal bovine serum and 10 U/mL of human interleukin (IL)-2 (PEPRO Tech ET Ltd.). After culture at 37°C for 24 hours, the plate was washed and then followed by incubation with 2.5 μg/mL biotinylated IFN-γ antibody in 100 μL PBS containing 10% FCS at 4°C overnight. After washing, avidin-HRP in 100 μL PBS was added and incubated for 1 hour at room temperature. After washing five times, spots were developed by adding 100 μL 5-bromo-4-chloro-3-indolyl phosphatase/Nitro Blue Tetrazolium (Boehringer Mannheim). The color spots, representing cytokine-producing cells, were counted using an ELISPOT Reader System.
ELlSA assay
To determine the level of AFP or anti-AFP antibody in mice, we examined the serum of mice tail vein before the immunization or after the last immunization by ELISA. A 96-well microplate was coated with 100 μL of 5 μg/mL AFP or anti-AFP antibody and incubated at 4°C overnight. The wells were then blocked with PBS containing 5% bovine serum albumin. Sera were prepared from the mice on day 14 postimmunization, serially diluted in PBS, added to the ELISA wells, and incubated at room temperature for 2 hours. After washing with PBS-T containing 0.05% Tween-20, the plate was incubated with 1:3000 dilution of a HRP-conjugated goat antimouse IgG antibody (Sigma-Aldrich Corp.) at room temperature for 1 hour. The plate was washed five times, developed with O-phenylenediamine away from light at 37°C for 15 minutes, and stopped with 50 μL of 2 M H2SO4. The ELISA plate was read with a standard ELISA reader at 490 nm. The quantity of AFP or antibody was measured in comparison with standard sample diluents.
Cytotoxic T-lymphocyte assays
BALB/C mice were immunized s.c. as described above. Two weeks after the last boost, 2.5×107 splenocytes were cocultured with 5 μg/mL of AFP containing 10% fetal bovine serum, 10 U/mL of human IL-2 in RPMI 1640 supplemented with 10% FCS at 37°C in 5% CO2. After 5 days of stimulation, the viable splenocytes were recovered and used as effector cells, and the H22 cells or LLC cells were used as target cells. The Non-Radioactive Cytotoxicity Lactate Dehydrogenase (LDH) release assay (Promega) was performed to measure the effector cells against H22 or LLC tumor cells in the ratios of 10:1, 20:1, and 40:1, according to the manufacturer's protocol. Specific lysis was calculated according to the formula: percent specific lysis=(experimental release value − effector spontaneous release value − target spontaneous release value)/(target maximum release value − target spontaneous release value)×100. Results shown are representative of experiments repeated thrice.
CD8+ T-cell depletion cytotoxic T-lymphocyte assays
To test CD8+ T-cell response, a separate set of immunized mice were sacrificed and spleens were collected. CD8
In vivo tumor therapeutic experiments
To test the ability of gp96/AFP vaccination to inhibit the growth of established tumors, BALB/C mice in every group were injected s.c. in the left flank with 1×106 H22 cells per mouse on day 0, and then injected s.c. in the right flank with different vaccination treatments on day 7, day 14, and day 21 as described above. H22 tumor cells were washed after enzymatic digestion and resuspended in 0.2 mL of PBS per animal to be injected s.c. into the left flank, while PBS was used as control. The tumor growth was monitored every day. Tumor size was measured in two dimensions with calipers every 3 days 1 week after tumor inoculation. At each time point, tumor size was determined by measuring the smallest diameter (a) and the biggest diameter (b). Tumor volume was calculated using the formula: V=(a 2 b)/2. Tumor size measurements were performed by the mean value of each group and executed in duplicate to confirm the results. Percentage of tumor-free mice was recorded and the survival of mice was monitored for 10 weeks from the day of tumor challenge.
Immunostaining of AFP in tumor-loaded mice after treatment with gp96/AFP
Fourteen days after the last immunization with gp96/AFP, gp96, and AFP, the mice were sacrificed and the tumors were dissected and fixed in 10% buffered formalin and embedded in paraffin wax. 5-μm-thick serial sections were cut and placed on silane-coated glass slides. All sections were deparaffinized and rehydrated with graded alcohol. Endogenous peroxidase was then blocked with 0.3% H2O2 diluted in methanol for 30 minutes at room temperature. Antigen retrieval was performed by treating the slides in citrate buffer in a microwave for 10 minutes. The slides were incubated in a moist chamber with AFP rabbit polyclonal antibody (1:100) at 4°C overnight. After a complete wash in PBS, the slides were treated with horseradish peroxidase-labeled goat antirabbit antibody (1:100) for 45 minutes at 37°C. After a complete wash in PBS, the slides were developed in 0.05% freshly prepared diaminobenzidine solution (DAB; Sigma Co.) with 0.03% hydrogen peroxide for 8 minutes, and then counterstained with hematoxylin, dehydrated, air dried, and mounted in neutral resins. Normal mouse IgG was used as a substitute for the primary antibody to serve as a negative control.
Statistical analysis
All statistical analyses were done using the SPSS software. Numerical results were expressed as mean±SEM of multiple experiments. ANOVA was used to determine the level of difference between groups. Pairs of groups were compared by unpaired two-tailed Student's t-test. Significance was determined as p-values<0.05.
Results
Identification of purified gp96/AFP complex
To obtain purified gp96/AFP complex, a sequential polyclonal antibody-affinity chromatography and preparation fractionation by electrophoresis was used. By gp96 polyclonal antibody-affinity chromatography and AFP polyclonal antibody-affinity chromatography respectively, the nonconjugated AFP and gp96 were eluted. After the preparation fractionation by electrophoresis, the 1:1 dimer of cross-linking complex gp96/AFP was obtained. The purification of gp96/AFP complex was confirmed by SDS-PAGE and western blot (Fig. 1). The purified gp96/AFP complex was identified by gp96 antibody and AFP antibody via western blot. The results confirmed that gp96/AFP complex was constructed and produced successfully.

Analysis of purified glycoprotein 96 (gp96)/α-fetoprotein (AFP) by western blot. The identity of the purified protein was verified by western blotting with polyclonal antibodies. The relative molecular weights of gp96, AFP, gp96/AFP, and β-actin were ∼96, 70, 166, and 42 kDa, respectively. Lane 1, purified gp96; Lane 2, AFP; Lane 3, purified gp96/AFP complex with gp96 antibody; Lane 4, purified gp96/AFP complex with AFP antibody; Lane 5, β-actin; M, protein molecular weight markers.
Prime-boost vaccines generate AFP-specific T cells in vivo
Since cytotoxic T-lymphocytes (CTLs) have been known to play critical roles in tumor immunity, we examined IFN-γ producing specific T-cell frequency after gp96/AFP vaccination by ELISPOT assays. Two weeks after the last immunization, splenocytes were harvested and stimulated with AFP. As shown in Figure 2, immunization of BALB/C mice with gp96/AFP vaccine elicited much more strong T-cell responses than those with AFP or gp96 immunization alone (p<0.01). There was no significant difference between the groups vaccinated with AFP, gp96, and PBS alone (p>0.05). Our data suggested that gp96/AFP vaccine generated significantly higher AFP-specific T-cell response compared with other vaccinated groups, while vaccination with AFP or gp96 alone failed to elicit a measurable T-cell response.
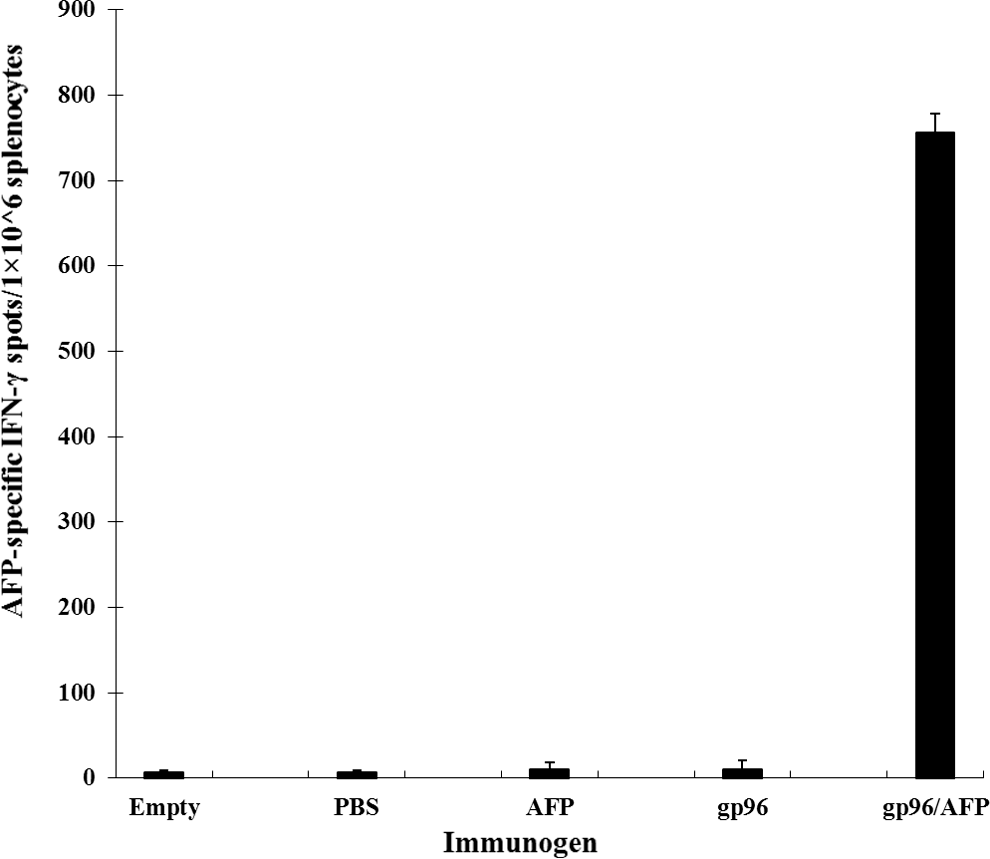
Quantity of AFP-specific T cell secreting IFN-γ in immunized BALB/C mice. The immunized mice spleen cells were tested by ELISPOT and AFP-specific IFN-γ spots were counted. The spot numbers were the mean of triplicates±SE in each vaccinated group at 1×106 cell number. Results shown here are AFP-specific spot-forming cells from five groups. Each group was tested in triplicate. PBS, phosphate buffer saline.
In vivo AFP-specific antibodies detected after Gp96/AFP immunization
The quantity of AFP or anti-AFP antibody in the sera of the vaccinated mice was determined by ELISA. 7 days after the injection with H22 cells, all the tumor-loaded mice produced higher level of AFP compared with the empty control group (p<0.01) (Supplementary Fig. S1; Supplementary Data are available online at
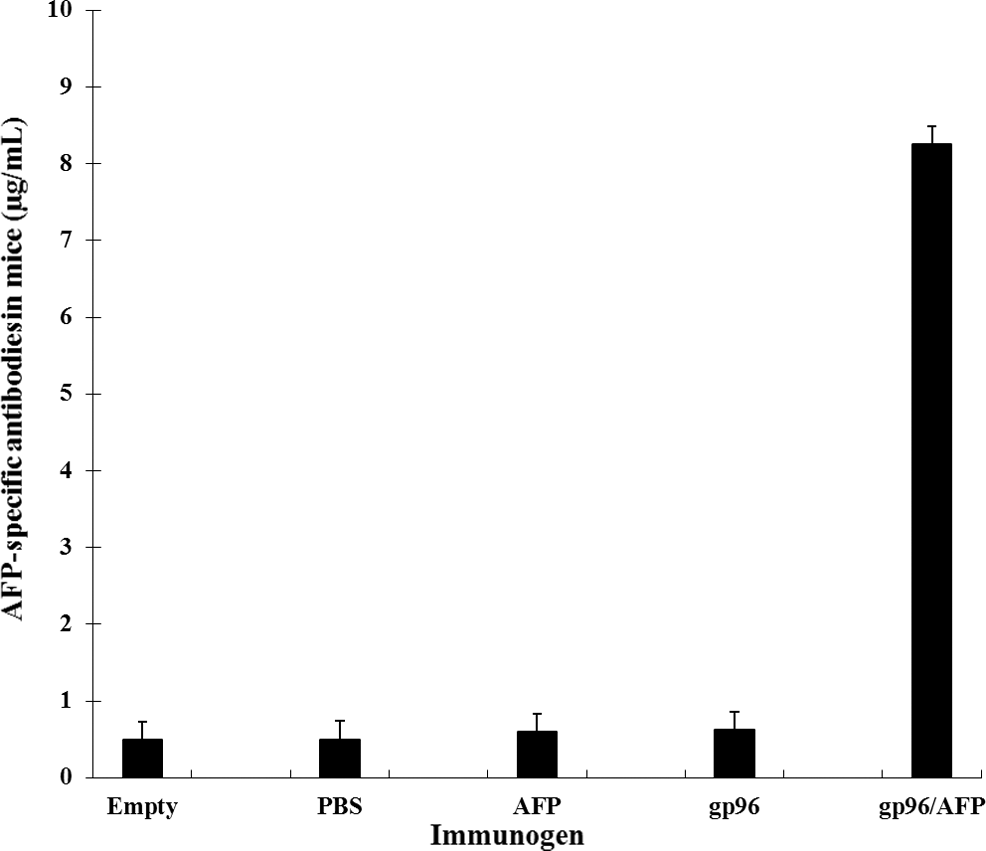
AFP-specific antibody responses in immunized BALB/C mice. BALB/C mice were immunized with gp96, AFP, and gp96/AFP. PBS was used as negative control. Serum samples were obtained from immunized mice 14 days after last vaccination. The presence of the AFP-specific antibody was detected by ELISA using serial dilution of sera. The results from the 1:20 dilution are presented showing the mean absorbance (A490 nm)±SE.
Gp96/AFP elicited specific cytotoxic effect against H22 tumor cells
It has been previously demonstrated that AFP/HSP70 recombinant vaccine is able to induce the regression of AFP-expressing tumors in mice through either prophylactic or therapeutic treatment. 10,21,22 To examine whether gp96/AFP vaccination is able to prime stronger AFP-specific cytotoxic T cells than gp96 or AFP immunization, we performed in vitro cytotoxicity assays of lymphocytes isolated from mice immunized with gp96/AFP, gp96, AFP, and PBS respectively. Splenocytes were harvested 2 weeks after immunization and restimulated in vitro with AFP. Viable effector cells were assessed for cytotoxic activity against H22 cells or LLC cells (AFP-negative control). As shown in Figure 4, effector cells from mice immunized with gp96/AFP showed much stronger cytolytic effects on H22 target cells than those from mice vaccinated with gp96, AFP, or PBS (p<0.01). Additionally, vaccination with either AFP or gp96 alone failed to induce measurable specific T cell-mediated cytotoxicity. There was no significant difference between the groups vaccinated with AFP, gp96, or PBS alone (p>0.05) (Fig. 4A). In addition, this cytolytic activity was specific for H22 cells, not for LLC cells (p<0.01) (Fig. 4B). Our results indicated that the cross-linkage of AFP to gp96 was required for enhancement of specific T cell activity, since the administration of AFP or gp96 alone failed to elicit a distinct cytolytic activity.
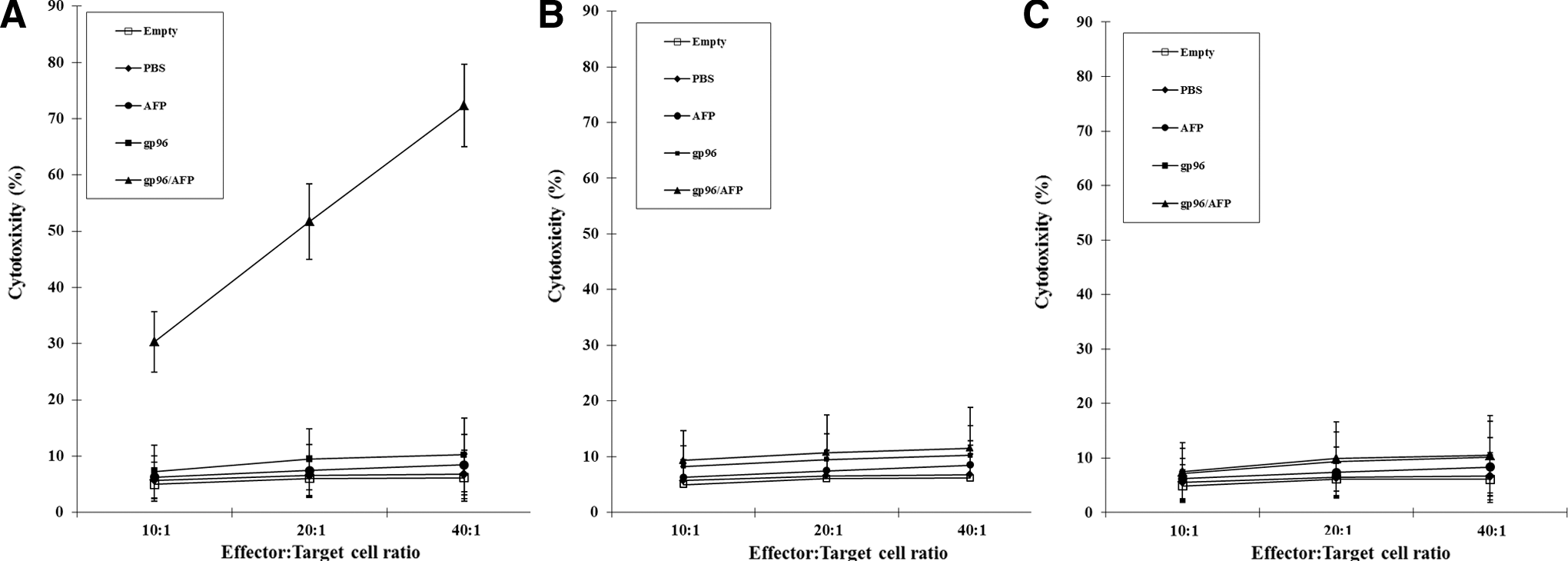
Gp96/AFP complex primed the strongest AFP-specific CD8+ T-cell responses. Mice were immunized with gp96/AFP, gp96, AFP, and PBS respectively. Mice were subcutaneously boosted with the above proteins twice at 2-week intervals after the first priming. Two weeks after the last boost, pooled splenocyte cultures were prepared and restimulated in vitro with AFP to generate effector cells separately. AFP-specific cytolytic activity was assayed against as target cells at 10:1, 20:1, and 40:1 effector/target (E/T) ratios for H22 cells
Specific CD8+ T cell cytotoxic effect against H22 tumor cells elicited by gp96/AFP
To elucidate whether CD8+ T cells participate in the cytotoxic against H22 tumor cells, CD8+ T-cell depletion assay was determined. After CD8+ T-cell depletion, the remaining splenocyte effector cells from mice vaccinated with gp96/AFP showed much lower cytolytic effects on H22 target cells. There was no significant difference from mice treated by gp96, AFP, or PBS alone (p>0.05) (Fig. 4C). Our results indicated that CD8+ T cells participated in the specific cytolytic activity against H22 tumor cells via gp96/AFP immunization.
Gp96/AFP vaccination induce therapeutic immunity against H22 tumors
In view of the AFP-specific CTL response elicited by gp96/AFP complex, we examined the therapeutic effect of different protein vaccines to induce regression of pre-existing AFP-expressing H22 tumor in vivo. In these studies, female BALB/C mice were injected with 1×106 H22 cells s.c. in the left flank, and then vaccinated s.c. in the right flank with gp96/AFP, AFP, and gp96 on days 7, 14, and 21. PBS (100 μL/mouse) treatment was used as negative control. The growth of tumor was monitored for about 4 weeks since day 4 post tumor inoculation. As shown in Figure 5A, 100% of the mice immunized with gp96/AFP had tumor-free survival for more than 70 days. In contrast, some of mice receiving gp96 or AFP developed tumor growth within 7 days after tumor challenge. All of the mice receiving PBS developed a tumor growth on day 7 after the tumor challenge. Only 20% or 30% of the mice immunized with AFP or gp96 remained tumor-free 70 days after H22 challenge. In addition, tumor-free percentage of AFP immunized mice (20%) was significantly lower than that of gp96/AFP immunized mice, indicating that cross-linkage between AFP and gp96 is essential for effective therapy. For comparison, the mean tumor volumes from various groups of mice were shown in Figure 5B; mice immunized with gp96 or AFP alone demonstrated higher average tumor volumes than mice immunized with gp96/AFP (p<0.05). There was a statistically significant difference in the therapeutic effect of H22 tumor between gp96/AFP and AFP vaccinated group (p<0.01). However, there was no statistical difference in the tumor mass among the groups immunized with AFP, gp96, and the group administrated with PBS. Different vaccinations greatly influenced the survival of mice. About 70%–80% mice immunized with AFP or gp96 died before day 49 and all the mice immunized with gp96/AFP still survived at day 70, whereas all the mice treated with PBS died before day 42. These data suggested that gp96/AFP immunization could significantly reduce tumor size and prolong the survival time of the tumor-bearing mice than AFP immunization alone, indicating that the vaccination of gp96/AFP induced a desirable therapeutic effect against H22 tumor cells loaded in vivo. In summary, these results showed that vaccination with gp96/AFP could eradicate previously inoculated AFP-expressing tumors in mice and induced a robust antitumor activity in vivo.
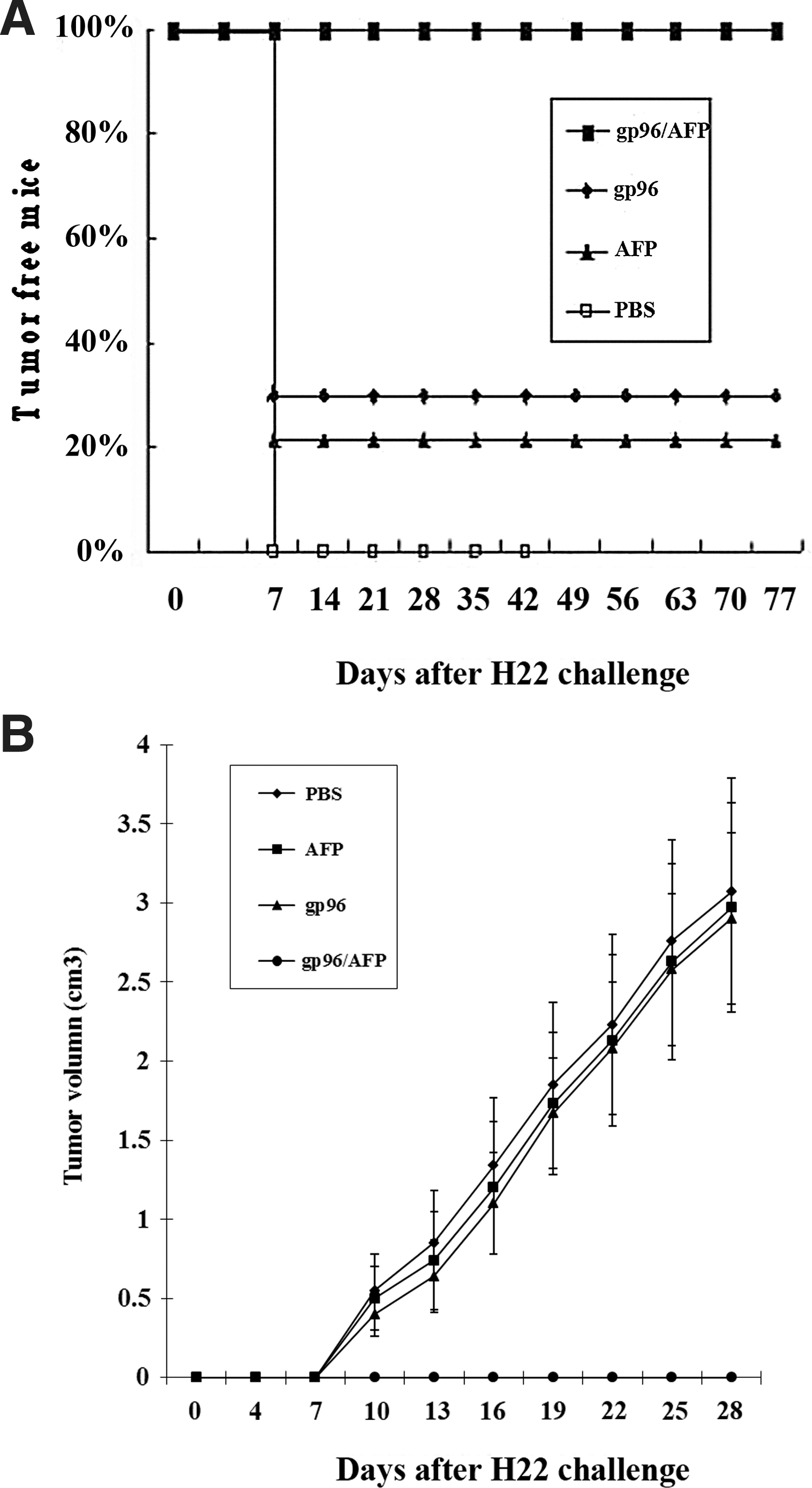
Therapeutic immunization of mice with gp96/AFP vaccination.
Morphological changes of mice loaded tumor treated with gp96/AFP
The immunostaining showed that AFP was mainly localized in the cytoplasm of HCC cells. The tumor loaded mice that were treated with gp96/AFP displayed the shrunk restricted cancer nests, most of which occurred to be necrotic and replaced by fibrous and connective tissues. AFP staining was also confined in part of the cell cytoplasm. The AFP and gp96 vaccinated groups showed a large area of living cancer nests. Most cell cytoplasm were stained with AFP, similar to the empty control group (Supplementary Fig. S2).
Discussion
AFP always accompanies the growth of liver cells, and it has been confirmed that AFP is probably related to the proliferation of tumor or fetal cells. 4,23 The mechanism for growth-promoting activity of AFP is still unclear. Escaping from the surveillance of immune system is the primary cause for malignant growth of HCC cells. 24,25 Several investigations have showed that AFP could be synergistic with other growth factors individually to promote the growth of many tumor cells. 26,27 AFP receptors have been found anchoring on the membrane of various tumor cells. 21,28 The receptor may mediate intercellular signal transduction, which influences the expression of genes related to proliferation. 28 AFP could stimulate the expression of some oncogenes that control cell cycle, and then enhance the proliferation of human HCC. When BEL-7402 cell line was treated with AFP, oncogene proteins, such as c-fos, c-jun, c-ras, and mutant p53 and p21ras rapidly increased, which exerted important functions in modulating the growth and differentiation of the tumor cells. 28,29
Recent studies on the immunodominant epitopes of AFP have provided a solution to the obstacle of HCC immunotherapy. AFP is produced at low serum levels after birth throughout life. The majority of human HCC overexpress the oncofetal antigen AFP, Mr 64,000–72,000 glycoprotein. Despite being exposed to high plasma levels of this oncofetal protein during embryonic development, body still displays a low immunity to it. 28 –30 Butterfield has found that four peptides of human AFP processed and presented in the context of HLA-A0201, could be recognized by the human T cell repertoire, and could be used to generate AFP-specific CTL in human T-cell cultures. 6 It was also found that murine immune system could generate T-cell responses to this oncofetal antigen. Therefore, AFP may be an ideal target for immunotherapy. But AFP immunization alone still results in lower levels of specific response and poorly reproducible protective immunity. 4,5
How to enhance host's active immunity to AFP may be an interesting strategy for HCC therapy. In the present study, we tested a novel strategy to induce antitumor immunity by a reconstructed vaccine conjugation AFP to gp96 in mice via glutaraldehyde cross-linking. We found that compared with immunized with AFP or gp96 alone, the gp96/AFP vaccine could elicit strong AFP-specific T-cell responses and produce a distinct protective effect on AFP-producing tumor. CD8+ T cells depletion assay further confirmed that vaccination with gp96/AFP could elicit relatively strong specific CD8+ T-cell response and cytotoxic antitumor effect, whereas vaccination with either AFP or gp96 alone failed to induce measurable CD8+ T cell-mediated cytotoxicity.
It is of interest to note that the gp96/AFP cross-linking protein vaccine provoked not only the considerable stability of specific cellular immunoprotection, but also a detectable level of anti-AFP antibody, whereas vaccination with AFP or gp96 alone failed to elicit a measurable humoral response. The above results indicated that AFP, by itself, is not immunogenic or weakly immunogenic and conjugating to gp96 rendered AFP highly immunogenic.
Protein-based vaccine has become an attractive approach to generate antigen-specific immunotherapy because of its simplicity, safety, efficacy, and capacity for repeated administration. Protein vaccines generate effective CTLs and antibody responses by involvement of APCs that stimulate CD4+ and CD8+ T cells. Protein vaccines could be easily prepared on a large scale with high purity and stability and could also be repeatedly given to the same patient safely and effectively. 1,5 The features make protein vaccine a potentially attractive approach for cancer therapeutic vaccine development.
However, there is still a need to increase the potency of protein vaccines. Since tumor antigens are weakly immunogenic, researchers are currently emphasizing the use of versatile immunostimulatory molecules in the newer therapeutic cancer vaccines. Various HSP-based vaccine strategies have been described to enhance fusion vaccine potency. In early studies from our group, we constructed an eukaryotic expression vector containing the molecular chaperon-HSP70 and AFP fragments, and then primed mice with the genetic vaccine, eliciting a robust strong protective immunity. 10,21 Another research group also confirmed that the linkage of HSP70 to target antigen dramatically enhanced the potency of AFP fusion protein vaccines. 22 Therefore, an innovative approach that combined HSPs with tumor antigen would probably generate more potent antitumor effects. Gp96 is one member of the HSP family that has been shown to act as a potent adjuvant to enhance antigen-specific tumor immunity and innate immunity. 31 –33 The use of gp96 conjugated antigen vaccines represents a promising approach for enhancing antigen-specific T cell-mediated immune responses and humoral responses for cancer vaccine development. 13,32,33 Currently clinical trials on gp96 vaccine have confirmed the antitumor effect for certain cancers. 34 –36
A few researches reported that HSPs could enhance the risk of autoimmunity due to cross-reactive T-lymphocytes primed by the HSP vaccines. 35,37,38 However, the experiments that we have conducted showed that vaccination with gp96/AFP protein vaccine did not result in pathologic changes in the major organs of mice (data not shown). In this study, we described a novel AFP protein-based immunization strategy by cross-linking gp96 and AFP protein. Our data demonstrated that gp96/AFP vaccination could elicit strong AFP-specific CD8+ T-cell immune responses and induce established H22 tumor regression. Consistent with the in vivo results, the gp96/AFP protein immunization induced specific CTLs that recognized H22 tumor cells in vitro, but not LLC cells. Our data also demonstrated that gp96/AFP protein vaccine generated the strongest therapeutic effects compared with AFP or gp96 immunization alone. Therefore, a novel strategy that combined linkage of gp96 with antigen in the form of cross-linking protein cooperatively could enhance the potency of antigen-expressing protein vaccines.
Cross-linking of gp96 and AFP with lower concentration of glutaraldehyde not only established the gp96/AFP complex but also preserved the activity of gp96 immunoadjuvant and immunogenicity of AFP antigen, which was confirmed by the strong ex vivo and in vivo antitumor effect. We attributed the successful AFP-specific T-cell responses in mice to the gp96 through mediating APCs to efficiently take up and process AFP. Researches have shown that gp96 itself has no antigenicity and its immunogenicity has been attributed to the peptides chaperoned by gp96. 18,32,33 It has been verified that gp96 was a better molecule chaperon and adjuvant that could process and present weak tumor antigen to MHC-I of host APCs via the “cross-priming” mechanism, 32 –34 eliciting specific T-cell response and CTL reaction. 13,36,39 Several studies have shown that gp96-associated peptides could anchor antigen on the cell membrane and directly present it to natural killer cells or γδ+ T cells as superantigen without being dependent on the stimulation of MHC-I molecules. 33,34,40,41 In the present study, tumor rejection assay demonstrated that cross-linking protein vaccine gp96/AFP elicited strong specific antitumor immunity against AFP-producing H22 cells than AFP vaccination alone. Results indicated that AFP immunogenicity could be greatly improved by gp96 and vaccination with gp96 could increase T-cell proliferation responses to AFP.
Conclusions
In summary, we developed a strategy to construct an innovative protein vaccine gp96/AFP. Our results indicate that conjugating to gp96 renders AFP highly immunogenic. Gp96/AFP protein vaccine can generate an impressive antitumor effect against AFP-expressing murine tumors through enhancement of AFP-specific CD8+ T cell-mediated immune responses. Therefore, cross-linking oncofetal antigen AFP and gp96 is a promising approach for HCC immunotherapy, which may be potentially applied to other cancer systems with the known tumor-related antigens.
Footnotes
Acknowledgments
This work is supported by the Scientific Research Program Funded by Shaanxi Provincial Education Department (No. 2007JK233, 2010JK484), the Key Project of Ministry of Education of China (No. 205002) and the National Natural Science Foundation of China (No. 81172135).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.