
Abstract
Purpose:
To monitor cell death in tumors during the rejection process after treatment with an antibody radiolabeled with a β-emitter.
Methods:
Tumors during rejection after treatment with 177Lu-labeled antibody BR96 and after administration of unlabeled BR96 were compared with untreated tumors from the same immunocompetent syngeneic rat tumor model. Cell death was monitored with the terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay and immunohistochemical staining of activated caspase-3 and γH2AX. These data were evaluated together with histopathological morphology, BR96-binding antigen expression, and 177Lu radioactivity distribution imaged by digital autoradiography.
Results:
The untreated tumors showed staining for all the markers, mainly in and around the necrotic areas. One to 2 days p.i. large areas were stained with anti-γH2AX, followed by a slight decrease. Staining of activated caspase-3 was intense and extensive 1–2 days p.i., while found in and around necrotic areas 3–8 days p.i. TUNEL staining was similar to activated caspase-3 staining 1–2 days p.i. but more extensive than activated caspase-3 staining 3–4 days p.i. Digital autoradiography revealed activity concentration in granulation tissue from 1 day p.i.
Conclusion:
Following radioimmunotherapy in an immunocompetent syngeneic colon carcinoma model, tumor cells did not only die through caspase-3-dependent apoptosis, but also by other mechanisms.
Introduction
Colorectal cancer is one of the most common carcinomas in humans. 1 Metastases are common, 2 necessitating development of systemic treatment strategies such as radioimmunotherapy. The limited efficacy of radioimmunotherapy against solid tumors hitherto can be partly attributed to the difficulty in achieving high tumor uptake and sufficient penetration of radiolabeled monoclonal antibodies. 3,4
Radiation is traditionally believed to induce cell death by apoptosis or mitotic catastrophe, 5,6 but a number of other mechanisms have also been described. 7 –10 Cell death can be classified according to morphological appearance, enzymatic criteria, or functional and immunological characteristics, and should not be based on one parameter alone, but by a combination of characteristics. 11 Most publications on the mechanisms of cell death describe studies performed in vitro. However, it is important to evaluate cell death pathways in vivo since tumors consist not just of tumor cells, and constitute an interactive, dynamic system.
Subcutaneous xenografts in mice are artificial tumor models that have little interaction with the host, and limited stromal and vascular interactions between human and mouse components, and will, therefore, have a low probability of metastasizing. 12 To better mimic the clinical situation, our group has developed a syngeneic immunocompetent rat tumor model of colon carcinoma for studies of radioimmunotherapy using the chimeric monoclonal antibody BR96, which targets the tumor-associated antigen Lewis Y (Ley). 13 In a previous study, we demonstrated that treatment with 400 MBq 177Lu-DOTA-BR96/kg body weight resulted in complete response (CR) in 17 of 19 tumor-bearing animals. 14 We have also previously shown that the activity distribution was heterogeneous when 50 MBq/kg was given. 15
The aim of the present study was to investigate cell death markers in tumors during the rejection process induced by radioimmunotherapy, and to compare the findings with untreated tumors and tumors treated with an unlabeled antibody. The cell death markers analyzed were terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay; γH2AX; and activated caspase-3. Also, the histopathology and the target antigen distribution were evaluated. The TUNEL assay stains fragmented DNA, which occurs late during all types of cell death. 16,17 γH2AX is a marker of double-strand breaks through foci formation. Activated (cleaved) caspase-3 is an executer caspase during apoptosis, which is active both in the extrinsic and intrinsic pathways. 16,17 The BR96-binding antigen makes it possible to determine whether the targeted antigen disappears from the tumor cells during rejection. The change in the intratumoral distribution of radiolabeled 177Lu-DOTA-BR96 in relation to viable tumor cells was also evaluated over time using digital autoradiography, as recently described by our group. 15
Materials and Methods
The monoclonal antibody
The chimeric (mouse/human) monoclonal IgG1 antibody BR96 (Seattle Genetics, Inc.), which binds to the Ley antigen, was employed. Ley is a tumor-associated antigen that is expressed on the majority of human epithelial tumors. Normal human tissue contains the BR96-binding antigen primarily in the epithelial cells of the gastrointestinal tract. 18 The binding affinity between BR96 and the cell line used in this study is strong; the dissociation constant being 4 nM. 13
The radioimmunoconjugate
Conjugation was performed according to Forrer et al. 19 Briefly, BR96 was transferred to 0.2 M sodium carbonate buffer, pH 9.5, by repeated centrifugation using an Amicon-15 filter (MWCO 30,000; Millipore). The DOTA-chelate (S-2-(4-isothiocyanatobenzyl)-1,4,7,10-tetraazacyclododecane tetraacetic acid (Macrocyclics; 2 mg/mL H2O) was added to the BR96 antibody (100 mg/mL) at a molar ratio of 3:1 (DOTA:IgG) and incubated for 1 hour at 37°C. The conjugate was purified by repeated centrifugation, using an Amicon-15 filter, transferred to 0.25 M ammonium acetate buffer, pH 5.3, and the final concentration was adjusted to 10 mg/mL BR96. All empty vials were pretreated with 1% HNO3 and all buffers were pretreated with Chelex-100 (BioRad) to remove metals.
Matrix-assisted laser desorption ionization mass spectroscopy (MALDI-MS) was used to determine the number of DOTA moieties per BR96 molecule, by desalting the sample to 18 MΩ·cm H2O using a centrifugation filter device, and dividing the increase in molecular mass by 688 (the molecular mass of the DOTA-chelate).
The immunoreactivity (i.e., the antigen-binding properties) of DOTA-BR96 relative to BR96 was determined by saturation binding curve analysis, using the tumor cells as target antigen. Briefly, increasing concentrations of BR96 and DOTA-BR96 (40 ng/mL–40 μg/mL) were added to the cell plate in triplicate and incubated for at least 90 minutes. The bound BR96/DOTA-BR96 conjugates were detected using rabbit HRP-anti-human-IgG (Dako), and the equilibrium binding constant (Kd) was calculated using Prism 5.02 software (GraphPad Software, Inc.; binding saturation–one site total, nonspecific binding and background constrained to a constant value of zero). The immunoreactivity is given by the ratio of the binding constants: Kd(BR96)/Kd(DOTA-BR96).
Both the DOTA-BR96 conjugate in 0.25 M ammonium acetate buffer and the 177LuCl3 solution (MDS Nordion) were preheated to 45°C for 10 minutes. The DOTA-BR96 solution was then added to the vial containing the radionuclide and incubated at 45°C for 15 minutes. The reaction was quenched with excess of DTPA (diethylene triamine pentaacetic acid) for 5 minutes. The radiolabeled immunoconjugate was diluted in 1% human serum albumin (HSA; Baxter Medical AB) to prevent radiolysis from affecting the immunoreactivity. The radiochemical purity was determined by instant thin-layer chromatography (ITLC), using a 1×9 cm silica-gel-impregnated fiberglass sheet as solid phase and 0.1 M EDTA as mobile phase. To confirm the radiochemical purity and to detect signs of aggregation or fragmentation, separation was performed using size-exclusion chromatography and high-performance liquid chromatography (HPLC) (using a 7.8×300 mm molecular sieving column, Phenomenex SEC S3000; Phenomenex, eluted in 0.05 M sodium phosphate at 1.0 mL/min).
Syngeneic animal model
BN7005-H1D2 is a cell line established from a 1,2-dimethylhydrazine-induced rat colon carcinoma in a Brown Norway (BN) rat. We have determined the radiosensitivity for this cell line, expressed as the fraction of survival after 2 Gy (S2Gy) to 0.5 (137Cs radiation source, unpublished data). This is similar to the radiosensitivity of human colorectal carcinoma cell lines. 20 The cells were cultured in RPMI-1640 medium supplemented with 10% fetal calf serum (both from PAA Laboratories GmbH), 1 mM sodium pyruvate, 10 mM HEPES buffer, and 14 mg/L Gentamicin (all from Gibco, Invitrogen) at 37°C, in a humidified environment containing 5% CO2. The cells were washed in phosphate-buffered saline and detached by treatment with trypsin (both from PAA Laboratories GmbH).
BN rats are immunocompetent and express the BR96-binding antigen in normal tissues, mainly in the epithelium of the gastrointestinal tract, 21 similar to humans. The animals were inoculated with 3×105 cells between the peritoneum and the abdominal wall under anesthesia (isoflurane; Abbott Scandinavia AB). All experiments were conducted in compliance with Swedish legislation on animal welfare, and these were approved by the Regional Animal Ethics Committee. The animals were housed under standard conditions and fed with standard pellets and fresh water ad libitum.
Radioimmunotherapy with 177Lu-DOTA-BR96
Thirty male BN rats (Harlan) were included in the study. Their median weight on the day of administration of radioimmunotherapy (day 0) was 238 g (interquartile range 8.5 g). Twenty-one animals were treated with 400 MBq/kg body weight of 177Lu-DOTA-BR96 (150 μg DOTA-BR96 in 0.4 mL saline with 1% HSA) by intravenous injection 13–14 days after cell inoculation. Our previous studies using the same model have shown that administration of this activity resulted in CR in 17 of 19 animals, 14 and that the activity is below the maximum tolerable dose of 600 MBq/kg. 22 The remaining 9 animals were left untreated as a control group. Tumor volumes were calculated as [tumor length×tumor width2×0.4]. 23 The median tumor volume on day 0 was 1140 mm3 (interquartile range 683 mm3). The treated animals were euthanized and dissected 1, 2, 3, 4, 6, and 8 days p.i. in groups of 3 animals per day (6 animals, 8 days p.i.), while the untreated animals were all euthanized and the tumors excised on day 0. The tumors were cut into two halves; half was snap-frozen and the other half was fixed in 4% paraformaldehyde and embedded in paraffin.
Treatment with unlabeled BR96 antibody
To evaluate the mechanisms of cell death resulting from the unlabeled antibody, 12 animals with tumors were treated with the unlabeled chimeric BR96 monoclonal antibody. Six animals were given 0.1 mg BR96/kg body weight and 6 animals were given 1.0 mg/kg. The animals were euthanized and dissected 1 and 2 days p.i. in groups of 3 animals per day. The tumors were cut in half and treated as described above.
Analysis and evaluation of biomarkers
The paraffin-embedded tumor samples were sectioned and consecutive sections were mounted on slides and analyzed for fragmented DNA (TUNEL assay, TACS® 2 TdT-Blue Label, and In Situ Apoptosis Detection Kit; Trevigen), activated caspase-3 (polyclonal rabbit antibody, Pierce Biotechnology, Rockford, IL), γH2AX (monoclonal rabbit antibody; Epitomics, Inc.), and BR96-binding antigen.
The TUNEL assay was conducted according to the manufacturer's instructions. Briefly, after storage at 4°C the 10 μm sections were incubated at 60°C, rehydrated, and treated with Proteinase K for 15 minutes at 37°C. Sections were incubated with Labeling Buffer for 5 minutes at room temperature, then with Labeling Reaction Mix for 1 hour at 37°C, and finally, with Stop Buffer for 5 minutes at room temperature. After washing, the sections were incubated with Strep-HRP Solution for 10 minutes at 37°C, and after additional washing, with TACS-Blue Label for 3 minutes at room temperature. Finally, the sections were counterstained with Nuclear Fast Red for 1 minute and dehydrated before mounting.
For immunohistochemical staining, 4 μm sections were incubated at 60°C prior to storage at 4°C. Sections were rehydrated and antigen retrieval was performed using the PT Link pretreatment module (Dako) with acidic target retrieval solution (pH 6), preheated to 65°C. The solution containing the sections was then heated to 99°C for 20 minutes, after which the slides were rinsed with distilled water. The sections were incubated overnight at room temperature with BR96, anti-activated caspase-3, or anti-γH2AX diluted in Antibody Diluent (Dako). After washing with Tris-buffered saline containing Tween 20, the sections were incubated with anti-human IgG H+L Donkey F(ab)2 HRP (Jackson ImmunoResearch Laboratories, Inc.) (for BR96-binding analysis) or Donkey anti-rabbit HRP (Jackson Laboratories) (for anti-activated caspase-3 and anti-γH2AX analyses) in Antibody Diluent for 3 hours (BR96) or 1 hour (anti-caspase-3 and anti-γH2AX) at room temperature. Finally, 3-amino-9-ethylcarbazole (AEC; Dako) was added before counterstaining with hematoxylin and mounting of the slides.
Consecutive tumor sections with three cell death markers (TUNEL, anti-caspase-3, and anti-γH2AX) were evaluated blindly by an experienced clinical pathologist to identify, quantify, and localize stained cells from the tumors treated with radioimmunotherapy and the untreated tumors.
Evaluation of histopathology
Consecutive tumor sections to those used for biomarker staining were stained with hematoxylin and eosin (H&E). The H&E-stained sections were used to evaluate the histopathological morphology, and to quantitatively score the amount of viable tumor cells, necrotic cells, granulation tissue, fibrous tissue, and infiltration of granulocytes and lymphocytes, on a scale from 0 to 5. A score of 0 corresponded to 0% of the total area, 1: 1%–20%, 2: 21%–40%, 3: 41%–60%, 4: 61%–80%, and 5: 81%–100%. The scores for granulocytes and lymphocytes corresponded to the percentage of all infiltrating cells rather than that of the total area. The samples from tumors treated with radioimmunotherapy and untreated tumors were evaluated by the same pathologist, who carried out the evaluation thrice, with intervals of at least 1 month between evaluations.
Digital autoradiography
The 177Lu activity distribution in frozen tumors was imaged, using a double-sided silicon strip detector with an intrinsic spatial resolution of 50 μm (Biomolex 700 Imager; Biomolex AS), as described previously. 15 Microscope slides containing three frozen tumor sections (thickness 10 μm) were analyzed for a minimum of 40 minutes each. Corrections were applied for dead or miscalibrated strips, using IDL 6.4 software (ITT Visual Information Solutions). After imaging, the sections were H&E stained, to evaluate the activity distribution in relation to tumor cells, granulation, and fibrous tissue.
The mass per pixel was determined by dividing the mass of the sections on one slide by the area of the sections in the activity image. The mass was calculated by multiplying the thickness of the section by the area of the section obtained from histological staining, and then multiplying this by an assumed density of 1.0 g/cm3. Finally, all images were scaled and corrected for decay to give the activity per gram of tissue at the moment of sacrifice.
Results
The radioimmunoconjugate
The average number of DOTA moieties conjugated per BR96 molecule was 2.4. The immunoreactivity was 0.9, which is not statistically different from unlabeled BR96, indicating that the antigen-binding properties did not change as a result of conjugation with DOTA. The radiochemical purity of 177Lu-DOTA-BR96 was analyzed using ITLC and found to be 97%. Less than 1% of labeled aggregated antibody was detected by HPLC.
Sampling during radioimmunotherapy
All 30 animals developed tumors before day 0. The 9 untreated controls were euthanized and the tumors excised on day 0. The animals treated with radioimmunotherapy were euthanized and the tumors excised at 1, 2, 3, 4, 6, and 8 days p.i. in groups of 3 animals per day, apart from day 8 p.i., when 6 animals were euthanized and dissected. Tumor tissues could not be sampled in 5 of the animals, due to CR on the day of sacrifice and sampling: day 4 (1 CR), day 6 (1 CR), and day 8 (3 CR) p.i. (Table 1).
RIT, radioimmunotherapy; n, number of animals; CR, complete response; TUNEL, terminal deoxynucleotidyl transferase dUTP nick end labeling.
Sampling during antibody therapy
Twelve animals with tumors were treated with the unlabeled chimeric BR96 monoclonal antibody. Half of the animals were given 0.1 mg BR96/kg body weight and the other half was given 1.0 mg/kg. The animals were euthanized and dissected 1 and 2 days p.i. in groups of 3 animals. None of these animals had CR.
Cell death markers
In the untreated tumors, all three cell death markers activated caspase-3, γH2AX, and the fragmented DNA (TUNEL assay), and showed positive staining in and around the necrotic areas, as shown in Table 1 and Figures 1 and 2. In tumors from animals treated with radioimmunotherapy, staining for activated caspase-3 showed disseminated positively stained cells in the vicinity of necrotic areas and in areas of viable tumor cells early in the rejection process (1–2 days p.i.). At 3–8 days p.i., activated caspase-3 staining was mostly seen in and around necrotic areas, similar to untreated tumors. TUNEL-positive cells were more widely dispersed during both the early and intermediate (1–4 days p.i.) phases of the rejection process. Later (6–8 days p.i.) TUNEL-positive staining was seen only in and around necrotic areas as observed in the untreated tumors. At 8 days p.i. one of three tumors displayed TUNEL-positive cells as seen day 1–4 days p.i. Massive staining with anti-γH2AX was seen in the early phase of rejection (1–2 days p.i.), which decreased slightly during the intermediate and late (3–8 days p.i.) phases, but remained elevated compared with the untreated tumors during the whole study period. γH2AX-positive cells were not detected in areas with tumor cells in the intermediate or late phases of the rejection process. This is in contrast to the staining pattern of activated caspase-3 and TUNEL-positive cells, which showed localization mainly in tumor cell-rich areas for all points in time. Analysis of the binding to the BR96-target antigen showed a decrease in the fraction of cells binding BR96 throughout the study period, as the fraction of tumor cells in the tumor decreased. However, all the detected tumor cells were BR96-positive.
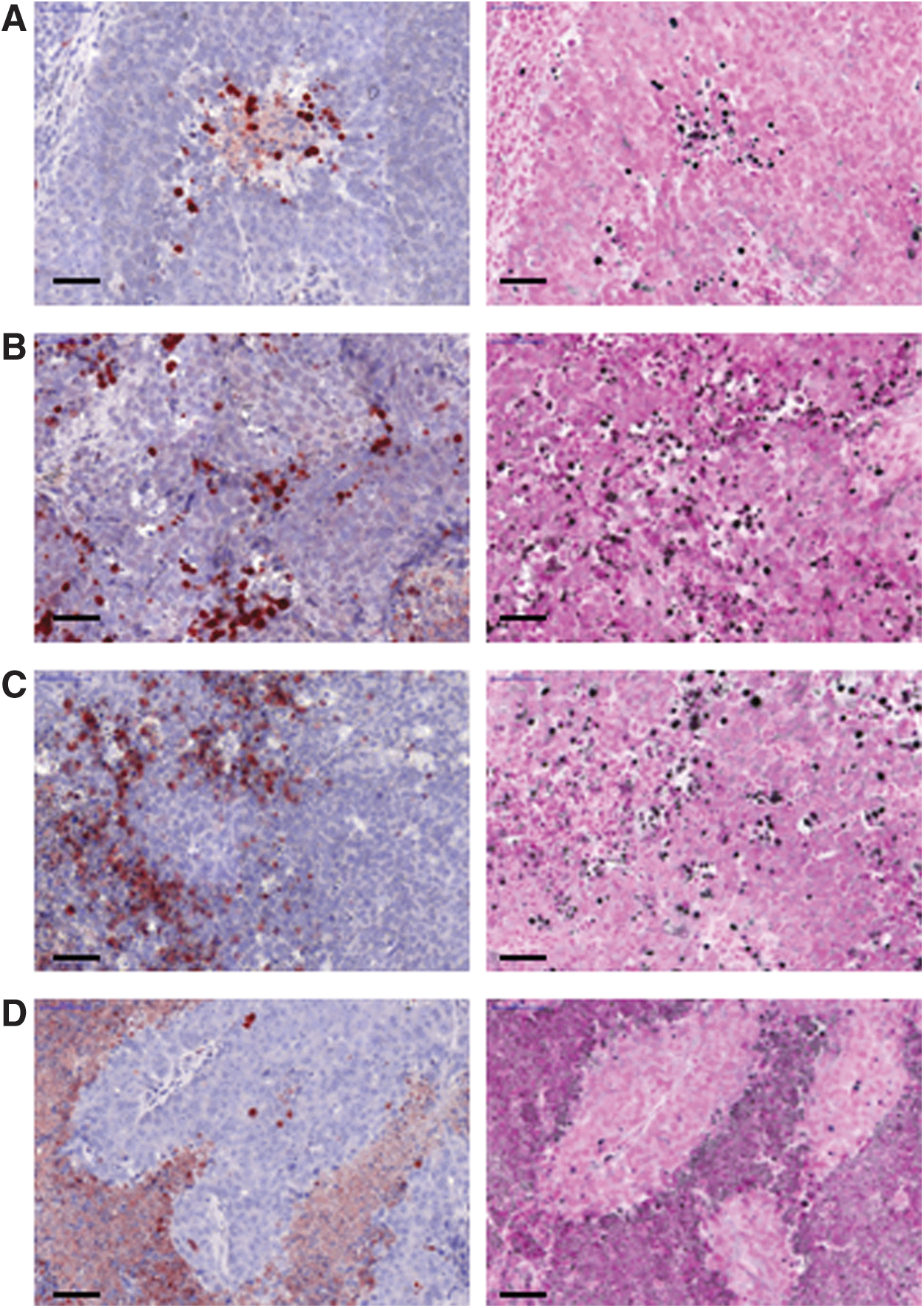
Left: Tumors stained with anti-activated caspase-3: the nuclei in positive cells are stained red. Right: Tumors stained with TUNEL assay: nuclei in the positive cells are stained black.

Sections of tumors stained with anti-γH2AX: positive cell nuclei are stained red.
The tumors from animals treated with the unlabeled antibody showed no increase in γH2AX or activated caspase-3 compared with the untreated tumors, as seen in Table 1. The fraction of TUNEL-positive cells increased in tumors treated with 1.0 mg/kg both 1 and 2 days p.i., compared with the untreated tumors. The TUNEL-positive cells were mainly found in the edges of areas containing tumor cells. At the lower dose of BR96 (0.1 mg/kg), the results were comparable to those in untreated tumors.
Histopathological evaluation
All tumors were individually and quantitatively scored from 0 to 5, for each of the following six histopathological characteristics: viable tumor cells, necrotic cells, granulation tissue, fibrous tissue, infiltration of granulocytes, and infiltration of lymphocytes. Typical examples of the histopathological characteristics are given in Figure 3, and the average score for the variables analyzed is given in Table 2. The intraobserver variation obtained from evaluating the scores three times at intervals of at least 1 month was at most one score, apart from 3 out of 125 cases. The fraction of viable tumor cells decreased continuously from day 1 after radioimmunotherapy, resulting in the first CR occurring 4 days p.i. The fraction of necrotic tissue increased initially, showing a maximum on 2 days p.i., and then returned to about the same level as seen in untreated tumors. Granulation tissue also increased initially, showing a maximum on 4 days p.i., before declining to a value similar to that seen in untreated tumors. The fraction of fibrous tissue started to increase a few days after treatment and increased throughout the entire study period. Untreated tumors did not demonstrate any fibrous tissue. The fraction of infiltrating granulocytes initially showed a slight increase, but the individual variation increased with time. Lymphocyte infiltration decreased slightly during the early phase of rejection (1–4 days p.i.), but returned to the same levels as in untreated tumors at the end of the study. Representative examples of the histological characteristics of tumor sections are shown in Figure 4.

Pictures illustrating the histological characteristics evaluated
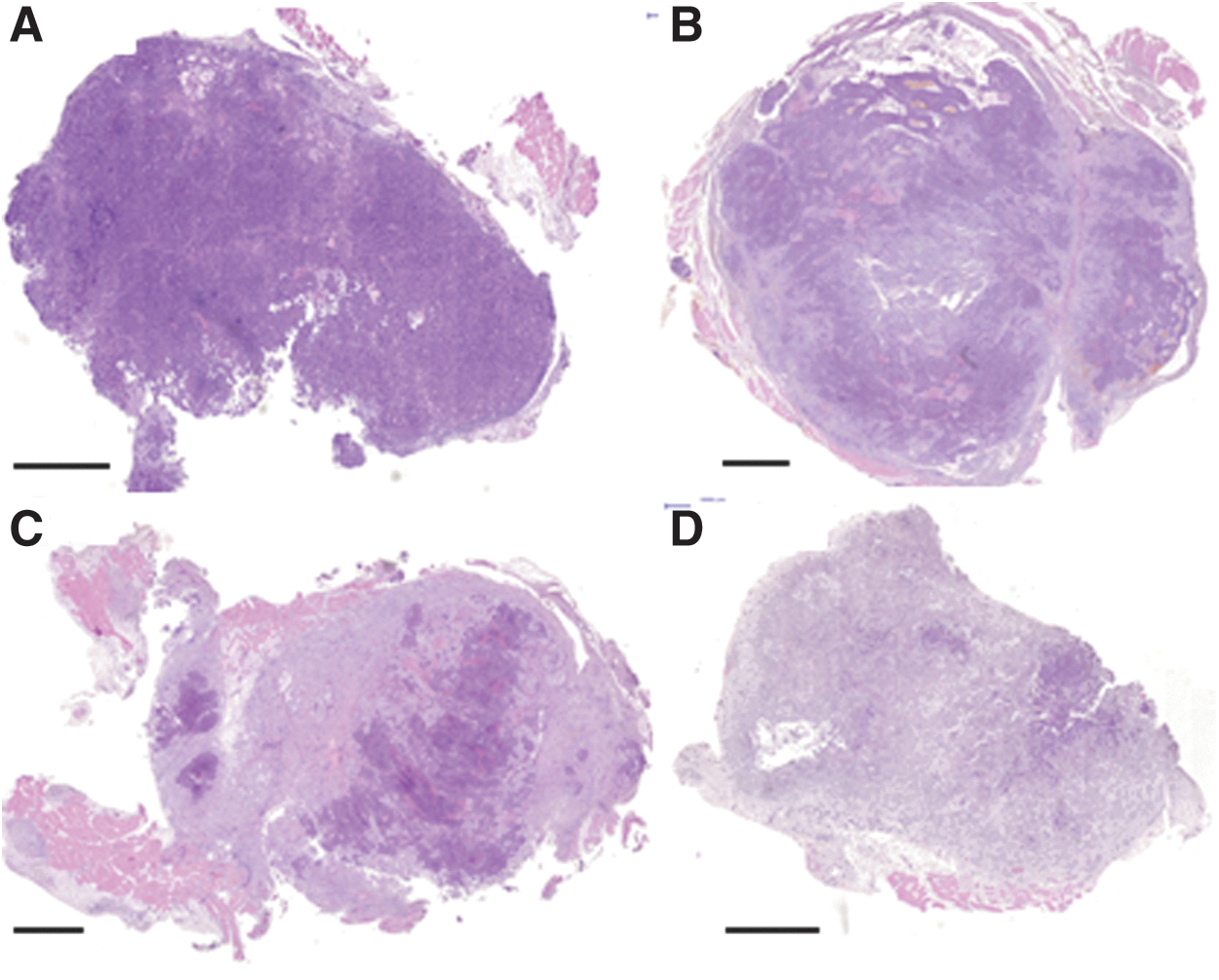
H&E-stained tumors during the rejection process following radioimmunotherapy; the dark purple areas illustrate viable tumor areas while the lighter areas correspond to granulation/fibrous tissue
All tumors were individually scored for each variable from 0 to 5. A score of 0 corresponds to 0% of the total area, 1: 1%–20%, 2: 21%–40%, 3: 41%–60%, 4: 61%–80%, and 5: 81%–100%. The score for granulocytes and lymphocytes corresponds to the percent of all infiltrating cells rather than the total area. The average scores for viable tumor cells, necrotic tissue, granulation tissue, fibrous tissue, lymphocytes, and granulocytes are based on the remaining tumors.
Digital autoradiography
Areas with high activity uptake relative to the whole tumor section generally exhibited a low tumor cell density and signs of granulation tissue, compared with areas with lower activity uptake, at 1 day p.i. Areas with high activity were generally correlated to a continuous increase in granulation and fibrous tissue 2–6 days p.i. A clear decrease in tumor cell density was seen when evaluating the entire tumor section. At the end of the study, 8 days p.i., two of three tumors showed a decrease in tumor cell density and volume (3 other animals displayed CR, and no tumor was detected). The third tumor initially showed a reduction in size and then stayed stable, and histopathological analysis revealed an activity uptake in areas of granulation tissue, while the tumor cell areas displayed low activity uptake. The two tumors that had decreased most in volume (99% and 76%) exhibited more homogeneous activity distribution (as shown in Fig. 5) than the tumor that had decreased least in volume (56%).

Top panels: activity distributions in sections of tumors 1, 4, and 8 days after the injection of 177Lu DOTA-BR96. Bottom panels: the same sections stained with H&E. The figures illustrate heterogeneous activity distribution, mainly located in tumor cells 1 day after injection, and mainly located in granulation tissue 4 days after injection. The activity distribution was more homogeneous 8 days after injection, and was distributed over the entire tumor section. Scale bars: 2 mm. Color images available online at
Overall, digital autoradiography revealed an initially heterogeneous distribution of the activity in all tumor sections, with a more homogeneous distribution later in the rejection process. The activity was mainly located in areas of granulation and fibrous tissue, at all points in time.
Discussion
Cells exposed to external beam radiation are generally reported to succumb by apoptosis or mitotic catastrophe. 6 However, the most important finding of this study was that the tumor cells not only died through caspase-3-dependent apoptosis after radioimmunotherapy, but also by other mechanisms, for example, caspase-3 independent apoptosis, necrosis, and/or necroptosis. This is illustrated by the more extensive distribution of TUNEL-positive cells than that of activated caspase-3-positive cells. The results of this study also indicate that cell death is not delayed, since the activated caspase-3 and fragmented DNA (TUNEL assay) levels had returned to their initial values 3–4 days p.i. (activated caspase-3) and 6–8 days p.i. (fragmented DNA). It is difficult to determine the energy deposited in individual tumor cells by radioimmunotherapy as the local uptake and kinetics due to redistribution within the tumor are not known. Our findings indicate that the mechanisms causing cell death as a result of radioimmunotherapy might differ from reports using external beam radiation. This can be partly explained by the effects of the antibody, as illustrated by the increase in TUNEL-positive cells, and the lack of increase in γH2AX and activated caspase-3, in tumors treated with unlabeled BR96.
Radioimmunotherapy combines the therapeutic effects of antibodies and radionuclides, as exemplified by studies with Bexxar (131I-labeled anti-CD20), in which cell death was induced by antibody-dependent cellular cytotoxicity (ADCC), receptor-mediated apoptosis, and the absorbed dose from the radioisotope. 24,25 In the present study, we have demonstrated caspase-3-independent cell death caused by unlabeled BR96, and both caspase-3-dependent and caspase-3-independent cell death due to 177Lu-BR96. Earlier studies have shown that administering monoclonal antibodies usually results in apoptosis. 24,26 The histological changes after administration of unlabeled antibodies described in this study, illustrating an increase in granulation tissue, immune cells, and a decrease in tumor cells, have been previously described by us. 15 The antibody treatment in this study results in caspase-3-independent cell death; others have shown that antibodies can induce receptor-mediated or immune-mediated tumor cell death, by phagocytosis, complement-dependent lysis, ADCC, or T-cell activation. 24,25,27
Few studies have addressed cell death mechanisms in vivo, despite the fact that such knowledge is important for the evaluation of the possible limitations of existing therapies and the design of new treatment modalities. In one study on the mechanisms of cell death after radioimmunotherapy in a p.i. xenograft model with the α-particle emitter 212Pb-trastuzumab, the induction of G2 cell-cycle arrest and delayed DNA damage repair were demonstrated, and the induction of apoptosis was illustrated, using the TUNEL assay 24 hours after exposure. 28 Apoptosis was not found to be caspase-3-dependent, and the inhibition of DNA damage repair was not completely reversed 96 hours after exposure.
Activated caspase-3 influence the macrophage activation and cytokine production after engulfment of the apoptotic cells, resulting in the recruitment of more macrophages and more efficient phagocytosis of the activated caspase-3-positive cells. 29 We detected a slight increase in the infiltration of granulocytes, at least early in the rejection process, indicating that cell death in this tumor model recruited neutrophils, which may lead to engulfment of dead tumor cells. The fact that the levels of both activated caspase-3 and TUNEL returned to baseline values indicate that tumor cell debris is transported away from the tumor. The initial decrease in lymphocyte infiltration might be a side effect of the initially higher dose rate, and the return of the amount of lymphocytes to baseline levels may indicate an immune response against the tumor cells. The lymphocytes present after radioimmunotherapy may be functionally different from the lymphocytes in untreated tumors, especially if radioimmunotherapy has induced immunogenic cell death. 30 –32 We have initiated studies on characterization of infiltrating inflammatory cells before and during radioimmunotherapy.
Staining of γH2AX concentrated as foci in the nucleus is considered a marker of DNA double-strand breaks. However, it has been reported that apoptotic cells may be γH2AX positive by pan-nuclear nuclei staining. 33 In this study, tumors treated with radioimmunotherapy displayed intense nuclear staining of almost the whole tumor section during the early phase of the rejection process, although the distributed radioactivity was heterogeneous at this point in time. The level of γH2AX was increased throughout the entire period of the study, compared with that of untreated tumors. 177Lu, which is a β-particle emitting radionuclide with a physical half-life of 6.65 days, is generally not regarded as a double-strand break inducer at therapeutic activities. 34 In the present study, γH2AX staining was mainly seen in granulation and fibrous tissue 3–4 days and 6–8 days after treatment, which could be explained by a corresponding distribution of 177Lu.
According to the results of in vitro studies, γH2AX foci are detectable minutes after γ-irradiation, reaching a peak after 30 minutes. 35 –38 After the plateau phase of 1 hour, the number of foci decreased, and returned to background levels within 24 hours. 37,39 We have evaluated γH2AX in vitro on the cell line used in this study (unpublished data) and detected γH2AX foci 1 hour after exposure to 2 Gy of γ-irradiation (using a 137Cs radiation source; Gamma Cell-3000 Elan; 8 Gy/min). The number of foci had decreased to background levels after 24 hours.
Others have shown that γH2AX can be used as a marker of inherent genomic instability, 40 indicating that results based on γH2AX should be interpreted with caution. In this study, there was a clear difference between γH2AX staining pattern in treated and untreated tumors, indicating that the results do not only depend on genomic instability. γH2AX should be regarded not only as a marker for double-strand breaks, but also as a general biomarker of agents that damage DNA. 41,42
Studies on cell cultures treated with 90Y-cetuximab have shown that the number of γH2AX foci was correlated to the amount of membranous target antigen of cetuximab, and that survival was directly correlated to the number of γH2AX foci after 48 hours. 43 The authors concluded that these cells do not succumb to delayed apoptosis due to mitotic catastrophe. Mitotic catastrophe has been detected in in vitro studies 2–6 days after external beam radiation, 6 compared with apoptosis that is observed ∼4 to 6 hours after exposure. 16 In our study, the therapeutic response was fast. Thus, a fast cell death was more plausible making mitotic catastrophe unlikely. It is, however, difficult to detect mitotic catastrophe on tissue sections since the 3D structure of the tumor makes the morphological evaluation of the nucleus more complicated compared with cell cultures.
Histopathological evaluation with H&E staining of tumors treated with radioimmunotherapy showed a reduction in the fraction of viable tumor cells in the remaining tumor throughout the period studied, which was macroscopically confirmed by the decrease in tumor volume. Necrotic areas represent histological changes resulting from cell death, for example, apoptosis or oncosis, 10 often depending on lack of oxygen and nutrition. The maximum area of necrotic tissue was observed 2 days p.i. The amount of granulation tissue was maximal 4 days p.i. Granulation tissue matures into fibrous tissue, which increased throughout the study period. Evaluating the numbers of granulocytes and lymphocytes is more difficult, as these cells are mainly found in the necrotic areas (granulocytes) and tumor margins (lymphocytes), and the differences between treated and untreated tumors were not as large as for the other characteristics.
Digital autoradiography showed activity in necrotic and granulation tissue 1 day p.i. In a previous study using the same tumor model, we found activity in the tumor margins 2–8 hours p.i., in viable antigen-expressing areas within the tumor 24 hours p.i., and increasing activity in antigen-negative areas of granulation tissue 48 hours p.i. and later. 15 The difference in observations 1 day after injection may be due to the higher activity of 177Lu-BR96 administered in the current study, which was 400 MBq/kg, compared with 25 and 50 MBq/kg in the previous study. The higher administered activity resulted in increased therapeutic effects, as verified by histopathology. Based on calculations presented in the previous study, 15 the mean absorbed dose rates over the whole tumor could be expected to be in the range of 0.2–0.6 Gy per hour, 24 hours p.i., with the activity used in the current study.
The results of the present study also showed that the activity per gram tumor increased with time, and that the distribution became more homogeneous later in the rejection process, probably due to the decrease in the tumor volume. Of the tumors not showing CR, the tumors excised 4 days p.i. (CR in 1 of 3 animals) had decreased by 75% and 94% in volume, whereas the tumors dissected 8 days p.i. (CR in 3 of 6 animals) had decreased by 56%, 76%, and 99% in volume. This highlights the need to incorporate changes in tumor volume in calculations of absorbed dose from a therapeutic activity. In the case of lymphoma, Hindorf et al. 44 have shown that the decrease in tumor volume can increase the calculated absorbed dose by up to 75%, illustrating that the decrease in tumor volume can lead to concentration of the activity.
The present study showed that some, but not all, of the tumor cells in this preclinical syngeneic rat colon carcinoma model succumbed due to caspase-3-dependent cell death. This illustrates that cell death in a syngeneic model can be caused by a mixture of cell death mechanisms when using 177Lu (β-particle emitting) radioimmunotherapy. Radioimmunotherapy can induce diverse cell death mechanisms as a result of the heterogeneous distribution of absorbed dose, but also the combined effects of the antibody and radiation as indicated in this study by the comparison between tumors treated with antibodies and radioimmunotherapy.
Conclusions
Analysis of cell death markers after radioimmunotherapy in an immunocompetent syngeneic rat colon carcinoma model revealed that tumor cells not only succumbed due to caspase-3-dependent apoptosis, but also by other mechanisms, as evidenced by the results of the TUNEL assay. This illustrates that fragmented DNA is distributed differentially compared with activated caspase-3. These other cell death mechanisms were not further evaluated in this study, but might be, for example, caspase-3-independent apoptosis, necrosis, and/or necroptosis. Cell death could be demonstrated already 1 day after the injection of 177Lu-BR96. Digital autoradiography revealed activity in fibrous and granulation tissue 1 day p.i. The scoring of histopathological variables showed a maximum area of necrotic tissue 2 days p.i. and of granulation tissue 4 days p.i.
Footnotes
Acknowledgments
We would like to thank Dr. Peter Senter (Seattle Genetics) for kindly providing the monoclonal antibody BR96. This research was supported by grants from the Swedish Cancer Society, Mrs. Berta Kamprad's Foundation, Gunnar Nilsson's Foundation, The Crafoord Foundation, Governmental Funding of Clinical Research within the National Health Service, King Gustaf V's Jubilee Foundation, The Lund University Medical Faculty Foundation, and The Lund University Hospital Fund, and the Eurostars program through the Swedish Governmental Agency for Innovation Systems (VINNOVA).
Disclosure Statement
The authors have no affiliations, memberships, funding, or financial holdings that might be perceived as affecting the objectivity of this publication.