
Abstract
Leucine-rich pentatricopeptide repeat motif-containing protein (LRPPRC) is a multifunctional protein involved in the mitochondrial gene expression and function, cell cycle progression, and tumorigenesis. However, the functional role of LRPPRC in prostate cancer (
Introduction
Prostate cancer (PCa) is the second most frequent tumor and the sixth leading cause of cancer-related death in men. PCa incidence has increased in the past few years. Despite recent therapeutic advances in the management of PCa, ∼15%–30% of patients with localized disease at diagnosis develop recurrence within 5–10 years, and most of these patients subsequently show poor therapeutic outcome. 1,2 Therefore, there is an urgent need to further understand the molecular mechanisms underlying the process responsible for the development of PCa.
PCa is a double-stage disease, which usually starts as a treatable and poorly aggressive neoplasm. For early-stage localized
Previous studies have demonstrated that LRPPRC is highly expressed in most cancers, such as hepatocellular cancer, lung adenocarcinoma, esophageal squamous cell carcinoma, colon cancer, and lymphoma, and is associated with tumorigenesis.
8
Recently, one study has reported that LRPPRC is highly expressed in
Materials and Methods
Cell culture
Human PCa cell lines DU-145 and PC-3 were purchased from the Cell Bank of the Type Culture Collection of Chinese Academy of Science (Shanghai, China). Cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum, 1% 1 M HEPES buffer, and 1% minimum essential medium nonessential amino acids in a 37°C, humidified incubator under 5% CO2.
Small interfering RNA
For the siRNA-knockout experiment, a double-stranded RNA duplex that targeted the human LRPPRC gene was used (sense: 5′-CACCGGAGGAGCATTTGAGACAATAT-3′; antisense: 5′-GATCCAAAAAAGGAGGAGCATTTGA-3′); negative control siRNA was also synthesized. In vitro cotransfections were performed using Lipofectamine™2000, according to the manufacturer's protocols.
Real-time quantitative PCR analysis
Total RNA was extracted from DU-145 and PC-3 cells using the RNA plus kit (Takara, Dalian, China). Two micrograms of total RNA per sample was reverse-transcribed into cDNA using the RT2 first strand kit (Takara). Then, cDNA dilutions (1:100) were mixed with SYBR Green PCR Master Mix (Invitrogen). The specific primers for human LRPPRC were: sense 5′-GAGAGATGCCGGAATTGAGC-3′ and antisense 5′-CTCGGACTTCTCCACCTTCT-3′; for β-actin, these were: sense 5′-GAT CAT TGC TCC TCC TGA GC-3′ and antisense 5′-ACT CCT GCT TGC TGA TCC AC-3′. Assays were performed with a 7300 RT-PCR System (Takara). PCR amplification was conducted at 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds and 60°C for 1 minute. Results were analyzed using the 7300 System software, and the expression of target genes was normalized to β-actin.
Western blotting
Cell extracts were prepared with the RIPA lysis buffer (Beyotime Biotechnology, Shanghai, China) according to the operating instructions. Protein concentrations were measured by a BCA protein assay kit (Beyotime Biotechnology). For western blotting, a total of 40 μg of protein samples from each cell line was subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes (Millipore, Bedford, MA), and blocked with 5% nonfat dry milk in 0.05% Tween 20 phosphate buffered saline for 1 hour at room temperature. After blocking, the membrane was probed with primary antibodies to LRPPRC, Bcl-2, Bax, caspase-3/-8/-9, and β-actin, followed by horseradish peroxidase-conjugated secondary antibody. An enhanced chemiluminescence system was applied according to the manufacturer's protocol (Boehringer Mannheim, Mannheim, Germany). All experiments were independent and performed at least three times.
Proliferation assay and BrdU uptake
For the BrdU incorporation, the DU-145 and PC-3 cells were plated at a density of 1.0×104 per well in a 24-well tissue culture plate. Cell proliferation was confirmed by quantifying cell numbers using a cell counter (Kaihong, Beijing, China), and BrdU incorporation into DNA was quantified using the Cell Proliferation ELISA BrdU kit (Takara). Cells were treated with siRNA-LRPPRC. After 24 hours of serum starvation, the cells were incubated in the presence of BrdU at 37°C in a humidified incubator containing 5% CO2 for 24 hours.
Invasion assay
The cell invasion assay was performed using a Boyden chamber coated with matrigel, according to the manufacturer's protocol. Cells transfected with siRNA-mock and siRNA-LRPPRC were serum-starved, and ∼1×106 cells/mL were suspended in serum-free media; the cell suspension was added on to the top of the chamber and the lower chamber was filled with 500 μL DMEM media and incubated at 37°C in 5% CO2. After 48 hours, media was aspirated and noninvasive cells were removed from the top chamber using cotton swabs and stained with the staining solution provided in the kit. Invasive cells in the bottom of the chamber were counted in high-power fields under an inverted microscope.
Detection of apoptotic cells by flow cytometry
To quantify the percentage of cells undergoing apoptosis, we used Annexin V-FITC kit (Beckman-Coulter, Brea, CA). Briefly, the cells were seeded (2×105 cells/well) in a six-well plate and incubated for 48 hours. Then, the cells were treated with siRNA-LRPPRC or siRNA-mock. Cells were trypsinized and harvested by centrifugation and then incubated with Annexin V-FITC and PI for 15 minutes at room temperature in the dark. After the end of incubation, 400 μL of binding buffer was added, and the cells were analyzed immediately by flow cytometry.
Statistical analysis
All results are reported as means±standard deviation. Statistical analysis involved Student's t-test for the comparison of two groups or one-way ANOVA for multiple comparisons. p<0.05 was considered to be significant.
Results
Expression of LRPPRC in PCa cells
To test the functional role of LRPPRC in PCa, we treated PCa cells with an LRPPRC-specific siRNA to suppress the expression of LRPPRC and achieved a high efficiency of suppression. As shown in Figure 1A, real-time quantitative PCR (RT-PCR) and western blot results confirmed a remarkable downregulation of LRPPRC expression in DU-145 cells after transfection with siRNA-LRPPRC. The siRNA-LRPPRC reduced LRPPRC mRNA levels to 32.9%±3.7% of the control and reduced LRPPRC protein levels to 23.7%±2.1% of the control (*p<0.05). Consistent with the results of DU-145 cells, siRNA-LRPPRC also significantly reduced the expression of LRPPRC level in PC-3 cells (Fig. 1B).

Knockdown LRPPRC gene in DU-145 and PC-3 cell lines.
Effect of LRPPRC on the proliferation of PCa cells
We performed a BrdU incorporation assay to identify the effect of LRPPRC on the proliferation of PCa cells. As indicated in Figure 2, compared with the control, knockdown of LRPPRC did not significantly decrease BrdU incorporation. These results indicate that LRPPRC had no obvious effects on the proliferation of PCa cells.
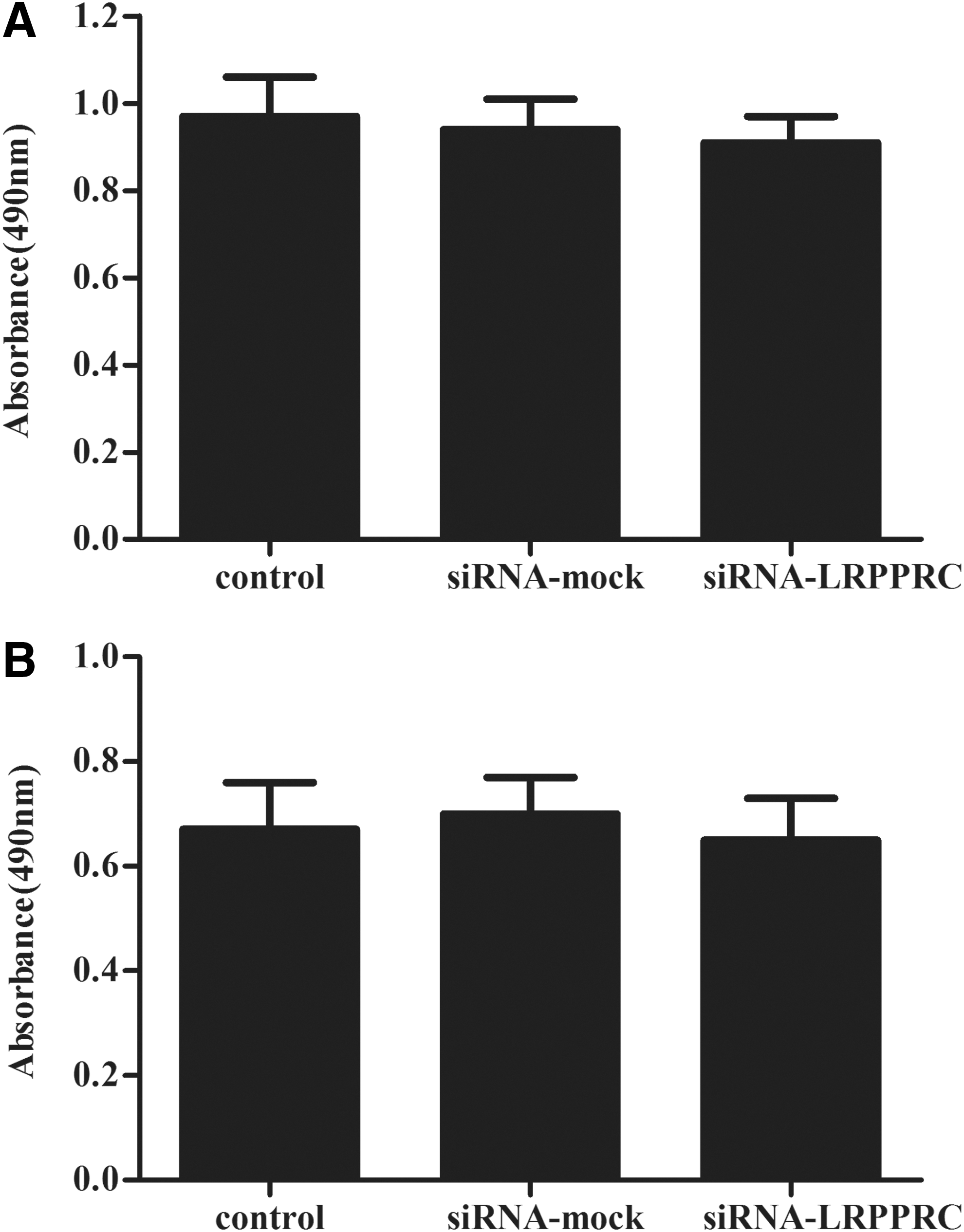
Effect of LRPPRC on the proliferation of prostate cancer (PCa) cells in vitro.
Effect of LRPPRC on the invasion of PCa cells
To explore the effect of LRPPRC on PCa cell invasion, PCa cells were transfected with siRNA-mock or siRNA-LRPPRC and were placed in a Boyden chamber coated with matrigel. As shown in Figure 3, the number of PCa cells that had invaded through the matrigel was significantly reduced to 75 and 47 in the siRNA-LRPPRC group, as compared with the mock group (*p<0.05), suggesting that LRPPRC enhances the invasion of PCa cells.

Knockdown of LRPPRC by siRNA inhibited the invasion of PCa cells in vitro.
Effect of LRPPRC on the apoptosis of PCa cells
The execution of apoptosis is associated with characteristic morphological and biochemical changes mediated by a series of gene regulation and cell-signaling pathways. 10 To further investigate the effect of LRPPRC on PCa cell apoptosis, PCa cells were treated with siRNA-LRPPRC and stained by flow cytometry-based Annexin V-FITC/PI double staining. As shown in Figure 4A, after treatment with siRNA-LRPPRC for 24 hours, the percentage of DU-145 cells in early apoptosis and late apoptosis was 16.9% and 11.2%, respectively, in comparison with the control group (6.8%/4.9%). Also in PC-3 cells, the proportion of apoptotic cells was higher than that of the control group (Fig. 4B).

Knockdown of LRPPRC by siRNA promoted the apoptosis of PCa cells in vitro. PCa cells apoptosis were detected through PI staining and the Annexin V method after 24 hours of siRNA-LRPPRC transfection, followed by flow cytometry.
To further confirm the apoptotic effect of LRPPRC on PCa cells, the expression of Bcl-2 and Bax, key factors of the apoptotic signaling pathway, was analyzed. As shown in Figure 4C and D, expression of the Bax protein was higher in the siRNA-LRPPRC group, whereas expression of the Bcl-2 protein was lower in the siRNA-LRPPRC group, compared with the control group. These results demonstrated that LRPPRC suppressed the apoptosis of PCa cells.
LRPPRC suppresses apoptosis of PCa cells through mitochondrial apoptosis pathway
Since caspases are known to play a central role in mediating various apoptotic responses, we investigated which caspases are involved in the LRPPRC-suppressed apoptosis of PC-3 cells. We first examined whether LRPPRC affects the activation of procaspase-8 in PC-3 cells. The expression levels of procaspase-8 were not changed after siRNA-LRPPRC treatment. Next, we found that the knockdown of LRPPRC significantly increased the levels of the cleaved form of caspase-3 and caspase-9. These results suggest that LRPPRC-suppressed apoptosis is associated with the activation of caspase-9 and caspase-3 (Fig. 5).
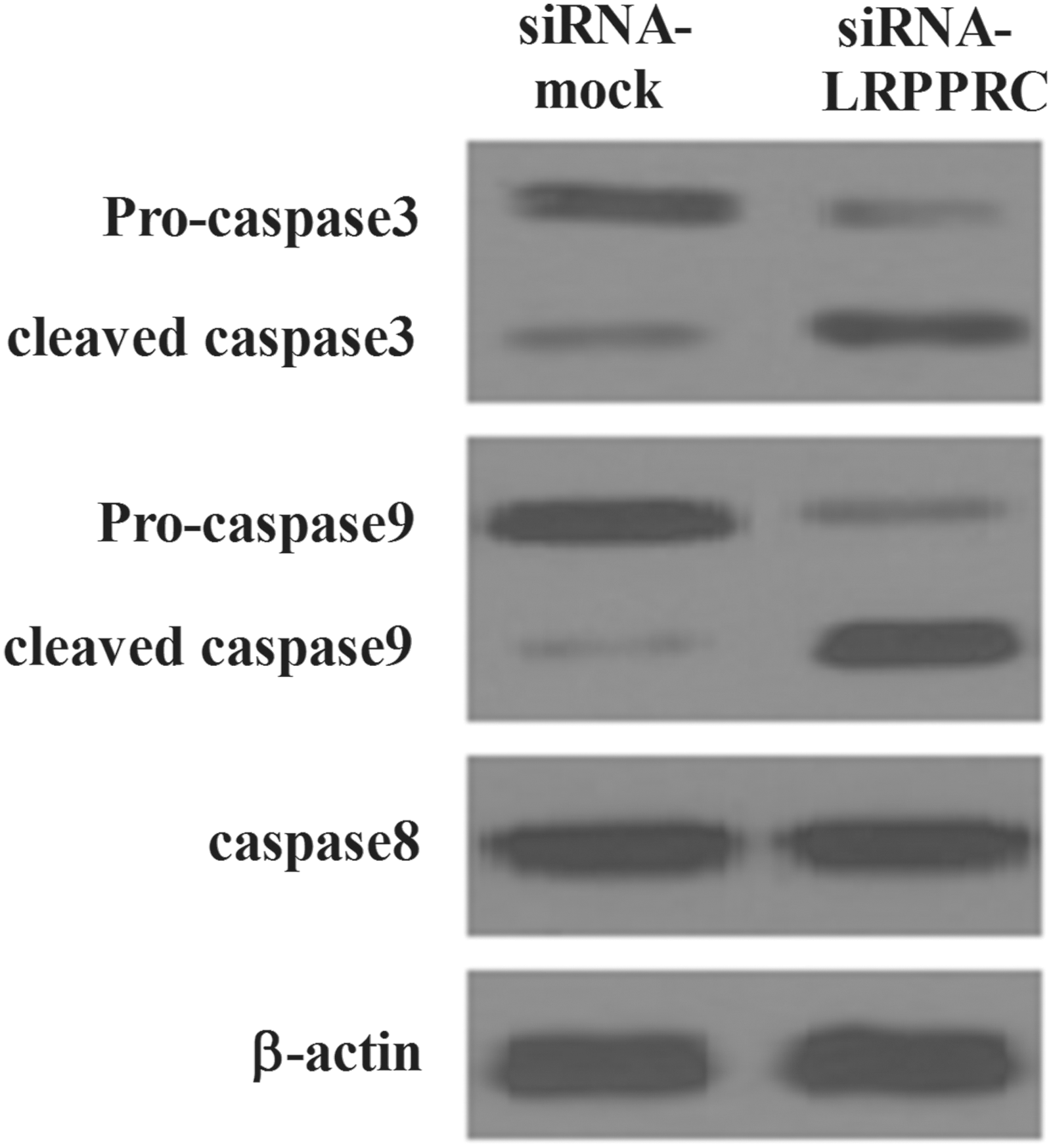
Effects of LRPPRC on expression of caspase-3/-8/-9 proteins in PC-3 cells. Cells were treated with siRNA-mock or siRNA-LRPPRC for 24 hours. Equal amounts of whole cell extracts were fractionated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and protein were detected by western blot analysis. The expression levels of proteins were normalized based on the β-actin levels. All experiments were repeated at least three times.
Discussion
Regulatory proteins which have been associated with programmed cell death are under intense research as potential cancer therapeutic targets. LRPPRC is present in the nucleus and is activated in regulating mitochondrial metabolism by the control of nuclear gene expression. 11 Therefore, it plays an important role in the function of mitochondria, which might be a prerequisite for tumor development. In this study, we have shown that knockdown of LRPPRC inhibits the invasion of PCa cells. Moreover, knockdown of LRPPRC promotes the apoptosis of PCa cells through the mitochondria-mediated pathway.
In advanced PCa, tumor cells are highly invasive and subsequent cancer metastasis leads to death. Deregulated cell invasion contributes to the attainment of the metastatic ability of the tumor cells. 12 In this study, we found that knockdown of LRPPRC inhibits the invasion of PCa cells, suggesting that LRPPRC plays an important role in cancer metastasis.
It has been reported that LRPPRC has antiapoptotic properties in hepatocellular cancer. 13 Consistent with other reports, our results showed that siRNA-LRPPRC obviously promotes the apoptosis of PCa cells. The apoptotic protein Bax is a Bcl-2 family cytoplasmic protein, which, after activation by diverse stimuli, inserts into mitochondria, leading to mitochondrial depolarization and the release of proapoptotic proteins from the mitochondria; this results in the initiation of caspase activation and apoptosis plays a critical role in the intrinsic apoptotic pathway. 14,15 Therefore, alterations in the levels of anti- and proapoptotic proteins influence apoptosis. 16 Bcl-2 has also been implicated in the development of androgen-independent PCa because of its increased expression in the advanced stages of disease, 17 and the overexpression of Bcl-2 in PCa protects the tumor cells from apoptosis. 18 In the present study, we found that LRPPRC-suppressed apoptosis in PCa cells was accompanied by the downregulation of Bax and the upregulation of Bcl-2, suggesting that LRPPRC suppressed apoptosis in PCa cells by modulating Bcl-2 family proteins.
Apoptosis can be divided into extrinsic and intrinsic pathways. The extrinsic pathway involves the initiation of death receptor stimulation and caspase-8 activation, whereas the intrinsic pathway involves mitochondrial dysfunction, resulting in the release of proapoptotic proteins from mitochondria and the activation of caspase-9. 19 The execution of apoptosis requires the accurate coordination of the caspase protein family. 20 However, alterations of apoptosis-related protein expression levels may damage the mitochondria by increasing the permeability of the outer mitochondrial membrane. This is associated with the collapse of the mitochondrial membrane potential, resulting in the degradation of caspase-3/PARPC. 21,22 Caspase-3, known as the death enzyme, has an important role in the controlled execution of programmed cell death. 23 In this study, we found that knockdown of LRPPRC significantly increased the cleavage of procaspase-3 to the active form. These results suggest that siRNA-LRPPRC-induced apoptosis may involve a caspase-3-mediated mechanism and that the activation of caspase-9 may act upstream of caspase-3 activation, suggesting involvement of the intrinsic pathway.
In conclusion, this report demonstrates that LRPPRC is involved in the invasion and apoptosis of PCa cells. Therefore, LRPPRC may play an important role in PCa and may represent a novel therapeutic target for PCa treatment.
Footnotes
Disclosure Statement
No competing financial interests exist.