
Abstract
Background:
Aberrant activation of phosphatidylinositol 3-kinase (PI3K)/Akt/mammalian target of rapamycin (mTOR) pathway may account for development of radioadaptation and is not rare in prostate cancer. Neither PI3K nor mTOR blockade could completely inhibit the pathway owing to paradoxical feedback, so we anticipate dual PI3K/mTOR blockade by NVP-BEZ235 to radiosensitize prostate cancer cells.
Methods:
We investigated into the radiosensitizing effect of NVP-BEZ235 on PC-3 cells, which are devoid of androgen receptors. Clonogenic survival and MTT assays were performed, and to pursue underlying cellular changes flow cytometric analysis of cell cycle and apoptosis as well as western blot were carried out.
Results:
Exposure to NVP-BEZ235 and irradiation caused a greater degree of survival inhibition than ionizing radiation (IR) or BEZ235 alone. Dual PI3K/mTOR blockade along with IR induced a G2/M arrest and enhanced proapoptotic effect. NVP-BEZ235 radiosensitized PC-3 cells through counteracting constitutive as well as IR-triggered activation of Akt/mTOR signaling.
Conclusions:
Our study demonstrated that the dual PI3K/mTOR inhibitor NVP-BEZ235 prominently improved the radiosensitivity of PC-3 cells. It sensitized tumor cells to irradiation via interruption of cell cycle progression and augmentation of cell apoptosis, which was due to its constraint on constitutive and IR-elicited PI3K/Akt/mTOR signaling activation.
Introduction
Prostate cancer is the most common malignant neoplasm in male urinary system. Combined management for prostate cancer has been developed, among which radiotherapy is a most important modality of treatment. Nevertheless, the intrinsic or acquired radioresistance of prostate cancer has presented itself as the main obstacle to achieve long-term control, despite remarkable advances achieved in radiation facilities and technology. That provides a strong rationale for exploration into potential radiosensitizing agents of prostate cancer.
Phosphatidylinositol 3-kinase (PI3K)/Akt/mammalian target of rapamycin (mTOR) make up a vital pathway that regulates cell cycle, proliferation, apoptosis, and biosynthesis process in eukaryocytes. 1 Indicated by numerous studies and investigations in recent years, the aberrant activation of PI3K/Akt/mTOR pathway, either inherent or induced, plays an indispensable role in neoplastic origination, metastasis, and development of radioadaptation. 2 –4 Over-activation of the pathway often occurs through mutation of coding genes (PIK3CA, AKT1, AKT2 etc.), constitutive activation of up-stream molecules, such as epidermal growth factor receptor (EGFR) and Ras, as well as the depletion of phosphatase and tensin homologue deleted in chromosome 10 (PTEN), most of which are not rare in prostate cancer. 5
The kinase mTOR forms two distinct complexes known as mTORC1 and mTORC2. The former consisting of mTOR and raptor (regulatory-associated protein of mTOR) can be inhibited by rapamycin, while the latter, including mTOR and rictor (rapamycin-insensitive companion of mTOR) is insensitive to rapamycin. 5 The inhibition of mTORC1 alone, as in the case of rapamycin, can lead to mTORC2-mediated feedback activation of Akt on Ser473. 6 The existence of the paradoxical feedback loop may abrogate the efficacy of mTORC1 inhibitors and thus, rationalize the application of dual inhibitors.
NVP-BEZ235 is a novel antitumor drug developed by Novartis Pharma, which functions as a dual PI3K/mTOR inhibitor. 7 Preclinical studies have described its antiproliferative effect in several cell lines, especially those with PI3K or K-Ras mutation. 7,8 Of note, NVP-BEZ235 has been demonstrated to improve the sensitivity of cancer cells to chemotherapy, indicating its potential role as a sensitizing agent in combined modalities of treatment. 9
Since the proapoptotic activity of ionizing radiation (IR) can be compromised by IR-triggered activation of PI3K/Akt/mTOR pathway, 3,4,10 –13 we are justified to hypothesize that dual PI3K/mTOR inhibition would sensitize prostate cancer cells to radiation therapy. Here in this study we examined the capacity of NVP-BEZ235 to reverse the radiation resistance of prostate cancer cell line PC-314–17 and explored possible molecular mechanism. It was concluded that dual PI3K/mTOR blockade by NVP-BEZ235 could synergize with IR in PC-3 cells. The underlying mechanisms may include cell cycle arrest and enhancement of IR-induced apoptosis.
Materials and Methods
Cell culture and reagents
The human prostate cancer cell line PC-3 was obtained from ATCC. Cells were cultured in RPMI-1640 (Hy Clone; Thermo Scientific) with 10% of fetal bovine serum, and incubated at 37°C in 5% CO2/95% air.
Dual PI3K/mTOR inhibitor treatment
NVP-BEZ235 was obtained from Novartis Pharma. The inhibitor was added to mid-log-phase cell cultures at the indicated concentrations. After treatment, the culture medium was replaced with drug-free medium. For the control group, cells were incubated with medium containing dimethyl sulfoxide (DMSO) at a concentration corresponding to the highest dose used in inhibitor-treated cells.
Clonogenic survival assay
The exponential growth cells (500-1000/well) were seeded into six-well plates. 12 hours after plating, NVP-BEZ235 was administered at concentrations of 50 nM and 100 nM 1 hour before irradiation. The cells were then exposed to IR at doses of 2, 4, 6, and 8 Gy at an average dose rate of 200 cGy/min. Twenty hours after irradiation, the medium was replaced by basal culture medium free of the inhibitor. The cells were incubated for 7 to 10 days at 37°C in 5% CO2/95% air to allow the colony formation. The plates were fixed with pure methanol and stained with 1% crystal violet. We scored colonies of more than 50 cells and graphed the surviving fraction (SF) versus dose of IR. SF was estimated by the following formula: SF=number of colonies formed/number of cells seeded×plating efficiency. Plating efficiency is the ratio of colony number of the control group to cells seeded. Dose–response clonogenic survival curves were plotted on a log-linear scale. Three cell concentrations were applied for each dose of IR. D0 and Dq values, reflecting the sensitivity and the shoulder of the survival curve, were calculated as described. 18
MTT assay
Cells (1×104/well) were seeded into 96-well plates and incubated for 24 hours to allow adherence. NVP-BEZ235 was administered at concentrations of 50, 150 and 250 nM 1 hour before irradiation. The plates were then exposed to IR at doses of 2, 4, 6, 8 and 10 Gy at an average dose rate of 200 cGy/min. 20 hours after irradiation, the medium was replaced by basal culture medium free of the inhibitor. Thereafter, 10 μL 5 mg/mL MTT was added into each well and the cells were incubated for another 4 hours before termination of experiment. The medium was discarded and 100 μL DMSO was added in each well to dissolve the formazan produced by viable cells. OD values at 490 nm were measured in a microplate reader. Then the rate of inhibition was calculated using the formula: rate of inhibition=1−OD value of the experimental group/OD value of the control group.
Flow cytometric analysis for cell cycle and apoptosis
Tumor cells were incubated overnight to allow adherence. The inhibitor was added 1 hour before exposure to IR at a single dose of 4 Gy. Twenty hours after irradiation, the medium was replaced by basal culture medium free of the inhibitor. The cells were then cultured for another 4 or 28 hours before harvesting. Cell cycle analysis was carried out according to a standard procedure in Cell Cycle Flow Cytometry Testing Kit (Genmed Scientifics, Inc.). Analysis of apoptosis was performed according to a standard procedure in Annexin V-FITC/PI Apoptosis Detection Kit (ABGAB). Afterwards the samples were subjected to assessment with a FACSCalibur flow cytometer (Becton Dickinson). The data representative of three independent experiments were analyzed with FlowJo v 7.6.5 software.
Western blot
Western blot was performed as described earlier. 19 The expression of actin was set as the protein loading control. Antibodies against phosphorylated-(P)-(Ser473)-Akt, P-(Thr308)-Akt, total Akt (T-Akt), P-mTOR, p21cip1, p27kip1, cyclin A, cyclin B, cyclin E, actin as well as horseradish peroxidase-conjugated secondary antibodies were from Abcam.
Statistical analyses
All data are presented as the average±SD. Experiments were conducted in triplicate. Significance of differences was determined using two-tailed unpaired Student's t test or the one-way ANOVA test with SPSS 13.0 software. A value p<0.05 was considered statistically significant.
Results
The dual PI3K/mTOR inhibitor NVP-BEZ235 reversed radioresistance of PC-3 cells and sensitized the tumor cells to the proapoptotic effect of IR
To evaluate the radiosensitizing effect of the dual PI3K/mTOR inhibitor, MTT and clonogenic survival assay were performed. Compared with the control group that excluded treatment of NVP-BEZ235, tumor cells exhibited higher rate of inhibition under the same IR dose after dual PI3K/mTOR inhibition, which demonstrated the radiosensitizing activity of the inhibitor (Fig. 1A). Notably, with the increase of inhibitor concentration, a climbing degree of inhibition was observed, suggesting that the sensitizing effect was also inhibitor dose-responsive (Fig. 1B). Besides, combination of IR (4 Gy) and NVP-BEZ235 (50 nM) resulted in a greater degree of inhibition on cell viability than treatment with IR or NVP-BEZ235 alone (p<0.05), implying a synergistic action between the agent and irradiation (Fig. 1C).

Effect of combination of NVP-BEZ235 and IR on cell viability in PC-3 cells. Cells were seeded into 96-well plates and were exposed to NVP-BEZ35 at indicated concentrations 1 hour before irradiation. Twenty hours after IR the medium was replaced by basal culture medium and cell viability was assessed by MTT assay.
The clonogenic survival assay was carried out to estimate the survival rate of clonogenic tumor cells after different means of interference. The tumor cells were exposed to increasing doses of IR (2–8 Gy) with/without the presence of the inhibitor. According to preliminary experiments that were intended to determine the optimal incubation time with NVP-BEZ235, continuous treatment with the inhibitor led to failure of colony formation, while incubation for no more than 21 hours didn't affect the plating efficiency significantly. Therefore, for subsequent assays we exposed PC-3 cells to the inhibitor 1 hour before IR and limited the total incubation time to 21 hours. Pretreatment with NVP-BEZ235 resulted in an obvious reduction in the survival rate of tumor cells after exposure to different doses of IR. An increase of the radiosensitizing effect was uncovered accompanying the elevation of inhibitor concentration (Fig. 2A).
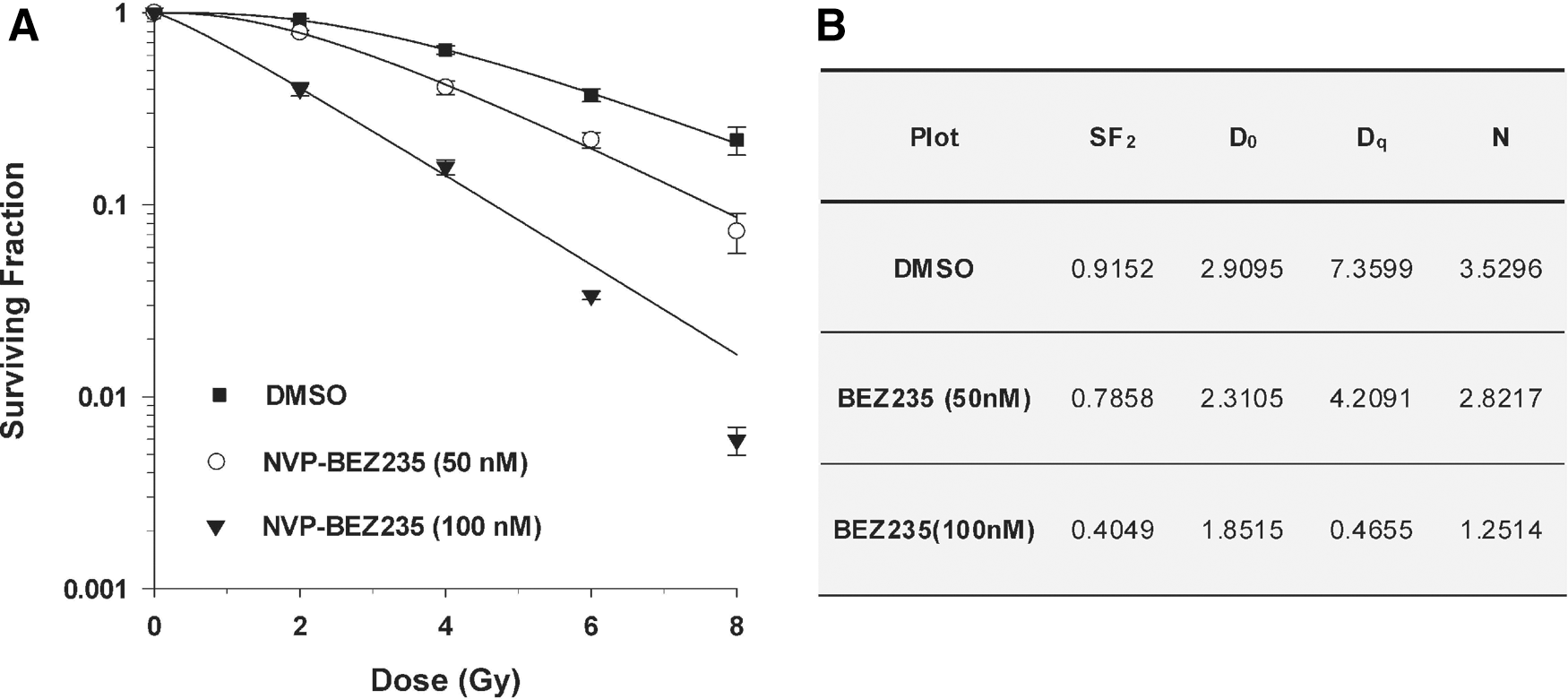
Clonogenic survival curves
To derive a quantified evaluation of the effect, the data were fitted into single-hit multitarget curves and some relevant parameters were calculated. Addition of the dual inhibitor apparently decreased the mean lethal dose D0 as well as the quasi-threshold dose Dq, suggesting the radiosensitivity of PC-3 cells was improved by the inhibitor. Also not surprisingly, the elevation of the inhibitor concentration brought about a further decline in the radioresistance of the tumor cells (Fig. 2B). All those stated above proved that NVP-BEZ235 was indeed able to sensitize PC-3 cells to IR, and most probably by augmenting DNA damage caused by IR or interfering with the cell cycle checkpoints.
Dual PI3K/mTOR blockade along with IR altered cell cycle distribution and induced a G2/M arrest via cell cycle modulating proteins
To live out whether the dual PI3K/mTOR inhibitor radiosensitized PC-3 cells via redistribution of cell cycle, we conducted cell cycle assay on tumor cells exposed to IR and/or the inhibitor. Cell cycle was detected at 24 and 48 hours after irradiation, respectively to help reveal any time-related alteration. NVP-BEZ235 (50 nM) alone resulted in growth arrest in the G1 phase of cell cycle, whereas IR alone led to the G2/M arrest. Strikingly enough, the combination of IR with the inhibitor induced a further accumulation of PC-3 cells in the G2/M phase, which highlights the potential mechanism of NVP-BEZ235 as a radiosensitizing agent. Different time course points at assay didn't exert profound influence on the distribution of cell cycle (Fig. 3).

Evaluation of cell cycle distribution in PC-3 cells. Cells were harvested at 24 hour
Further western blot detection helped reveal cellular changes underlying different actions of IR and BEZ235 (Fig. 5). In accordance with previous observations, IR alone caused a G2/M arrest in PC-3 cells. Meanwhile irradiation promoted the expression of p21 and p27, both of which are vital cyclin-dependent kinase (CDK) inhibitors, resulting in diminished levels of cyclin E and B (Fig. 5B). Unlike IR, NVP-BEZ235 alone elicited G1/S detention. Increased level of p21 as well as p27 was observed to concur with dramatic decrease of cyclin E and A, all of which participate in G1/S checkpoint (Fig. 5B). Though exerting opposing effect on cell cycle compared with IR, NVP-BEZ235 unexpectedly primed more cells to G2/M arrest when combined with IR. Consistently changes in p21 and cyclin B were detected (Fig. 5B).
NVP-BEZ235 radiosensitized PC-3 cells through enhancement of proapoptotic effect of IR
Irradiation causes cyto-lethal effect mostly by DNA damage and programmed cell death, namely apoptosis. Yet on the other hand, IR-induced activation of the PI3K/Akt/mTOR pathway may finally counteract the proapoptotic outcome of IR by initiation of antiapoptosis cascade. Hence, we were fully convinced that dual PI3K/mTOR blockade would cut off this pernicious activation and augment proapoptotic action of IR. Detection of apoptotic PC-3 cells after exposure to IR and/or NVP-BEZ235 showed that combined treatment led to a significantly higher fraction of apoptosis. Tests were done at 24 and 48 hours after IR, respectively. Quite intriguing results were obtained. The DMSO or NVP-BEZ235 group saw no prominent difference between two time points of detection, while the proportion of apoptotic cells in IR group lowered at 48 hours after irradiation than 24 hours, implying the occurrence of anti-apoptosis events, such as repair of DNA damage. The combination of IR with the inhibitor led to a relatively constant fraction of apoptosis despite delay of the inspection point (Fig. 4).
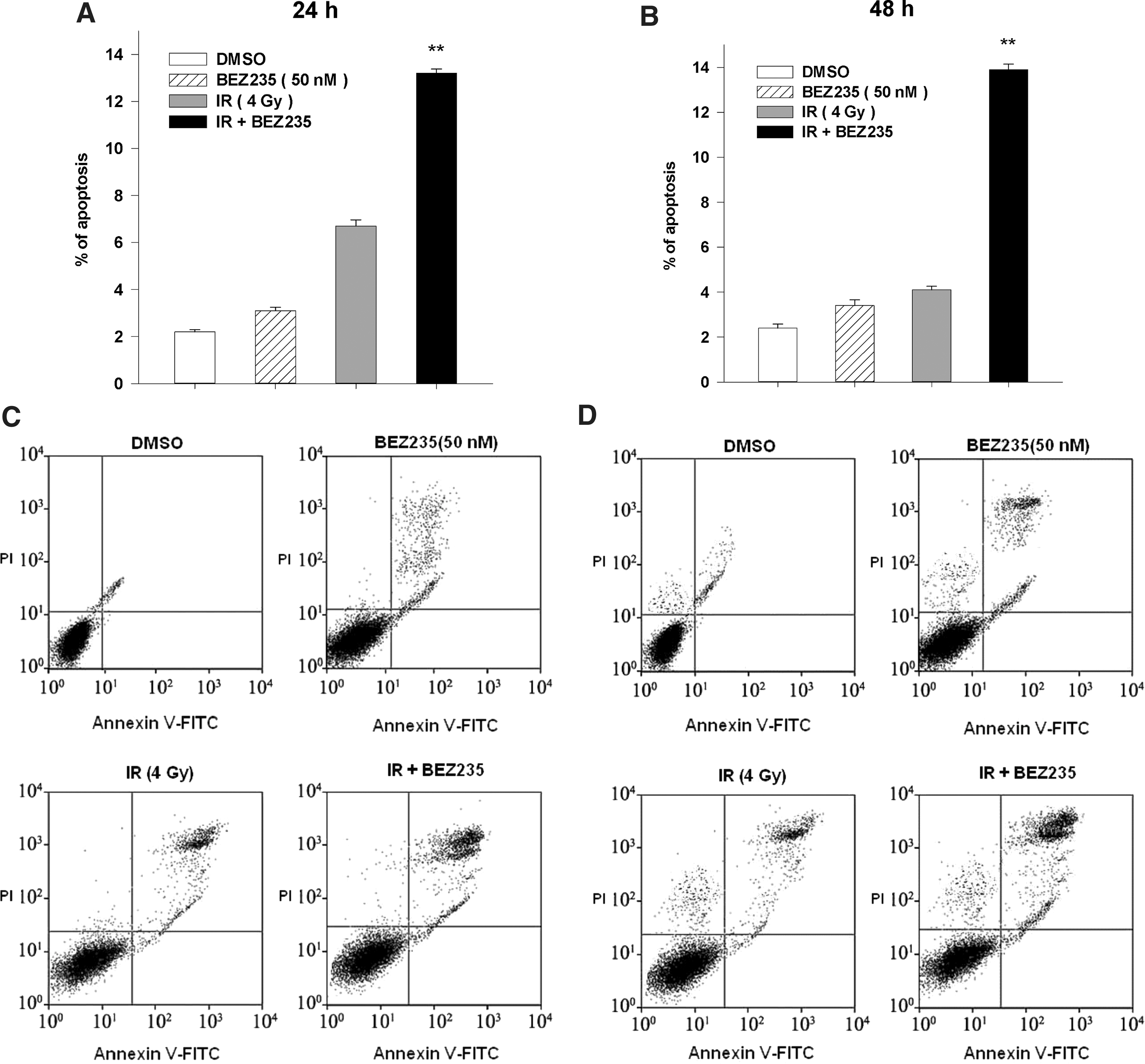
Analysis of apoptosis in PC-3 cells by flow cytometry. Cells were harvested at 24 hours
NVP-BEZ235 counteracted constitutive as well as IR-triggered activation of Akt/mTOR signaling in PC-3 cells
Western blot assay was carried out to throw light upon potential radiosensitizing mechanism of NVP-BEZ235. As anticipated, IR induced further phosphorylation of Akt (on both S473 and T308) and mTOR, while the total content of Akt remained stable. When applied alone, NVP-BEZ235 suppressed intrinsic dysfunction of Akt/mTOR signaling although the effect was not a notable one. Combination of the agent with IR created a desirable compromise of IR-induced response, cracking down the activity of the pathway to the basic level (Fig. 5A).

Detection of relevant proteins involved in cellular responses to IR and/or NVP-BEZ235 treatment by western blot analysis.
Discussion
As indicated previously, intrinsic and IR-induced activation of PI3K/Akt/mTOR pathway in prostate cancer cells is considered to compromise the cytotoxic effect of irradiation and mostly accounts for the development of radioresistance. 20 –22 Rapamycin analogues (rapalogs) have been discovered to exhibit anti-proliferative effect in some tumor cell lines, 23 but with limited efficacy in tumors that have PI3K/Akt and mTOR activation, for they only inhibit mTORC1 rather than mTORC2, which can phosphorylate Akt at Ser473 and eventually restart the deregulation of PI3K/Akt pathway. 24,25 As a dual PI3K/mTOR inhibitor, NVP-BEZ235 not only inhibits the activity of PI3K and mTORC1 but also effectively blocks mTORC2. 7 Therefore, we have every reason to expect NVP-BEZ235 to abrogate the cell survival response elicited by IR and reverse the radioresistance of prostate cancer. 14 –17
Former research work has revealed that prostate cancer can be divided into two categories, namely androgen receptor (AR) positive and AR-negative. 26 Development of abnormal AR activation with the deprivation of androgen accounts for the failure of castration therapy commonly seen in the clinical setting. 27 AR activation finally directs to a series of cellular pathways that reportedly cross-talk with the PI3K/Akt/mTOR pathway. 28,29 Here in our study PC-3 cells, which are AR-negative, 26 were opted so that influence from AR pathways could be avoided.
In some preclinical models, NVP-BEZ235 have showed potent antiproliferative activity in tumors, such as renal cell carcinoma, glioblastoma, colorectal, and breast cancer. 8,30 It has been proposed as a promising antitumor agent owing to its properties as orally-available and well-tolerated. 7 Nevertheless description of NVP-BEZ235 being applied to radiosensitize tumors has been scarce. Our work has provided insight into this inhibitor as a radiosensitizing agent in prostate cancer, which possesses poor radiosensitivity 14 –17 and high occurrence of PI3K/Akt/mTOR pathway deregulation. Here in this study we demonstrated that when used at minimal concentration NVP-BEZ235 already displayed remarkable radiosensitizing activity, and with the increase of inhibitor dose such effect was amplified. When administered alone at low concentration, the dual inhibitor had little impact over the proliferative activity of PC-3 cells, supplying further evidence for its role as a radiosensitizing agent.
Actually we did conduct preliminary experiments to determine the best treatment schedule for administration of NVP-BEZ235 (data not shown). The drug was applied 24 hours/12 hours/6 hours/1 hour before, coinstantaneously with and after IR, yet, the latter two arrangements failed to bring about any positive sensitizing effect, revealing that complete blockade of target kinases requests pre-exposure to NVP-BEZ235 and thus, suppresses constitutive as well as IR-induced PI3K/Akt activation. Among all those schedules giving NVP-BEZ235 before IR, 1 hour of pre-exposure followed by IR demonstrated the most prominent radiosensitizing effect, while preincubation for 12 and 24 hours before IR only caused a mild to moderate degree of cytotoxicity, similar to that of IR alone. That timing-dependent sensitizing effect of NVP-BEZ235 provided cues for our decision on the treatment schedule.
To confirm the cellular changes concurring with the radiosensitizing action of the inhibitor, we implemented concerning experiments on cell cycle analysis and detection of apoptosis, from which we were compelled to conclude that NVP-BEZ235 abated the radioresistance of PC-3 cells by inducing cell cycle arrest and proapoptotic response. In accordance with previous findings about NVP-BEZ235 and other PI3K inhibitors, NVP-BEZ235 alone aroused the G1 phase block. More importantly, exposure of PC-3 cells to clinically relevant dose of IR with NVP-BEZ235 pretreatment resulted in the detention of cancer cells in G2/M phase. Similarly, rapalogs, namely rapamycin analogues, have been advanced to produce G2/M phase arrest along with irradiation. 31 Detection of cell cycle regulating proteins supplied hints as to the observed changes. What should be noted here is that the main components involved in G1/S and G2/M checkpoints were assessed except p53 since PC-3 cells are p53 null. Interestingly, contrary to IR, NVP-BEZ235 alone induced G1/S arrest by modulation of p21, p27, cyclin E, and A. Yet, when followed by irradiation, BEZ235 apparently further promoted IR-triggered G2/M arrest instead of combating it. Why such seemingly contradictory consequences occurred still needs more pursuit, but it seems safe to conclude that p53-independent p21 and p27 activity indeed are inducible in PC-3 cells.
In addition, the influence of the dual inhibitor on cellular apoptosis was investigated. Prostate cancer cells were subjected to NVP-BEZ235 and/or IR. We also drew a comparison between cells collected at different time points of detection. Here from our statistics it was clearly displayed that the combination of the dual inhibitor and IR led to greater degree of cellular apoptosis compared with the separate action of the agent or IR. Particularly, when the samples were examined at 48 rather than 24 hours after IR; thus, sparing the cancer cells adequate time to initiate inner prosurvival response, a paradoxical descent in the percent of apoptosis was observed. This conferred persuasive evidence for the occurrence of IR-elicited antiapoptotic events, which partly offset the cytotoxic action of IR. On the contrary, such diminishment of cellular apoptosis with time delay was not seen in the group enduring combined treatment, and the level of apoptosis was slightly elevated instead of being lowered as time went by. We considered this observation a vital one, because it provided hints that NVP-BEZ235 most probably sensitized PC-3 cells to irradiation by constraining the IR-evoked cell survival reaction.
Consensus has been reached that the PI3K/Akt/mTOR pathway is implicated in cell proliferation and synthesis, 32 and IR-induced activation of the pathway may help explain the acquired insensitivity of PC-3 cells to irradiation, 20 which was demonstrated in the present study. We initially assumed complete and specific blockade of the PI3K/mTOR pathway by NVP-BEZ235 would ameliorate tumor sensitivity to IR via suppressing pernicious prosurvival cell response, and the western blot results finally bore out that hypothesis. Given that cell cycle and apoptosis related proteins belong to PI3K/Akt/mTOR signaling downstream targets, it may be justified to conclude that NVP-BEZ235 actually played its radiosensitising role by direct inhibition of PI3K/Akt/mTOR pathway and subsequent restraint of cell cycle progression and anti-apoptotic events. Yet, to detail the molecular mechanism involved in this interwoven network, such as the changes in some apoptosis-concerning markers, more fundamental experiments need to be carried out in our future exploration into this subject.
Apart from interference of cell cycle regulation and facilitating proapoptotic events, other molecular mechanisms underlying the potent radiosensitizing efficacy of NVP-BEZ235 have been proposed. Specific blockade of PI3K and mTOR activity by NVP-BEZ235 has been challenged by a recent study. Mukherjee et al. 33 reported that NVP-BEZ235 not only effectively inhibited PI3K/mTOR, but also exerted cross-reactivity toward ATM and DNA-PKcs, which belong to the PI3K-like kinase (PI3KK) family and mainly function in response to IR-induced DNA double-strand breaks (DSBs). This inevitably led to the attenuation of DSB repair, including nonhomologous end joining (NHEJ) and homologous recombination (HR), mediated by DNA-PKcs and ATM, respectively. Moreover, phosphorylation of ATM downstream targets as well as G2/M arrest were also abated by NVP-BEZ235. Altogether these effects conferred by NVP-BEZ235 resulted in its dramatic radiosensitizing action on glioblastoma. The current study in PC-3 cell line failed to bear evidence of DSB repair defects after NVP-BEZ235 administration, but whether or not modulation of multiple PI3KK family members by NVP-BEZ235 is ubiquitous in various classes of carcinoma warrants further investigation notwithstanding. Taking into account all that has been discussed, it's reasonable to arrive at the conclusion that NVP-BEZ235 indeed performs a strikingly potent radiosensitizing effect by disabling numerous cellular signaling pathways correlated with (1) cell cycle arrest owing to PI3K/Akt and ATM inhibition, (2) cell proliferation and survival due to PI3K/Akt/mTOR blockade, (3) DSB repair as a result of ATM/DNA-PKcs and less importantly PI3K/Akt inhibition.
In the current setting, NVP-BEZ235 and several other dual PI3K/mTOR inhibitors are still in phase I/II clinical trials. In this regard, our work added to its preclinical data and expanded its spectrum of application as not only a novel antitumor drug but also a radiosensitizing agent. Its radiosensitizing action has been determined in several cancer cell lines, 34 but our study pioneered the investigation of its sensitizing effect on prostate cancer, which is of clinical significance because prostate cancer is generally acknowledged to be radioresistant.
We arrived at a primary conclusion that NVP-BEZ235 indeed radiosensitized prostate cancer cells possibly through inhibition of cellular prosurvival stress response and enhancement of apoptosis caused by irradiation. For further look into this subject, subsequent experiments are necessary. Our data were gained from AR-negative PC-3 cell to avoid potential interaction between AR pathway and the target pathway. In future, we intend to investigate action of NVP-BEZ235 on AR-positive prostate cancer cell lines, such as LNCaP cell to throw light upon crosstalk between these two pathways. Androgen-independent AR activation eventually renders prostate cancer cells resistant to androgen deprivation therapy, and to make things worse prostate cancer is by nature insensitive to irradiation, resulting in a complicated but commonplace clinical circumstance. Research work stated above will no doubt present solutions of practical importance to this dilemma. Last but not least, animal models can be established to evaluate the radiosensitizing effect of NVP-BEZ235 in vivo, which will produce more direct insight into its practical efficacy.
In summary, our study demonstrated that the dual PI3K/mTOR inhibitor NVP-BEZ235 prominently improved the radiosensitivity of PC-3 cells in which aberrant activation of PI3K/Akt/mTOR pathway was common. It sensitized tumor cells via interruption of cell cycle checkpoints and augmentation of cell apoptosis, which was due to its constraint on IR-elicited prosurvival cellular response. In the future, we intend to obtain insight into potential molecular mechanisms associated with the radiosensitizing effect of dual PI3K/mTOR blockade, and optimize its clinical application via in vivo experiments.
Footnotes
Disclosure Statement
NVP-BEZ235 produced by Novartis Pharma is purchased from LC labs, which does not compromise the study scientifically.