
Abstract
The irradiated cells communicate with unirradiated cells and induce changes in them through a phenomenon known as the bystander effect. The nature of the bystander signal and how it impacts unirradiated cells remains to be discovered. Examination of molecular changes could lead to the identification of pathways underlying the bystander effect. Apart from microRNAs, little is known about the regulation of other non-coding RNAs (ncRNA) in irradiated or bystander cells. In this study we monitored the transcriptional changes of several small nucleolar RNAs (snoRNAs) host genes and long non-coding RNAs (lncRNAs) that are known to participate in a variety of cellular functions, in irradiated and bystander cells to gain insight into the molecular pathways affected in these cells. We used human lymphoblasts TK6 cells in a medium exchanged bystander effect model system to examine ncRNA expression alterations. The snoRNA host genes SNHG1 and SNHG4 were upregulated in irradiated TK6 cells but were repressed in bystander cells. The SNHG5 and SNHG11 were downregulated in irradiated and bystander cells and the expression levels of these ncRNA were significantly lower in bystander cells. The lncRNA MALAT1, MATR3, SRA1, and SOX2OT were induced in irradiated TK6 cells and their expression levels were repressed in bystander cells. The lncRNA RMST was induced in both irradiated and bystander cells. Taken together, these results indicate that expression levels of ncRNA are modulated in irradiated and bystander cells and these transcriptional changes could be associated with the bystander effect.
Introduction
The traditional paradigm of radiation-induced effects is based on cells directly damaged by the radiation. Current evidence suggests that in cell populations exposed to ionizing radiation, biological effects occur in a much larger proportion of cells than are estimated to be traversed by radiation. The irradiated cells are capable of providing signals to the neighboring cells and that signal results in damage to nearby unirradiated cells. This phenomenon has been termed as the bystander effect. Evidence for a bystander effect in irradiated cell populations was first reported in 1992. 1 Direct evidence of the bystander effect was provided in studies where the transfer of medium from irradiated cells was shown to induce cell killing of unirradiated bystander cells. 2
Many studies have investigated the biological responses in bystander cells. An increase in the frequency of micronuclei formation, sister chromatid exchanges, gene mutations, and chromosomal instability were seen in bystander cells. 1,3,4 Cell proliferation and micronuclei induction were found in unirradiated cells cocultured with irradiated cells. 5 Radiation-induced bystander mutagenesis in human lymphoblast cells has been described. 6 Changes in nuclear and mitochondrial gene expression have been observed in bystander cells. 7 –9 The existence of bystander effect after high linear energy transfer (LET) radiation, such as α-particles is very well documented. 10 Recent studies have shown that low LET X-irradiation is also capable of inducing medium-mediated bystander effects. 11
The mechanisms underlying the bystander effects remain unknown. It has been suggested that the production of reactive oxygen species, direct cell-to-cell signaling via gap junctional intercellular communication or soluble factor(s) released into the medium surrounding irradiated cells could be possible mechanisms responsible for the bystander effect. 1,2,12,13
This study was undertaken to examine the molecular processes by which the IR exposure may elicit bystander responses. Our goal was to examine non-coding RNAs (ncRNA) expression in irradiated and bystander cells. Changes in ncRNA levels have been linked to many human diseases. We recently published the modulation of miRNA in γ-radiation-treated human cells. 14 Later studies from our laboratory examined the impact of radiation dose, dose rate, cellular sensitivity to radiation, and DNA repair capability of the cell on the modulation of miRNA in low LET irradiated human cells. 15 –18 We also showed that the miRNA expression levels are altered in bystander cells. 17 In this study, we asked whether other ncRNAs beside miRNA are modulated in irradiated and bystander cells. If so, are there differences or similarities in the ncRNA responses in directly irradiated and bystander cells. We focused on small nucleolar RNAs (snoRNAs) host genes, and long ncRNAs (lncRNAs). The snoRNAs are a subset of ncRNA with a wide variety of cellular functions, such as chemical modification of RNA, pre-RNA processing, and control of alternative splicing. 19 Most snoRNAs are encoded from host genes and are processed from introns of premRNAs. 20 Long non-coding RNAs (lncRNAs) are more than 200 nucleotide in length and lack an open reading frame. 21 LncRNAs are expressed in a disease-, tissue- or developmental stage-specific manner suggesting their specific functions in development and diseases. 22 LncRNAs play an important role in regulating gene expression at various levels, including chromatin modification, transcription, and post-transcriptional processing. 22,23
X-irradiation can induce medium-mediated bystander responses in unirradiated human TK6 cells cocultured with irradiated cells in a transwell insert culture dish. 4,11 In the present study, we asked whether medium-mediated bystander effect in TK6 cells is accompanied by alterations in the ncRNA expression levels. To our knowledge the modulation of ncRNA in bystander cells cocultured with irradiated cell conditioned medium has not been reported. Here, we show differential ncRNA expression responses between irradiated and bystander cells. Our data indicate that irradiated cells have the ability to induce changes in the ncRNA expression levels of bystander cells.
Materials and Methods
Ionizing radiation treatment and cell coculture
The human lymphoblast cell line TK6, kindly provided by Dr. Howard Liber were irradiated with a RAD Source 2000 X-ray Biological irradiator. The cells were seeded at a density of 3×106 per mL and a dose of 2 Gy at a dose rate of 1.7 Gy/min was administered at room temperature. The cell culture Transwell inserts (Becton Dickinson Biosciences) were used to study the bystander effect. 11 The bottom of the insert is a membrane with 1 μm pores at a density of 1.6×106/cm2 to allow the passage of molecules. The polyethylene terephthalate membranes are highly permeable and allow increased rate of basolateral diffusion of nutrients and molecules of interest for transport, secretion, or binding studies. The transwell culture insert dish has a growth area of 4.2 cm2, and the companion well of a six-well plate has a growth area of 9.6 cm2. The distance from the membrane of the insert dish to the bottom of the well of the companion plate is 0.9 mm.
The cells were plated into six-well dishes at a density of 2×105/mL. The inserts containing 2×105/mL, irradiated cells were put into the wells of dishes and the six-well plates along with inserts were returned to the incubator. The irradiated and bystander cells were cocultured to allow the induction of X-ray-induced bystander effects until the time at which cells were harvested. The control sample was treated in the same way, except for irradiation. The treated cells and bystander cells were incubated at 37°C and harvested at 0, 0.5, 4, 8, 12, and 24 hours for isolating RNA. The experiment was repeated in triplicate.
Non-coding RNA
Assays-on-demand for SNHG1, SNHG4, SNHG5, SNHG6, SNHG7, SNHG8, SNHG11, MALAT1, MATR3, SRA1, SOX2OT, and RMST (Table 1) were purchased from Applied Biosystems Foster City. Standard TaqMan assays have been designed using PrimerExpress software. RNA samples for gene expression analysis were normalized based on the TaqMan Gene Expression Assays for human endogenous hypoxanthine phosphoribosyl transferase (HPRT) gene.
Quantitative real-time polymerase chain reaction and data analysis
The RNA isolation, reverse transcription, cDNA synthesis, and quantitative real-time polymerase chain reaction (QPCR) were performed as previously described. 24 The relative expression values of cycle thresholds were calculated by using the comparative delta delta cycle threshold, ΔΔCT method 25 by normalization to the endogenous control HPRT and to the control unirradiated sample. The statistics and data analysis was performed with ABI prism and GraphPad Prism 5 software. CT values of the ncRNA in irradiated and bystander cells were statistically evaluated using a one-way t-test. The experiments were repeated three times and the expression differences between the irradiated and bystander cells were statistically determined.
Results
We were interested in examining the response of ncRNA in TK6 and bystander cells after IR treatment. We searched commercially available assays on demand for TaqMan-based expression monitoring of ncRNA. A number of ncRNA assays on demand were obtained from Applied Biosystems. The testing of 21 assays in TK6 cells revealed that 9 of these ncRNA were not expressed in these cells with or without radiation exposure (data not shown). Following these observations we made a short list of 12 ncRNA for further analysis in irradiated TK6 and bystander cells. The selection of these ncRNA was purely based on the commercial availability of the reagents for their expression analysis and the observation that these ncRNA were expressed in the cell line under investigation.
TK6 cells were irradiated with 2 Gy dose of X-rays and cocultured with unirradiated cells using a Transwell system 4,11 to study the ncRNA responses in both irradiated and bystander cells. At 0, 0.5, 4, 8, 12, and 24 hours time points the irradiated and bystander cells were harvested. The relative gene expression was determined by the QPCR method. The results obtained with the analysis of various snoRNA host genes SNHG1, SNHG4, SNHG5, SNHG6, SNHG7, SNHG8, and SNHG11 are shown in Figure 1. The SNHG1 repression was observed at the 0.5 hour time point relative to the control and repression of SNHG4 was observed at the 8 hours time point (Fig. 1A, B). The expression level of SNHG4 was markedly declined in both irradiated TK6 cells and TK6 bystander cells at the 8 hours time point (Fig. 1B). The SNHG4 was induced at 12 hours in irradiated cells and remained upregulated until 24 hours time point. The expression of SNHG4 was significantly different in bystander cells compared with irradiated cells at 12 and 24 hours time points (p=0.03). The expression of SNHG5 (Fig. 1C) was significantly lower in bystander cells at 0.5, 4, and 8 hours time points (p=0.05). The expression pattern of SNHG6, SNHG7, and SNHG8 was not statistically different among irradiated and bystander cells (Fig. 1D–F respectively). The expression of SNHG11 was significantly lower in bystander cells (p=0.05) at 8 and 12 hours time points (Fig. 1G).
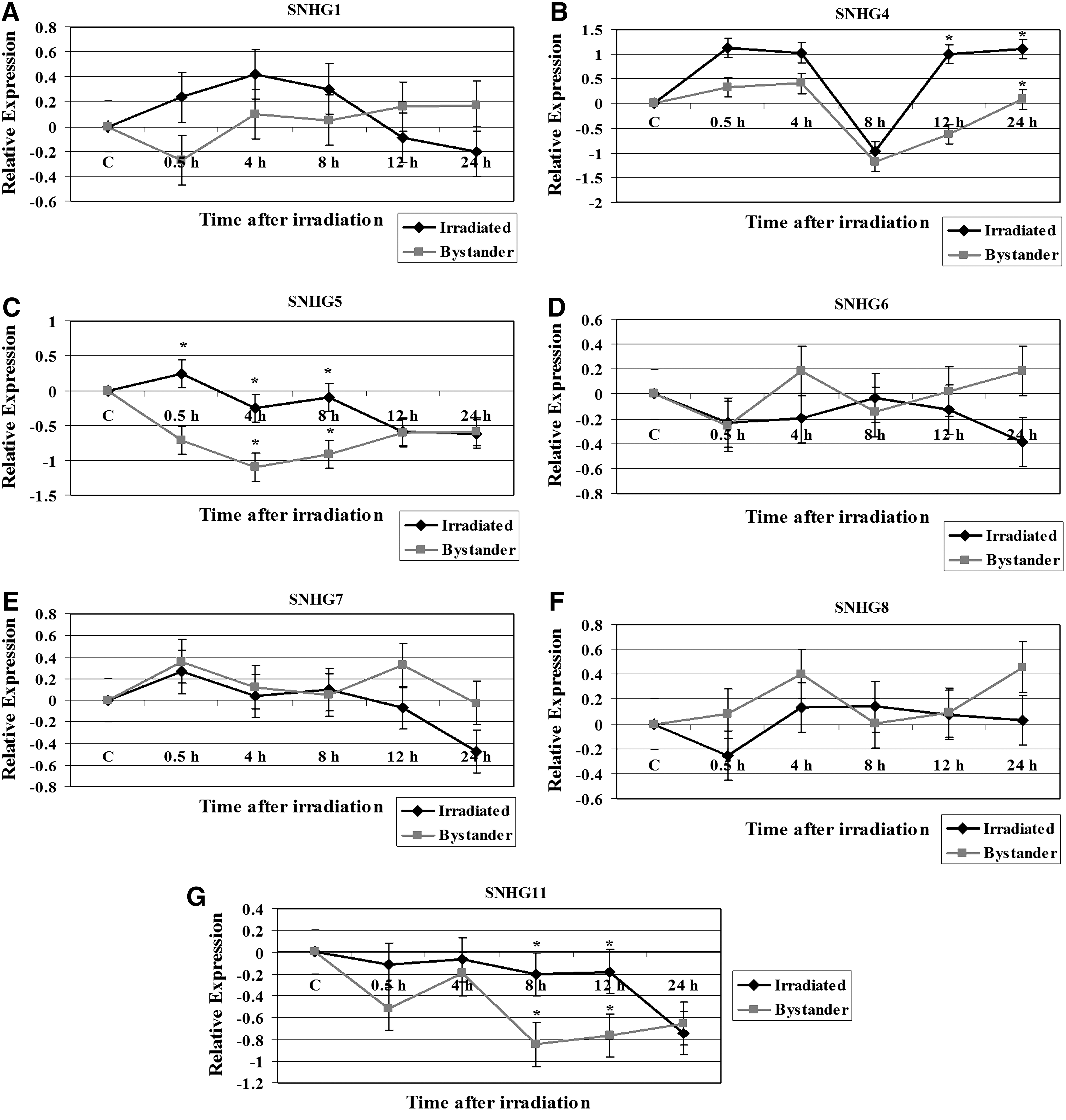
Modulation of various small nucleolar RNA host genes in irradiated TK6 and bystander cells. The relative expression, shown as Log2 values, was computed at 0, 4, 8, 12, and 24 hours time points.
We next examined the modulation of several lncRNA in irradiated TK6 and their bystander cells. The results for MALAT1, MATR3, SRA1, SOX2OT, and RMST expression alterations are shown in Figure 2. While MALAT1 was induced in irradiated TK6 cells, its expression level was repressed in bystander cells during the 8 to 24 hours time points (Fig. 2A). The MALAT1 expression was significantly different in bystander cells compared with the irradiated TK6 cells (p=0.03) at 8, 12, and 24 hours time points. The analysis of MATR3 expression in TK6 cells exposed to X-rays and the cocultured bystander cells indicated the induction of this ncRNA until 8 hours in irradiated TK6 cells, followed by a decline in its expression levels (Fig. 2B). MATR3 remained repressed in bystander cells. The SRA1 expression was also induced in X-rays-treated TK6 cells but was downregulated in bystander cells (Fig. 2C). The differences in the SRA1 expression levels between irradiated and bystander cells at 4, 8, and 12 hours time points were significant (p=0.03). Significant difference in the expression of SOX2OT was also observed in irradiated TK6 and bystander cells (p=0.02) at 4, 12, and 24 hours time points. The SOX2OT expression was induced following radiation treatment of TK6 cells (Fig. 2D). The expression of SOX2OT returned to baseline level at 8 hours post irradiation, followed by an increase in its expression level at 12 hours time point that continued until 24 hours after radiation exposure (Fig. 2D). SOX2OT was repressed in bystander cells and its lowest levels were seen at 4 hours time point. Finally, we monitored the expression pattern of RMST in irradiated TK6 and bystander cells. RMST was initially induced in both irradiated and bystander cells (Fig. 2E). While its expression level continued to increase until 8 hours in irradiated cells, the RMST expression level was decreased at the 8 hours time point in bystander cells (Fig. 2E).
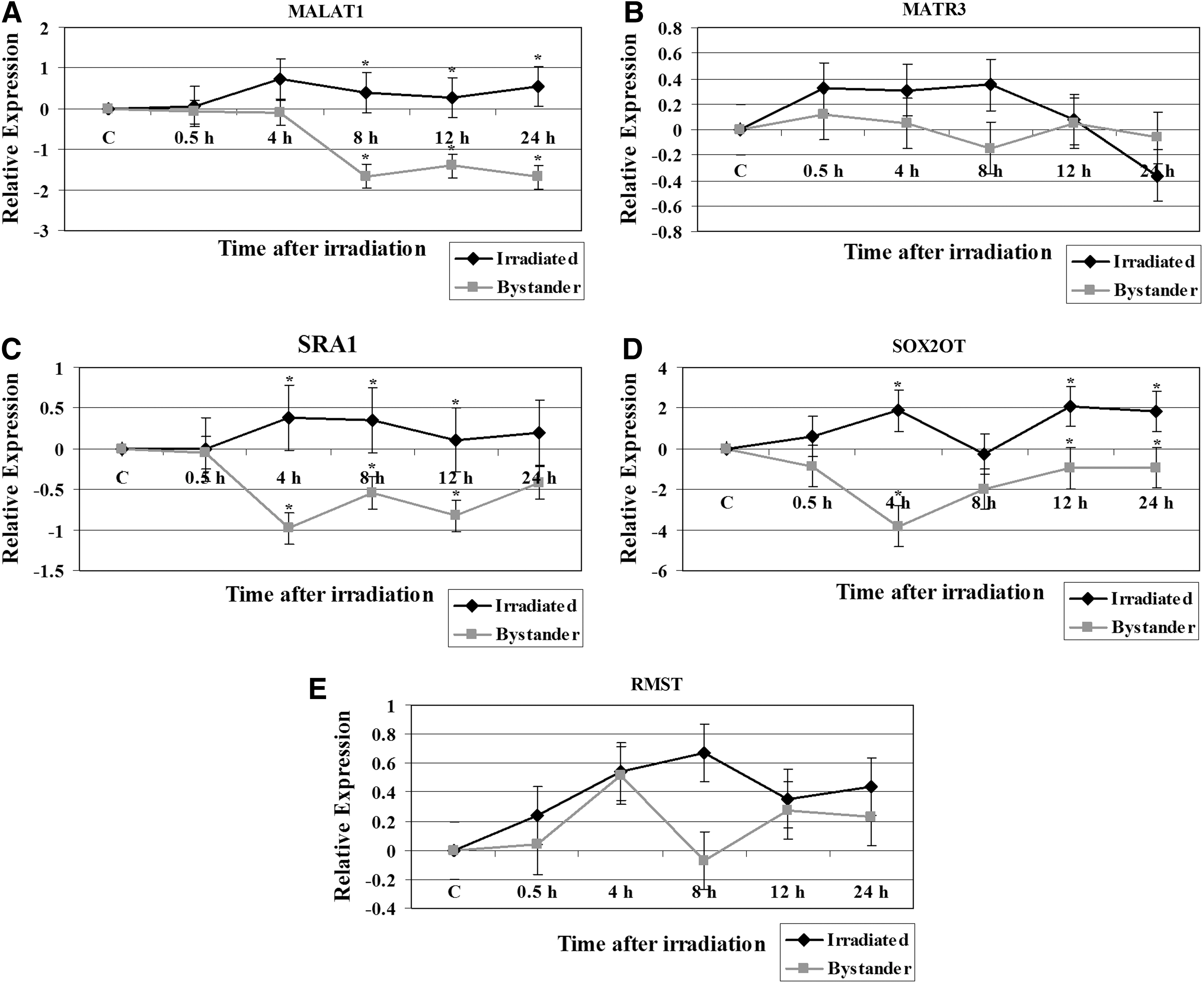
Expression analysis of non-coding RNA genes in 2 Gy irradiated TK6 and bystander cells. The data is plotted as Log2 values and indicate relative expression measured at 0, 4, 8, 12, and 24 hours time points after radiation treatment.
Discussion
Modern radiobiology documents the phenomenon of bystander effect, whereby cells that are not traversed by IR exhibit responses when in proximity to irradiated cells. The regulatory mechanisms involved in the bystander response have not been completely elucidated. To provide insight into the pathways responding in bystanders, we set out to measure ncRNA expression levels. In the present work we used 2 Gy X-ray irradiated normal human lymphoblasts TK6 cells in a transwell cell coculture system that precludes contact between targeted and bystander cells as a model system to examine ncRNA expression alterations. The bystander effect is known to occur in TK6 cells treated with 2 Gy dose of X-rays. 4 We made comparisons between directly irradiated and bystander ncRNA expression patterns after this single treatment. We show that alterations in the expression of ncRNA are involved in the radiation-induced medium-mediated bystander effect.
A number of ncRNA have been discovered, including miRNA, snoRNA, and lncRNA. We have previously reported the dysregulation of miRNA by radiation. 14 –18,26 Apart from miRNAs, the regulation of expression of other types of ncRNAs in bystander cells is unknown. Thus, we wondered whether these other ncRNAs were undergoing changes in bystander cells.
Expression alterations of snoRNA host genes in X-rays-treated and bystander cells
snoRNAs guide the modification of specific nucleotides in ribosomal RNAs (rRNAs) and small nuclear RNAs. The transcripts of these genes possess some characteristic features of an mRNA (splicing, polyadenylation, and cytoplasmic localization) but lack long open reading frames. 19 A number of tissue-specific snoRNAs have been identified that apparently do not target conventional substrates and are presumed to guide processing of primary transcripts of protein-coding genes. 27 We examined the modulation of several snoRNA host genes in TK6 and bystander cells exposed to 2 Gy of X-rays. The SNHG1 and SNHG4 were upregulated in irradiated TK6 cells but were repressed in cocultured bystander cells (Fig. 1). The expression of SNHG5 and SNHG11 was repressed in both irradiated and bystander cells. The SNHG6, SNHG7, and SNHG8 exhibited similar expression pattern in irradiated and bystander cells. Decreased snoRNA expression reduces the snoRNA-guided methylation of the target nucleotides and impaired rRNA modification lead to morphological defects. 28 Defects in ribosome biogenesis can have detrimental effects on cellular metabolism and vitality. Interestingly, a number of diseases have been associated with defects in ribosome synthesis pathways. 20 We previously reported that exposure of human cells to ionizing radiation results in the modulation of ribosomal genes. 29 We identified 31 ribosomal genes with altered expression in cells irradiated in G1 or G2 cell cycle phases. The snoRNAs have malfunctioning roles in tumorigenesis. The snoRNAs are overexpressed in lung tumors and act as oncogenes. 30 snoRNA expression patterns are negatively altered in leukemic cells and prompt cell growth through cell cycle modulation. snoRNA expression was implicated in the G0/G1 to S phase transition mediated by the Rb/p16 pathways. 31
Expression changes of long ncRNA in irradiated and bystander cells
Transcriptomic analyses have identified tens of thousands of intergenic, intronic, and cis-antisense lncRNA. Emerging evidence implicates the functional involvement of lncRNA in the regulation of gene expression, maintenance of subnuclear structures, transport of specific mRNAs, and control of pre-mRNA splicing. 32 The lncRNAs can regulate gene expression at epigenetic, transcription, and post-transcription levels and take part in cell development, immunity, oncogenesis, and clinical disease processes. 33 lncRNAs are dysregulated in a number of human diseases, including several cancers and neurological disorders and show tissue-specific expression. 34 The importance of the regulatory role played by ncRNAs in cancer-associated pathways such as cell growth, invasion, and metastasis has been documented. 35 Several lncRNA have increased expression in a number of cancer cells. 36 Genotoxic stress-inducible nuclear lncRNA have been identified. 37 The cigarette smoke exposure and aberrant expression and function of lncRNA have been linked. 38 To our knowledge, the modulation in the expression of lncRNA in irradiated and bystander cells has not been investigated.
MALAT1 is an lncRNA that is highly expressed in several tumor types and its expression levels are associated with tumor-promoting functions. 39 The function of MALAT1 is unknown in cells treated with ionizing radiation. While MALAT1 was induced in irradiated TK6 cells, its expression level was repressed in bystander cells (Fig. 2A). Other studies have shown that MALAT1 was downregulated in bleomycin-treated cells. 40 MALAT1 interacts with pre-mRNA splicing factors and regulates cancer cell migration, synapse formation, cell cycle progression, and responses to serum stimulation. It is proposed that MALAT1 function becomes apparent under particular conditions. 41,42 The upregulation of MALAT1 after radiation exposure suggests its function in DNA damage response.
The SRA1 gene encodes both ncRNAs and protein-coding isoforms, and they represent alternatively spliced transcript variants. The non-coding SRA1 is a coactivator for several nuclear receptors and is associated with breast cancer. 43 The lncRNA transcripts interact with a variety of proteins, including the protein encoded by this gene. The encoded protein acts as a transcriptional repressor by binding to the ncRNA. SRA1 RNA levels affect various biological functions, such as proliferation, apoptosis, steroidogenesis, myogenesis, glucose uptake, cellular signaling, T(3) hormone generation, and invasion/metastasis. 44 There was significant difference in the SRA1 expression levels between irradiated and bystander cells. The SRA1 expression was induced in X-rays-treated TK6 cells but was downregulated in bystander cells (Fig. 2C).
The lncRNA SOX2OT is embedded within an intron of SOX2 gene. SOX2OT expression has been analyzed in several developmental systems. 45 The involvement of SOX2OT in cells recovering from ionizing radiation-induced DNA damage is unknown. Significant difference in the expression of SOX2OT was observed in irradiated TK6 and bystander cells. The SOX2OT expression was induced following radiation treatment of TK6 cells (Fig. 2D). SOX2OT was repressed in bystander cells. These observations suggest the involvement of SOX2OT in ionizing radiation-induced stress pathways.
Our data show that the expression alterations of various ncRNA differ between directly irradiated and bystander cells. On the other hand, we also noticed that expression patterns of some ncRNA showed similarities among irradiated and bystander cells. Other studies have reported different gene expression profiles in irradiated and bystander cells. Global gene expression analyses of bystander and irradiated cells showed that CDKN1A was expressed at higher levels in the directly exposed cells but no change in its expression levels was observed in bystander cells. 46 Genes related to the cell cycle or death (CDKN1A, GADD45A, NOTCH1, and BCL2L1), and cell communication (IL1B, TCF7, and ID1) were upregulated in irradiated cells, but not in bystander cells. 47 We previously reported that the radiation-induced gene expression profile in irradiated cells was different from unirradiated bystander cells. 7 The ncRNA expression data suggest that some of the pathways leading to biological effects in the bystander cells could be different from the directly irradiated cells.
Similar expression patterns observed for certain ncRNA in directly irradiated and bystander cells indicate common functions in the ionizing radiation induced pathways. Many studies have attempted to compare the gene expression in directly irradiated and bystander cells. NFkappaB regulated genes PTGS2 (cyclooxygenase-2), IL8, and BCL2A1, responded similarly in bystander and irradiated cells. 46 No differences in the transcriptomes of cells grown in medium from X-irradiated cells or directly irradiated were seen. 48 The transcript profiles of cells grown in culture medium from irradiated cells and irradiated cells concluded that X-irradiation and bystander effects induce similar changes of transcript profiles in most functional pathways. 49
Cells exposed to ionizing radiation release factors that induce DNA damage, chromosomal instability, apoptosis, alterations in gene expression, and changes in the proliferation rate of neighboring unexposed cells. Our findings suggest that ncRNA levels are disturbed in bystander cells and may have a role in regulating the response of these cells leading to the induction of non-targeted cellular effects. The alteration in the balance of signaling is likely to lead to different outcomes in irradiated cells and their bystanders. These results imply that intercellular signaling between irradiated and bystander cells activate intracellular signaling, leading to the ncRNA transcriptional stress response in bystander cells.
Footnotes
Acknowledgments
The author wishes to thank Romaica Omaruddin for technical assistance and Dr. Howard Liber, Colorado State University, Fort Collins, CO, for providing TK6 cells. The author is thankful to the DNA analysis facility, University of Vermont for assistance with the real-time PCR experiments. This work was supported by an endowment fund, College of Nursing and Health Sciences, University of Vermont.
Disclosure Statement
There are no existing financial conflicts.