
Abstract
Recent studies suggest that the use of cucurbitacins could inhibit cancer cell progression. In the current study, the authors analyzed the effect of cucurbitacin-E (CuE) in cancer cells using A549, Hep3B, and SW480 cells. The authors found that CuE inhibited cell proliferation and modulated the expression of cell cycle regulators in these cells. Moreover, the authors found that CuE inhibited Wnt/β-catenin signaling activation through upregulation of tumor suppressor Menin. Indeed, ablation of Menin using small interfering RNA (siRNA) oligos attenuated the antiproliferative roles of CuE. Taken together, the results of this study provide a novel mechanism that may contribute to the antineoplastic effects of CuE in cancer cells.
Introduction
Abnormal expression and (or) activity of many oncogenes and tumor suppressors, such as β-catenin, C-myc, PTEN, and P53, play critical roles in the development of human cancers. 1,2 Recent studies suggest that the anticancer ability of several natural products may be associated with regulation of these genes. 3,4 For instance, metformin, which is usually prescribed as an oral antihyperglycemic agent, could reduce risk and improve prognosis of certain cancers. 5,6 At the molecular level, metformin inhibits cancer cell proliferation and migration through repressing the insulin/insulin-like growth factor 1 (IGF1) signaling pathway and upregulation of p53 contents. 7,8
Cucurbitacins, which are widely distributed in plants, belong to a class of highly oxidized tetracyclic triterpenoids. 9 They have attracted much attention for their promising anticancer activities, including antiproliferation, cell cycle arrest, and induction of apoptosis. 10,11 It has been shown that cucurbitacin-B regulates immature myeloid cell differentiation and enhances antitumor immunity in patients with lung cancer. 12 Besides, cucurbitacin-B enhances the radiation sensitivity of human breast cancer cells via G2/M cell cycle arrest. 13 Moreover, cucurbitacin-D increases the proportion in the sub-G0/G1 phases and G2/M phases of the cell cycle, resulting in growth inhibition and apoptosis in human endometrial and ovarian cancer cells. 14
In the present study, the antiproliferative role and molecular mechanisms of cucurbitacin-E (CuE) were explored in A549, Hep3B, and SW480 cells. The results suggest that CuE could suppress cancer cells proliferation through attenuation of Wnt/β-catenin signaling.
Materials and Methods
Cell culture and reagent
A549, Hep3B, and SW480 cells were obtained from the Cell Bank of Type Culture Collection of Chinese Academy of Sciences (CAS, Shanghai, China). Cells were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum (Gibco, Shanghai, China). CuE was purchased from Sigma-Aldrich (St. Louis, MO) and dissolved in the DMSO solution.
BrdU assays
A cell proliferation enzyme-linked immunosorbent assay was used to analyze the incorporation of BrdU during DNA synthesis according to the manufacturer's protocols (Beyotime, Shanghai, China). Absorbance was measured at 450 nm in the Spectra Max 190 ELISA reader (Molecular Devices, Sunnyvale, CA). All experiments were performed in triplicate.
siRNA, RNA extraction, and real-time analysis
All small interfering RNA (siRNA) oligos were obtained from Dharmacon Company (Lafayette, CO). The siRNA designed for the knockdown of Menin had the following sequence: 5′-CAGCUUCAUCACAGGUUGGAG-3′. A scrambled sequence of 5′-AGUUCGAGGUGUACGAC UA-3′ was used as a negative control. Cells were seeded onto six-well plates and then transfected with 50 nM siRNA oligos. Total RNAs were isolated from cells by TRIzol reagent, and reverse transcriptions were performed by the Takara RNA PCR kit according to the manufacturer's instructions (Takara, Dalian, China). To quantify the transcripts of the interest genes, real-time PCR was performed using an SYBR Green Premix Ex Taq (Takara, Kyoto, Japan) on Light Cycler 480 (Roche, Basel, Switzerland).
Western blot
Cells were harvested and lysed with lysis buffer (50 mM Tris–HCl, pH 6.8, 100 mM 2-ME, 2% w/v sodium dodecyl sulfate [SDS], 10% glycerol). After centrifugation at 20,000 g for 10 minutes at 4°C, proteins in the supernatants were quantified and separated by 10%–12% SDS–polyacrylamide gel electrophoresis and then transferred to nitrocellulose (NC) membrane. After blocking with 10% nonfat milk in phosphate-buffered saline, membranes were immunoblotted with antibodies as indicated, followed by horseradish peroxidase-linked secondary antibodies. The signals were detected by the SuperSignal West Pico Chemiluminescent Substrate kit (Pierce, Rockford, IL) according to the manufacturer's instructions. Anti-Menin, APC, Axin, and β-catenin antibodies were purchased from Abcam Company (Burlingame, CA). Anti-cyclin D1 and cyclin E antibodies were purchased from Santa Cruz Company (Santa Cruz, CA). Protein levels were normalized to Lamin B (Millipore, Billerica, MA) or GAPDH (Abcam, Cambridge, MA).
Statistical analysis
Data are expressed as the mean±SEM (standard error of mean) from at least three separate experiments. Differences between the groups were analyzed using the Student's t-test. A p-value of <0.05 was considered statistically significant.
Results
The effects of CuE on cancer cell proliferation
To explore the effects of CuE on cell proliferation, three types of cancer cells were used in the current study. A549 cells are derived from lung cancer. Hep3B cells are from hepatocellular carcinoma. SW480 cells are from colon cancer. These cells were treated with CuE, and cell growth was examined by BrdU incorporation assays. As a result, CuE inhibited cell proliferation in a dose-dependent manner (Fig. 1A–C).

The effects of cucurbitacin-E (CuE) on cancer cell proliferation.
CuE inhibits Wnt/β-catenin signaling in cancer cells
To test whether the Wnt/β-catenin pathway mediates the anticancer action of CuE, abundance of nuclear β-catenin, the hallmark of Wnt signaling activation, was examined. As shown in Figure 2A–C, the protein contents of nuclear β-catenin were decreased in A549, Hep3B, and SW480 cells treated with CuE for 4 hours (Fig. 2A–C). Besides, cyclin D1 and cyclin E, two downstream target genes of β-catenin, were also downregulated by CuE treatment (Fig. 2D–F).
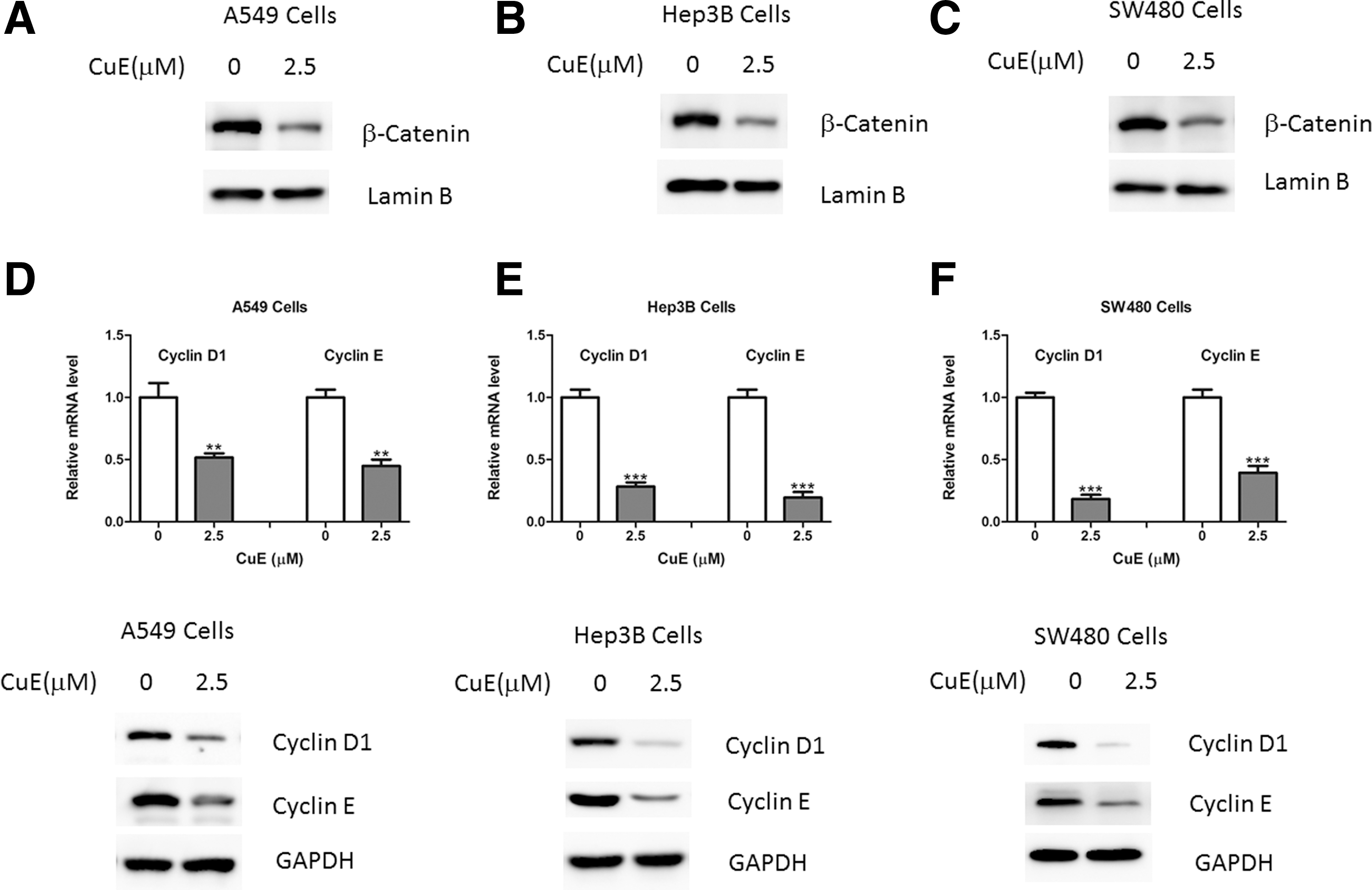
CuE inhibits Wnt/β-catenin signaling in cancer cells.
CuE upregulates Menin expression in cancer cells
It has been demonstrated that Wnt signaling is negatively regulated by several proteins, including adenomatous polyposis coli (APC), Axin, and Menin. 15,16 Here, the authors observed that Menin expression was markedly increased in A549 cells treated with CuE (Fig. 3A, B), whereas the expression levels of APC and Axin remained unaltered (Fig. 3A, B). Moreover, similar results were also observed in Hep3B and SW480 cells (Fig. 3C–F).
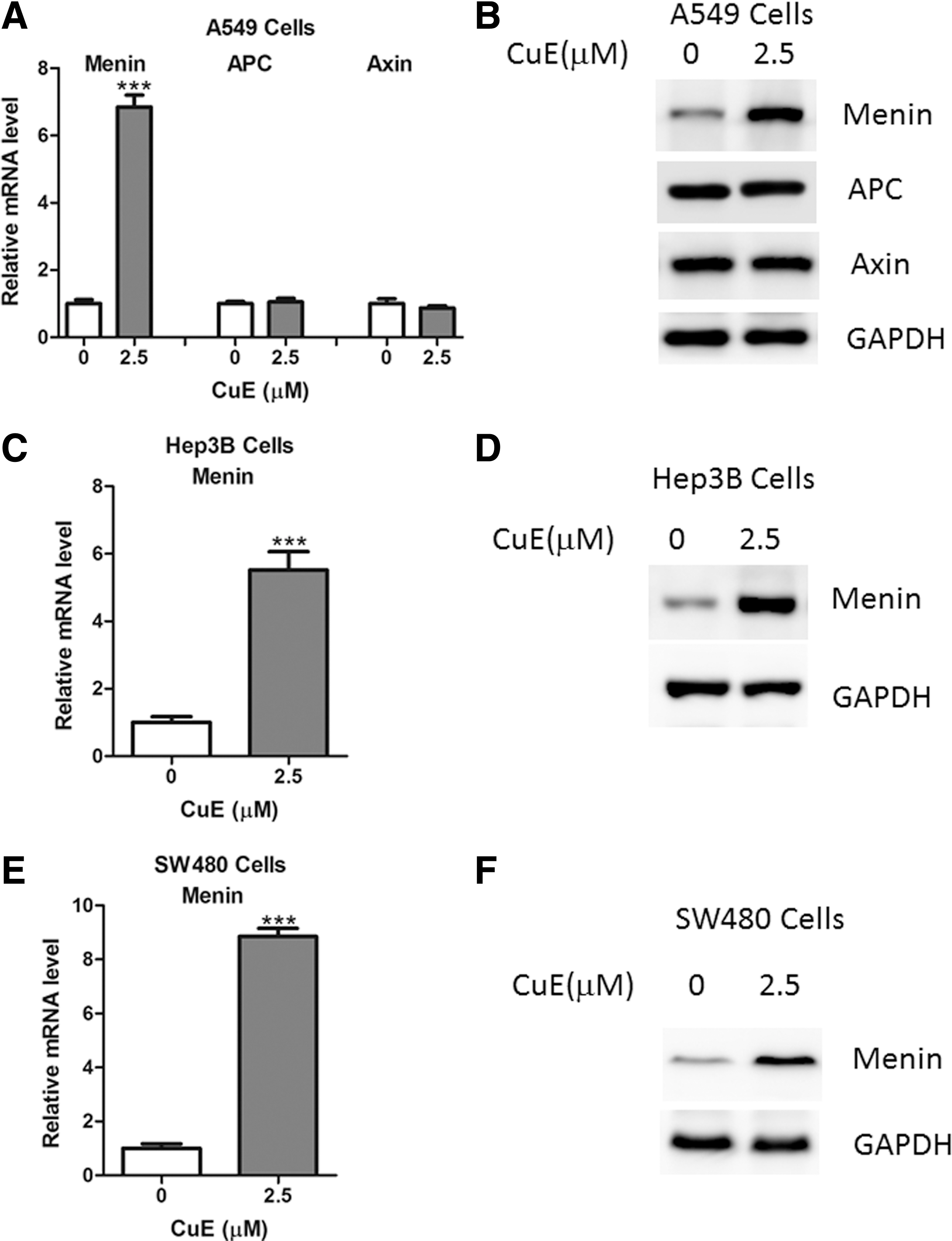
CuE upregulates Menin expression in cancer cells.
Inhibition of Menin reversed the antiproliferative roles of CuE
Next, the authors sought to determine whether the effect of CuE on cell proliferation relies on its upregulation of Menin. To confirm this hypothesis, siRNA oligos-mediated knockdown of Menin was performed in A549 cells (Fig. 4A). As a result, the authors observed that CuE could not regulate cell proliferation in A549 cells with Menin depletion (Fig. 4B). Moreover, similar results were also obtained in Hep3B and SW480 cells (Fig. 4C–F). Therefore, the results of this study suggest that the antiproliferative roles of CuE, at least in part, depend on Menin expression.
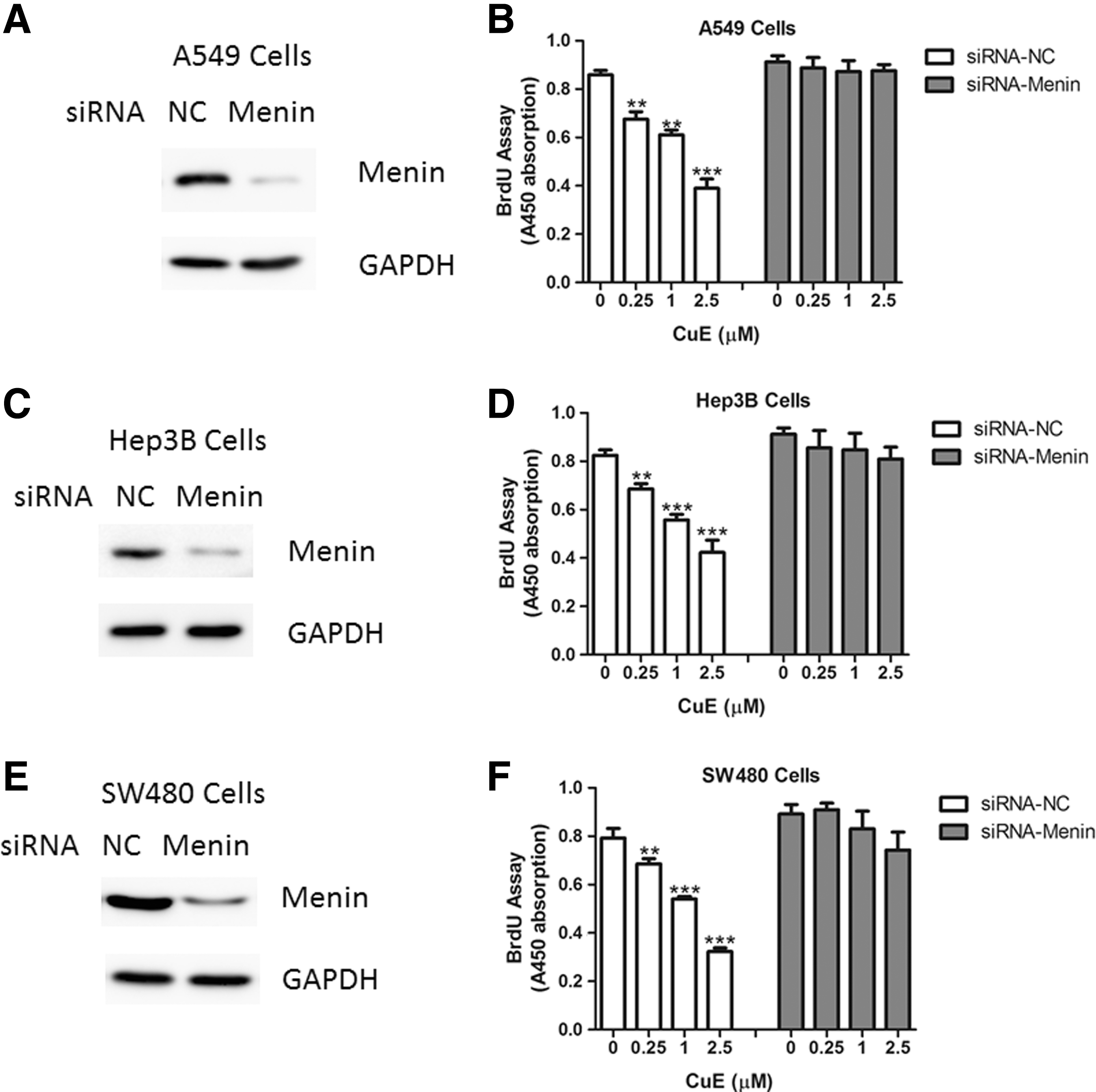
Inhibition of Menin by small interfering RNA (siRNA) oligos reversed the antiproliferative roles of CuE. Protein levels of Menin in A549
Discussion
In the present study, the authors explored the roles and molecular mechanisms of CuE in cancer cells. The antitumor activity of CuE has been documented in vitro and in animal models by previous studies. It has been shown that CuE impaired Arp2/3-dependent actin polymerization and suppressed Src/FAK/Rac1/MMP involved pathway, leading to the suppression of cell migration and invasion in breast tumor metastasis. 17 Therefore, the results of this study add a novel mechanism for the antitumor effects of CuE.
Hyperactivation of Wnt/β-catenin pathway is the common hallmark in various human cancer tissues. 18 Nuclear retention of β-catenin was shown to predict a poor outcome and favorable risk indicator. 19 In addition, Wnt/β-catenin pathway is also activated in the cells that are employed in the present study. Menin, encoded by MEN1 gene, is identified as a tumor suppressor in pituitary tumor, parathyroid tumor, adrenocortical carcinoma, insulinoma, and lung carcinoma. 20 Heterozygous germline-inactivating mutation in the MEN1 gene usually results in multiple endocrine neoplasia type 1 (MEN1), which is an inherited tumor syndrome that includes susceptibility to pancreatic islet tumors. 21 At the molecular level, Menin could directly interact with β-catenin and carry β-catenin out of the nucleus via nuclear–cytoplasmic shuttling in a CRM1-dependent manner. 22 Indeed, Menin overexpression reduces β-catenin nuclear accumulation and represses its transcriptional activity. 22 In agreement, the authors found that CuE could induce Menin expression, by which it attenuates the nuclear retention of β-catenin. However, the mechanisms of upregulation of Menin by CuE remain unclear, which needs to be investigated in the future.
In summary, the results of this study revealed that CuE exhibits direct antiproliferative actions on cancer cells through inhibition of Wnt/β-catenin activation. The presented data reinforce the fact that CuE might be a good candidate for clinical trial in cancer treatment.
Footnotes
Disclosure Statement
The authors declare that they have no conflicts of interest.