
Abstract
Purpose:
The therapeutic efficacy of a synthetic parasite-derived peptide GK1, an immune response booster, was evaluated in a mouse melanoma model. This melanoma model correlates with human stage IIb melanoma, which is treated with wide surgical excision; a parallel study employing a surgical treatment was carried out as an instructive goal.
Experimental Design:
C57BL/6 mice were injected subcutaneously in the flank with 2×105 B16-F10 murine melanoma cells. When the tumors reached 20 mm3, mice were separated into two different groups; the GK1 group, treated weekly with peritumoral injections of GK1 (10 μg/100 μL of sterile saline solution) and the control group, treated weekly with an antiseptic peritumoral injection of 100 μL of sterile saline solution without further intervention. All mice were monitored daily for clinical appearance, tumor size, and survival. Surgical treatment was performed in parallel when the tumor size was 20 mm3 (group A), 500 mm3 (group B), and >500 mm3 (group C).
Results:
The GK1 peptide effectively increased the mean survival time by 9.05 days, corresponding to an increase of 42.58%, and significantly delayed tumor growth from day 3 to 12 of treatment. In addition, tumor necrosis was significantly increased (p<0.05) in the treated mice. The overall survival rates obtained with surgical treatment at 6 months were 83.33% for group A, 40% for group B, and 0% for group C, with significant differences (p<0.05) among the groups.
Conclusions:
The GK1 peptide demonstrated therapeutic properties in a mouse melanoma model, as treatment resulted in a significant increase in the mean survival time of the treated animals (42.58%). The potential for GK1 to be used as a primary or adjuvant component of chemotherapeutic cocktails for the treatment of experimental and human cancers remains to be determined, and surgical removal remains a challenge for any new experimental treatment of melanoma in mouse models.
Introduction
Melanoma is the most malignant form of skin cancer, and the frequency of melanoma cases is rising rapidly, especially in the Caucasian population. 1,2 It is estimated that there will be 76,250 new cases and 9,180 deaths in the United States in 2012. 3 Metastases are the leading cause of death in advanced melanoma patients. Despite the numerous innovations that have been made for the treatment of this disease, the 5-year survival rate for patients with distant metastases is <10%. 4 Moreover, the survival times for patients with lymph node metastases average 12 to 15 months, whereas those for patients with liver and bone metastases average 3 to 4 months. 5
Surgical resection, chemotherapy, and immunotherapy are the conventional therapeutic strategies for patients with malignant melanoma. 6 Local melanoma can be cured by wide surgical excision in its early stages, 7 but metastatic melanomas are usually incurable. 8 Dacarbazine and cytokine adjuvant therapy (e.g., high-dose interferon alfa-2b [IFN-α2b] and interleukin-2 [IL-2]) are commonly used as palliative systemic chemotherapy in patients with advanced melanoma. 2,9,10
Despite extensive research, there has been no major improvement in the treatment of advanced melanoma for many years. 11 Cancer vaccines exhibit high specificity and low toxicity, but their therapeutic efficacy has been very low, with a reported overall objective response rate of only 3.3%. 12 However, cancer immunotherapy has undergone considerable progress since the first promising results with adjuvant therapy using IFN-α2b and IL-2. 13 –16
None of the clinical trials evaluating immunostimulatory strategies, such as the use of Bacillus Calmette-Guérin, have demonstrated any systemic benefit for melanoma patients. 17,18 Several pathogen-associated molecular patterns, such as lipopolysaccharides (LPS), peptidoglycan, outer membrane protein A, and unmethylated cytosine-phosphorothionate guanine-rich oligodeoxynucleotides, have been shown to provide adjuvant effects when used in combination with tumor antigen-specific dendritic cell (DC)-based immunotherapy in murine melanoma models. 19 –22 Recently, DC-based immunotherapy was clinically approved, and it has been used for the treatment of melanoma. 23
Previous studies have shown that GK1, an 18-amino-acid peptide derived from Taeniacrassiceps cysticerci, 24 induces a high level of protection against cysticercosis 25 without the need for an adjuvant. In addition, the subcutaneous co-administration of GK1 with an influenza vaccine (Fluzone™) increased the levels of anti-influenza virus antibodies in aged mice before and after infection. 26 The mechanisms of action of this parasite-derived peptide are related to the effective enhancement of antigen-specific CD4 T-cell priming in vivo and in vitro and to the direct up-regulation of costimulatory molecules on DCs. Moreover, GK1, used in combination with LPS, has been shown to stimulate the production of IFN-γ and tumor necrosis factor(TNF)-α and also to boost the immune response by inducing the secretion of CCL2, a proinflammatory chemokine that attracts monocytes, natural killer (NK) cells, immature DCs, and activated T cells to the lymph nodes draining the immunization site. 27 In this study, we determined whether peritumoral injections of GK1 would induce antitumor activity.
Materials and Methods
Animal studies
All of the experiments using mice complied with standards equivalent to the United Kingdom Coordinating Committee on Cancer Research (UKCCCR) guidelines for the welfare of animals with experimental neoplasia; the number of mice was restricted to a minimum that was consistent with the purpose of each assay, 28 and the experiments were performed once. In addition, all of the animals were treated in accordance with the recommendations of the Ethics Committee of the School of Medicine, Universidad Nacional Autónoma de México (UNAM), (project 040/2009 approved June 2, 2009), the Animal Care and Use Committee, and the Mexican Official Regulations (NOM 062-ZOO-1999). Male C57BL/6 mice, between the ages of 6 and 7 weeks old, were purchased from Harlan (Mexico City, Mexico). The animals were acclimated to the animal facility of the Department of Cell and Tissue Biology (School of Medicine, UNAM) for a minimum of 1 week. The mice were separately housed in ventilated cages under a controlled light cycle (12 h light/12 h dark) at standard room temperature (22°C–24°C), and they were allowed access to a conventional diet and tap water ad libitum.
Culture medium and tumor cell lines
B16-F10 melanoma cells (American Type Culture Collection, Manassas, VA) were grown in RPMI medium (GIBCO, Grand Island, NY) supplemented with 10% heat-inactivated fetal bovine serum (PAA, Pasching, Austria) and 1% penicillin and streptomycin (GIBCO, Grand Island, NY) at 37°C in a humid 5% CO2 atmosphere. The cells were first harvested from the culture by trypsinization (GIBCO, Grand Island, NY) and were then washed thrice with cold serum-free medium before a subcutaneous injection. 29
Experimental melanoma development
Thirty-two mice were injected subcutaneously in the flank with 2×105 B16-F10 murine melanoma cells. The tumor size was assessed by measuring the length of the short and long tumor diameter with calipers, and the size was expressed as the tumor volume using the following formula: V=(A 2 ×B)/2. 21 Only mice that developed a tumor of 20 mm3, usually 2 weeks after injections with tumor cells, were included in the GK1 (an 18-amino-acid synthetic peptide [GYYYPSDPNTFYAPPYSA]) (AnaSpec, Inc., Fremont, CA) therapy and control groups. The day that the tumor reached 20 mm3 in size was defined as day 0 of the study.
In vivo effect of GK1 on experimental melanoma
Tumor-bearing mice were separated into two different groups: GK1-treated group (16 mice) was injected weekly with an antiseptic peritumoral injection of GK1 (10 μg/100 μL of sterile saline solution), 27 and control group (12 mice) was treated weekly with an antiseptic peritumoral injection of 100 μL of sterile saline solution without further intervention. The mice were sacrificed when they became moribund or lethargic or when they failed to respond to gentle stimuli, according to the UKCCCR guidelines. 28
The mice were examined daily for their clinical appearance, and tumor size was assessed by measuring the tumor diameters with calipers, with the size expressed as tumor volume. Daily tumor sizes from all of the mice in each group were used to calculate growth differences between groups. In addition, the sacrifice day was recorded, and the data were used for the statistical analysis. If a mouse died of a cause unrelated to the neoplasm, the data for that mouse were censored in the analysis.
Assessment of necrotic areas
GK1-treated and control tumor-bearing mice were sacrificed at weeks 2, 3, and 5, at one mouse for each group per week (n=6) and were randomly selected for necrosis analysis. Tissue samples were excised and fixed in 10% neutral-buffered formaldehyde solution, and they were processed to obtain 4-μm-thick paraffin-embedded sections; the sections were stained with hematoxylin and eosin (H&E) and Fontana–Masson (MERCK, Darmstadt, Germany) to identify pigmented cells. After appropriate staining, images from the tissue sections were acquired at 200× (total magnification) with a digital camera to create an image collection in JPEG format (final resolution 3.638 pixels/m), and image analysis was conducted using open-source ImageJ software (
Lung histopathological analysis
GK1-treated and control tumor-bearing mice sacrificed at weeks 2, 3, and 5 (n=6) were randomly selected for lung histopathological analysis. After sacrifice, lungs were excised and fixed in 10% neutral-buffered formaldehyde solution; then, the lungs were examined macroscopically for pulmonary spontaneous metastases identified as a tattoo at the lungs' surface. All lung samples were processed to obtain 4-μm-thick paraffin-embedded sections, which were stained with H&E and Fontana–Masson. The slides were analyzed with a Nikon Eclipse 80i microscope that was equipped with a digital camera.
Surgical approach for the treatment of experimental melanoma
In a parallel study, surgical treatment was performed according to tumor size as follows: group A, 20 mm3 (n=6); group B, 500 mm3 (n=5); and group C, >500 mm3 (ranging from 950–10,512 mm3) (n=7). These tumor sizes corresponded to 2, 3, and 4–6 weeks after an injection of the tumor cells, respectively. Mice were anesthetized with intramuscular injections of 100/5 mg/kg ketamine/xylazine, 31 and a wide excision with 0.5-cm margins was performed under sterile conditions to resect the tumor. Then, the skin was closed with a simple interrupted suture technique using 4–0 nylon, except in the group with tumors larger than 500 mm3, as it was impossible to close the gaps in the skin of these animals. After surgical treatment, the mice were examined daily for clinical appearance and tumor recurrence. No signs of infection at the wound sites were observed. In addition, we recorded the day of death for each mouse. The study ended 6 months after surgery, although the surviving mice were not sacrificed due to their good clinical conditions. Of these mice, 4 of 6 mice in group A and 2 of 5 mice in group B survived for 1 year, at which time they were humanely sacrificed.
Statistical analysis
For each studied variable, the means and SEs were calculated. Survival curves were computed using the Kaplan–Meier method, and survival was validated with the log-rank test. Daily tumor growth differences between groups were evaluated using Mann-Whitney U-test and student's t-test. For tumoral necrosis, data were normalized using the arccos of the proportion of necrotic areas; student's t-test was used to identify significant differences between groups. All of the calculations were performed using SPSS for Windows, version 19 (Statistical Package for Social Sciences, Chicago, IL). Differences were considered statistically significant for p values <0.05.
Results
Experimental melanoma development
Thirty-two male C57BL/6 mice were injected subcutaneously in the flank with 2×105 B16-F10 murine melanoma cells, and only mice that developed tumors of 20 mm3 were included in the study. The presence of melanoma, according to the 20-mm3criterion, was diagnosed in 87.5% of the mice (28 of 32 animals). Tumor growth 2 weeks after the injections of the tumor cells was observed in 22 mice (78.5%). However, it took 3 weeks for the tumors to reach this size in 3 mice (10.7%), 4 weeks in 1 mouse (3.5%), and 8 weeks in 2 mice (7.4%). Interestingly, no tumors were observed in 4 mice up to 1 year after the injections (12.5%); instead, melanotic patches near the sites of the injections were observed, but they eventually disappeared.
In vivo effect of GK1 in experimental melanoma
The tumor size was assessed by measuring the tumor diameters, expressed as tumor volume, and in which moribund or lethargic mice were sacrificed and their data were recorded. The GK1-treated mice demonstrated a significantly longer survival time than the control mice, as the mean survival times were 30.3 days (CI: 25.5–35.2) for the GK1-treated group and 21.25 days (CI:17.38–25.11) for the control group (p<0.05) (Fig. 1A). According to this result, GK1 treatment increased the mean survival time by 9.05 days, corresponding to a 42.58% increase.

Therapeutic anti-tumor activity of the synthetic parasite-derived peptide GK1 against established melanoma. B16/F10 murine melanoma-bearing C57BL/6 mice were peritumorally injected weekly with GK1 (10 μg/100 μL) after their tumors reached 20 mm3 in size (day 0). The control mice were injected weekly with saline solution. Kaplan–Meier survival curves are shown in
From day 3 to 12 of treatment, the tumors in the mice that received GK1 were significantly smaller than those in the control mice (p<0.05) (Fig. 1B). These results suggest that GK1 significantly delayed tumor growth, a tendency also observed before day 3 and after day 12 but that was not statistically significant. Thus, GK1-treated mice lived longer and had smaller tumoral masses compared with the controls.
Assessment of necrotic areas
The experimental melanomas induced in the present work demonstrated highly pigmented, poorly pigmented, and non-pigmented areas in both GK1-treated and control mice. The presence of these areas was confirmed with H&E and Fontana–Masson staining. A quantitative approach revealed that the nonpigmented areas had more mitotic figures than the pigmented areas (p<0.05) (data not shown).
GK1-treated and control tumor-bearing mice were randomly selected for necrosis analysis, and after appropriate staining of tissue sections, an analysis was performed using open-source ImageJ software to measure the threshold that represented the necrotic area, expressed in square micrometers. Overall necrosis, which was recognized as eosinophilic areas inside of the tumors, was quantified as the sum of the necrotic areas from 3 GK1-treated mice and 3 control mice euthanized at 2, 3, and 5 weeks. The results demonstrated that the GK1-treated mice had more necrotic areas than the control mice (p<0.05) (Fig. 2). At the interface of the necrotic areas, numerous neutrophils were observed near or in apposition to distorted or apoptotic tumoral cells, mainly in GK1-treated mice after 2 weeks of treatment (Fig. 3).

Tumor necrosis. GK1-treated and control tumor-bearing mice were randomly selected for tumor necrosis analysis, image analysis was conducted using open-source ImageJ software, and the proportions of necrotic areas were determined for both groups. The tumors in the GK1-treated mice demonstrated more necrotic areas than those in the control mice (p<0.05). Vertical bars denote 95% confidence intervals.
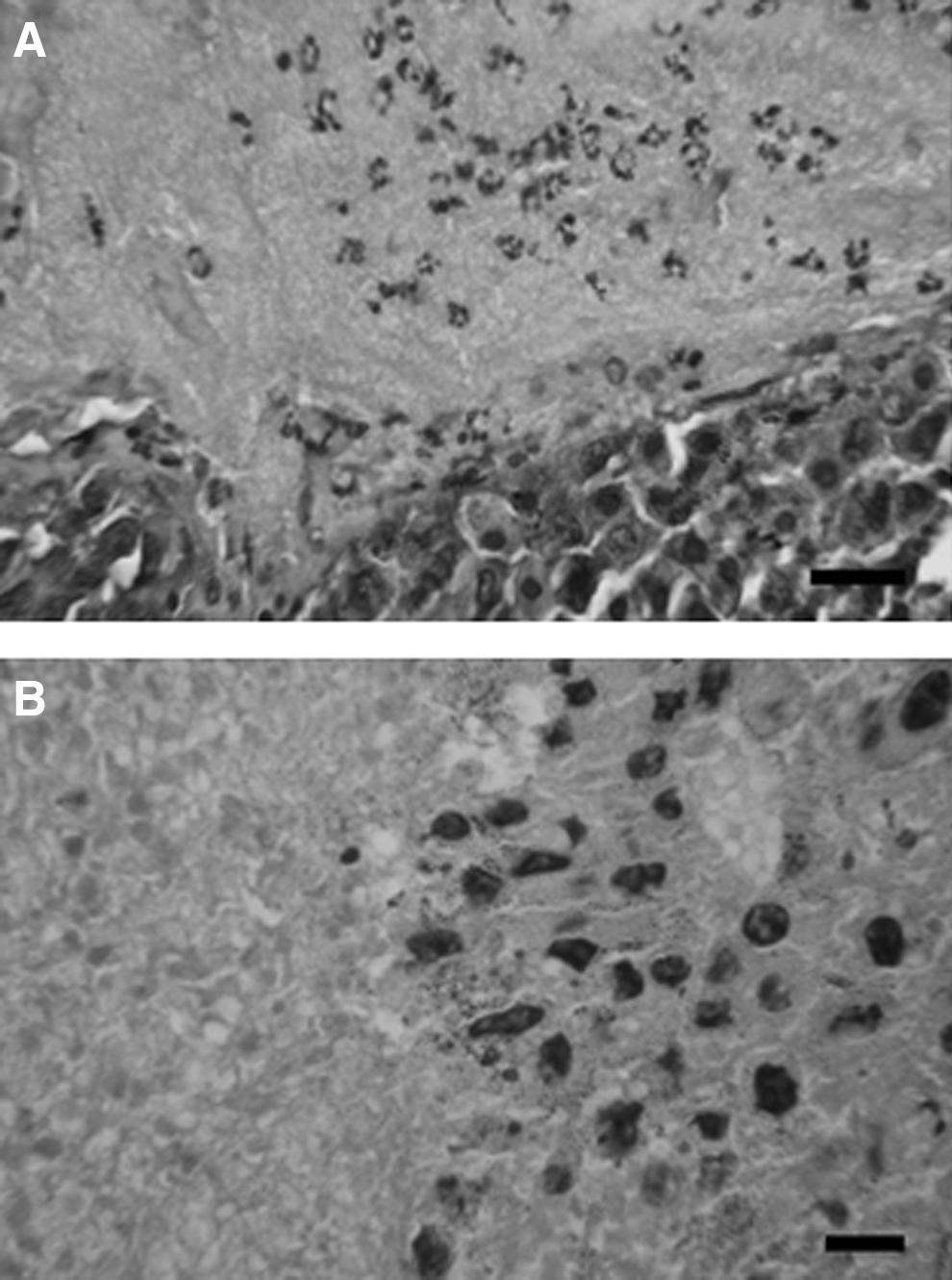
Experimental melanoma histopathology. GK1-treated mouse
Lung histopathological analysis. Tissue sections of lung samples from GK1-treated and control tumor-bearing mice sacrificed at day 15 and 16 showed neutrophilia inside the pulmonary blood vessels, slightly predominant in GK1-treated mice. At this time, no pulmonary metastases were observed macroscopically and microscopically. Control mice sacrificed at day 35 exhibited metastases in the lungs, and neutrophilia was not observed in the samples. Interestingly, neutrophilia was consistently observed in GK1-treated mice sacrificed at day 37 with non-macroscopic or microscopic metastasis. Metastases were frequently located nearby or surrounding bronchioles. Neutrophilia was never associated to any evident inflammatory lung disease or infectious causes (Fig. 4).
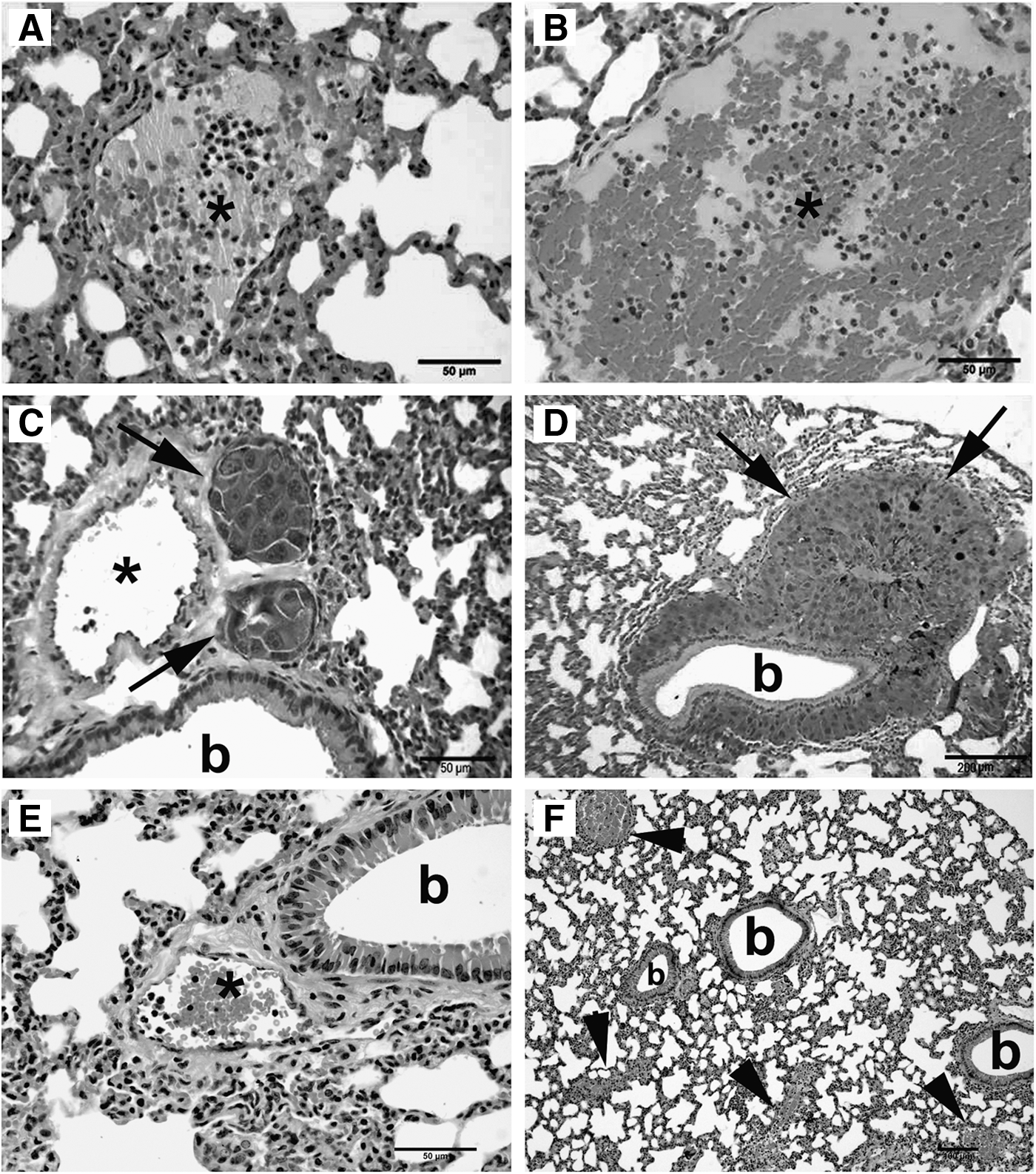
Pulmonary neutrophilia and lung metastases from melanoma. Pulmonary vessels (asterisks) of control
Surgical approach
Surgical treatment was performed according to tumor size; a wide excision with 0.5-cm margins was performed to resect the tumors. In addition, we recorded the day of death for each mouse. The mouse model of melanoma used in the present study was correlated with human Stage IIb melanoma (T4a, N0, M0, T4a>4.0 mm: w/o ulceration), according to the tumor-node-metastasis (TNM) classification system of the American Joint Committee on Cancer. 32 This type of cancer is treated by a primary wide excision with 2.0-cm margins, followed by adjuvant clinical trials or observations. 33 Surgical excision of the experimental melanomas was performed in groups according to tumor size as follows: group A, 20 mm3; group B, 500 mm3; and group C, 950–10,500 mm3. The procedural success rate was 100% for groups A and B and 48.5% for group C. Tumor re-growth was observed in 16.6% of the mice in group A (1/6), in 40% of the mice in group B (2/5), and in 75% of the mice in group C (3/4). One mouse in each group died of causes unrelated to surgery or tumor recurrence. In group A, one mouse died at 7 months, but necropsy did not show an obvious cause of death, and no microscopic metastases were observed; in group B, one mouse died due to stress during manipulation at 2 months; and in group C, one mouse died 2 days after surgery, but no infection was observed in the surgical wound. The overall survival rates at 6 months were 83.33% for group A, 40% for group B, and 0% for group C, with significant differences (p<0.05) among the groups (Fig. 5). These results indicate that early surgical tumor resection considerably improved the survival rate and led to fewer recurrences.

Survival at 6 months after surgery. B16/F10 murine melanomas were surgically removed from C57BL/6 mice, and survival was determined using a Kaplan–Meier survival plot. Surgery was performed when the tumor size was 20 mm3for group A (●), 500 mm3 for group B (♦), and 950–10,500 mm3 for group C (■). The day of surgery represented day 0. The survival rates were 100% for groups A and B and 48.5% for group C at 1 day after surgery. At 6 months after surgery, the survival rates were 83.33% for group A, 40% for group B, and 0% for group C (p<0.05).
Discussion
Peritumoral injections of the synthetic parasite-derived peptide GK1 increased mean survival time by 9.05 days compared with the control treatment, which corresponded to a 42.58% increase in survival time. Furthermore, the tumors in mice that received GK1 were significantly smaller from day 3 to day 12 of treatment than the tumors in the control mice.
It had been reported that GK1 alone was shown to partially enhance CD86 and major histocompatibility complex (MHC) class II expression by DCs and also to enhance antigen-specific T-cell activation both in vitro and in vivo. In addition, GK1 is known to stimulate the production of the proinflammatory cytokines IFN-γ and TNF-α and the inflammatory chemokine CCL2 (MCP-1) in dose-dependent manners. 25 These effects of GK1, along with other unexplored innate and adaptive immune mechanisms, could at least, in part, explain the observed anti-tumoral effects of this peptide in an experimental murine model of melanoma. Thus, it is feasible to speculate that IFN-γ could play a central role in the antitumor effects observed in this study, as IFN-γ has been shown to be a mediator of tumor immunity. 34 In this sense, it is significant that B16 melanoma cells treated with IFN-γ were shown to exhibit up-regulated expression of MHC class I and class II, increased expression levels of fas and fas ligands, and greater susceptibility to lysis mediated by non-antigen-specific mechanisms. 35 In addition, IFN-γ is known to mediate antitumor activity through its antiangiogenic effects. 34 Together, these effects might explain the presence of numerous apoptotic tumoral cells and the significantly greater necrotic areas in the GK1-treated mice than in the control mice.
Furthermore, a histological analysis demonstrated an increased inward flow of melanoma-associated neutrophils in the GK1-treated mice, and many of these cells were in close proximity to normal, distorted, or apoptotic tumor cells. It has been known for many years that polymorphonuclear (PMN) leukocytes have the ability to kill tumor targets. 36,37 In addition, potent, specific, and long-lasting antitumor immunity has been reported in murine models, including the B16 melanoma model, 38 and in cancer patients given recombinant cytokines, such as IL-4, IL-3, granulocyte-macrophage colony-stimulating factor (GM-CSF), and MCP-3. These cytokines have been shown to stimulate the production of PMN leukocytes and monocytes in the bone marrow, resulting in extensive tumor destruction after vaccination. 39 –41
Inflammation has been implicated in the promotion and progression of tumors. The production of proinflammatory cytokines, such as TNF-α, and the inflammatory chemokine CCL2 (MCP-1) has been shown to be stimulated by GK1 27 and could have had profound effects on tumor growth and metastasis. However, it remains unknown whether GK1 induces the expression of CCL2 by melanoma cells, and this effect was not evaluated in the present work. This topic is significant, because inflammatory chemokines (CCL2[MCP-1], CCL5 [RANTES], and CXCL8 [IL-8]) appear to regulate the influx of leukocyte sub-populations with pro-tumoral activities, such as tumor-associated macrophages, myeloid-derived suppressor cells, tumor-associated neutrophils, Th17 cells, and Tregs, and these effects are potentiated by TNF-α. 42 However, some tumors secrete CCL2, and it has been proposed that this chemokine (along with others, such as CCL5 and CXCL2 and their receptors) is a critical mediator for the optimal antimetastatic activity of G-CSF-stimulated neutrophils. 43 Furthermore, it is likely that neutrophilia in the peripheral blood, particularly in the pulmonary vessels of GK1-treated mice, is mediated by chemokines. Given the mechanisms of action of GK1, 27 this parasite-derived peptide could have both anti-tumoral and pro-tumoral effects. However, in the present study, we demonstrated that GK1 stimulated anti-tumoral therapeutic action, as reflected by the significant increase (42.58%) in survival time in experimental melanoma-bearing mice treated with GK1, compared with control mice.
Several features of the murine melanoma model support our results. First, the typical dose used was 1×105 B16-F10 cells/mouse, which is 1.5 to 2 times the minimal tumorigenic dose in normal C57BL/6 mice. The tumor cells injected typically form a palpable tumor in 5 to 10 days and grow to reach a tumor size of 1×1×1 cm within 14 to 21 days. 44 In our study, the inoculum was 2×105 tumor cells, which yielded tumors within 2 to 8 weeks of the appropriate size (20 mm3, day 0) in 28 of 32 mice (87.5%), which was similar to the results of a previous report. 45 However, this finding suggests that the murine melanoma model is unpredictable with regard to the specific timeline of tumor development. This unpredictability is particularly relevant for defining a strict criterion for tumor diagnosis (size or volume) and for setting a “day 0” when testing new therapeutic approaches. Moreover, the assessment of survival time will be influenced if treatment is initiated before a well-defined tumor is present.
Interestingly and close linked to previous reports, no tumors were observed in 4 mice for approximately 1 year after injection, representing a 12.5% of unsuccessful in the development of melanoma. This finding could be explained by human data; for example, the percentage of patients who present with partial or complete histological regression of the primary tumor is estimated to be as high as 25% to 40%, although reports of sustained long-term partial regression are rare. 46 –48 Several lines of evidence support a central role of immune mediators in spontaneous melanoma regression. Biopsies of regressing lesions have demonstrated overexpression of T-helper (Th)-1 cytokines, such as IFN-γ, IL-2, and TNF-α, when compared with nonregressing lesions. 49 In addition, spontaneous regression of metastatic melanoma has been associated with evidence of increased immune parameters, such as delayed-type hypersensitivity responses to skin testing. 50
Survival time should be the gold standard for defining success or failure when evaluating a new therapeutic approach. Consequently, it is very important to establish the presence of a tumor using a validated measuring technique. A validated technique should be used, because mouse models of melanoma have high rates of failure and/or spontaneous remission. 45,51 –53 In addition, the time necessary for a tumor to reach a predetermined size is variable. Thus, we suggest that each group should determine its own tumor success/failure rates and that researchers be particularly cautious when documenting survival and the inoculation-tumor interval in trials in which treatment is initiated before tumors are present.
In the current study, the melanomas induced in mice were correlated with human Stage IIb melanomas, which are treated with surgery, according to the TNM classification system of the American Joint Committee on Cancer. 32 In this study, the surgical removal of tumors of 20 mm3, 500 mm3, and >500 mm3 in size led to survival rates at 6 months of 83.3%, 40%, and 0%, respectively. In fact, 40%–60% of surgically treated mice with tumors smaller than 500 mm3 survived 1 year in excellent clinical condition. These survival rates are the highest obtained to date, as previously reported survival rates have ranged from 50 to 120 days after the application of diverse experimental treatments. 11,19,54 Since tumor recurrence occurred in all three groups, adjuvant treatment should be considered after surgical removal; however, we recommend that surgical treatment be performed on tumors 20 to 500 mm3 in size, because the mortality rates due to surgical treatment were low in animals with such tumors. In addition, we recommend adjuvant treatment after surgery in tumor-bearing mice with tumors ≤500 mm3 in size.
To our knowledge, this study was the first to demonstrate that treatment with the synthetic parasite-derived peptide GK1 effectively increased the mean survival time of mice with B16-F10 melanomas. This therapeutic effect could have been related to various immune mechanisms stimulated by GK1, 27 but its precise role as an anti-tumoral molecule remains to be determined. Since GK1 exerts its effects in a dose-dependent manner, and it is rapidly cleared from the body, 25 it has the potential to be used as a primary or adjuvant component of chemotherapeutic cocktails to treat experimental and human cancers. Currently, our group is performing studies to determine whether GK1 induces the same proinflammatory cytokine profiles in mouse melanoma models and whether these effects are dose dependent. In addition, extending our previous results, 29 we are conducting a similar descriptive study to define the potential synergism between GK1 and 4-hydroxycoumarin.
Footnotes
Acknowledgments
The authors are grateful to Ricardo Vargas-Orozco and to veterinarian Daniel A. Sánchez Almaráz from the Experimental Medicine Department (Facultad de Medicina, Universidad Nacional Autónoma de México) for their excellent animal breeding and care and also to María de la Cruz Contreras Guerrero and Verónica Rodríguez from the Department of Cell and Tissue Biology (Facultad de Medicina, UNAM) for their technical assistance. Grant support: The present work was funded by Facultad de Medicina, Universidad Nacional Autónoma de México (Project 040/2009 approved June 2, 2009).
Disclosure Statement
The authors declare no financial or other conflicts of interest.