
Abstract
Aims:
To investigate the effect of chloride intracellular channel 1 (CLIC1) on the proliferation, migration, and apoptosis of prostate cancer cell lines PC-3 and DU145 and the possible molecular mechanisms.
Materials and Methods:
Using the technique of RNA interference, the expression of CLIC1 was downregulated in the PC-3 and DU145 cell lines. MTT assay, Transwell chamber, and flow cytometry were used to determine the effect of CLIC1 on the proliferation, migration, and apoptosis ability of PC-3 and DU145 cells. The levels of phosphorylated extracellular signal-regulated kinase 1/2 (p-ERK1/2), ERK1/2, matrix metalloproteinase (MMP)-2, and MMP-9 were examined by western blotting.
Results:
The results showed that the knockdown of CLIC1 exerts inhibitory effects on the proliferation and migration of PC-3 and DU145 cells. At the same time, the authors found that the knockdown of CLIC1 has no effect on the apoptosis in PC-3 and DU145 cells. Meanwhile, the levels of p-ERK1/2, MMP-2, and MMP-9 were decreased in the CLIC1 small interfering RNA (siRNA) group compared with the control and vector groups.
Conclusion:
These results indicate that CLIC1 could regulate prostate cancer cell proliferation and migration by regulating the mitogen-activated protein kinase (MAPK)/ERK pathway and offers a candidate molecular target for prostate cancer prevention and therapy.
Introduction
Prostate cancer is the most commonly diagnosed cancer in men and the second leading cause of cancer death among men worldwide. 1 It is important for patients with prostate cancer to get early detection and treatment. Carcinogenesis and mechanisms influencing the progression and prognosis of prostate cancer are multistep processes, involving both genetic insults to epithelial cells and changes in epithelial–stromal interactions. 2 Recently, several effective therapies have been offered for the clinical treatment of prostate cancer. Localized prostate cancer is eminently treatable; however, no curative therapy is available once the cancer becomes invasive and metastasizes. 3,4 Therefore, identifying the role of new molecules of diagnostic and therapeutic significance remains a major focus of current cancer research.
Chloride intracellular channel (CLIC) proteins are components or regulators of novel intracellular anion channels in mammalian cells. The CLIC gene family consists of seven proteins, with CLICs 1, 4, and 5 known to possess the chloride channel activity. 5,6 CLIC1 is the first cloned human member of the CLIC family and is a 241 amino acid ion channel protein. 7 Recent studies have indicated that CLIC1 is significantly upregulated in tumor tissues, such as gastric carcinoma, 8 colorectal cancer, 9 prostate cancer, 10 and gall bladder carcinoma. 11 It has been demonstrated that CLIC1 is involved in the regulation of the cell cycle. CLIC1 is detected on the plasma membranes of cells in the G2/M phase. 12 It was presumed that an elevated expression of CLIC1 can modulate antiapoptosis signaling and cell division, resulting in cellular transformation. 13,14 However, a true understanding of its function is still incomplete.
In addition, a recent study has indicated that CLIC1 is highly expressed in prostate tumor tissues and can also be used as a potential biomarker for prostate cancer 10 ; however, it is unclear whether CLIC1 participates in biological functions relevant to prostate carcinoma. Previous studies have shown that CLIC1 plays an important role in proliferation, migration, and apoptosis of gallbladder and gastric carcinomas. 8,11 However, to the author's knowledge, the mechanisms behind how CLIC1 promotes tumor proliferation and migration while suppressing apoptosis remain to be elucidated.
In the present study, the authors intended to observe the biological function and molecular mechanisms in the PC-3 and DU145 cell lines following CLIC1 knockdown by the RNA interference technique. These data might provide scientific information for prognosis prediction and targeted therapy for prostate cancer.
Materials and Methods
Reagents
Antibodies against CLIC1, extracellular signal-regulated kinase 1/2 (ERK1/2), phosphorylated ERK1/2 (p-ERK1/2), matrix metalloproteinase (MMP)-2, and MMP-9 were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA). The antibody against β-actin was obtained from Good HERE Biotech, Inc. (Hangzhou, China). Horseradish peroxidase-conjugated secondary antibodies were obtained from Abgent Biotech (Suzhou, China).
Cell culture
The human prostate cancer cell lines PC-3 and DU145 were purchased from the Shanghai Academy of Life Sciences (Shanghai, China) and cultured as described. The cells were cultured in RPMI 1640 (Pierce, Rockford, IL) supplemented with 10% fetal bovine serum (FBS; Gibco BRL, Gaithersburg, MD), 1%
RNA interference and transfection
PC-3 and DU145 cells were divided into three groups: the control group (treated with Lipofectamine 2000 only), the vector group (treated with Lipofectamine 2000 and control siRNA), and the CLIC1 siRNA group (treated with Lipofectamine 2000 and CLIC1 siRNA). Small interfering RNA (siRNA) was synthesized by Genechem Co. Ltd. (Shanghai, China). The sequences of the human CLIC1 siRNA were as follows: 5′-GCCAAAGUUACACAUAGUATT-3′ (sense) and 5′-UACUAUGUGUAACUUUGGCTT-3′ (antisense). 7 The cultured PC-3 cells were transfected with 50 pM siRNA using Lipofectamine 2000 according to the manufacturer's instructions (Invitrogen, Carlsbad, CA). 15
Cell proliferation
The MTT assay was used as the qualitative index of cell viability. Briefly, prostate cancer PC-3 and DU145 cells were divided into three groups: the siRNA group, the vector group, and the negative control group. Cells were plated in 96-well microtiter plates at a density of 1×104 cells/well, and after transfection for 24, 48, and 72 hours, 20 μL of MTT (5 mg/mL; Sigma) solution was added to each well and then the samples were incubated for another 4 hours. Subsequently, the cells were lysed using 150 μL of dimethyl sulfoxide. When the formazan crystals were completely dissolved, the optical density was measured at 490 nm.
Cell migration assay
Cell migration assays were performed using FBS-coated polycarbonate filters (8 μm pore size, Transwell; Becton Dickinson, Franklin Lakes, NJ), as previously described, 16 with slight modifications. Briefly, the prostate cancer cell lines were grouped and transfected with CLIC1 siRNA and control siRNA, as described previously. After 48 hours of transfection, the cell media were removed and the cells were washed twice with phosphate-buffered saline (PBS). The membrane undersurface was coated with 200 μL of FBS for 1 hour at 37°C and blocked with 200 μL migration buffer (0.5% bovine serum albumin in OPTI-MEM) for 30 minutes at 37°C. The lower chamber was filled with 500 μL of migration buffer, following which the cells were plated in the upper chamber after transfecting for 48 hours, with four wells per treatment at a density of 1×105 in 100 μL of migration buffer and incubated at 37°C for 4 hours. Following incubation, cells in the upper compartment were trypsinized and counted with the CASY 1 counter (Sharfe System, Reutingen, Germany). Cells that had migrated to the lower surface of the filter were also trypsinized and counted. The migration rate was obtained by dividing the cell number in the lower chamber by the sum of the cell numbers found in both the lower chamber and the upper chamber ×100.
Determination of cell apoptosis by flow cytometry
Cell apoptosis analyses were performed as previously described with minor modifications. 17 Briefly, the prostate cancer cell lines were grouped and transfected with CLIC1 siRNA and control siRNA, as described previously. After 48 hours of transfection, the cell media were removed and the cells were detached by trypsinization and washed twice in PBS, centrifuged at 1000 g for 5 minutes, and resuspended in 195 μL of Annexin V-FITC binding buffer. A volume of 5 μL Annexin V-FITC was added and mixed. Then, the cells were stained in the dark for 10 minutes at room temperature. After that, the cells were centrifuged at 1000 g for 5 minutes and resuspended in 190 μL of Annexin V-FITC binding buffer. Finally, 10 μL of propidium iodide staining solution was added and mixed. The cells were kept on ice in the dark and immediately subjected to the flow cytometric analysis. The data were analyzed using the Cell Quest software. The experiment was repeated thrice.
Quantitative reverse transcription–polymerase chain reaction
Total RNA was extracted using the RNA plus kit (Takara, Dalian, China) and reverse transcribed to generate cDNA (Takara). β-Actin was used as an internal control. The levels of CLIC1 mRNA were quantified by the real-time PCR with the Applied Biosystems 7900HT Fast Real-Time PCR System using the SYBR Premix Ex Taq (Applied Takara Bio). The primers for the CLIC1 sequence used were as follows: 5′-AATCAAACCCAGCACTCAATG-3′ (sense) and 5′-CAGCACTGGTTTCATCCACTT-3′ (antisense). 7 The polymerase chain reaction (PCR) conditions included an initial denaturation step at 94°C for 2 minutes, followed by 35 cycles at 94°C for 30 seconds, 56°C for 30 seconds, and 72°C for 2 minutes, and a final elongation step at 72°C for 10 minutes. Relative quantification of the CLIC1 mRNA expression was calculated using the 2−ΔΔCT method. 18
Western blotting
The total proteins were extracted from cell cultures using the RIPA buffer (Beytime, Shanghai, China). The protein amounts were measured using the bicinchoninic acid method. Thirty micrograms of total protein was separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to 0.45 μm nitrocellulose filter membrane. The membrane was blocked with 5% skim milk for 1 hour. Then, the blots were incubated overnight at 4°C with each primary antibody, followed by incubation for 1 hour with the horseradish peroxidase-conjugated secondary antibody. To check for equal protein loading, enhanced chemiluminescence advanced detection reagents (Boehringer Mannheim, Mannheim, Germany) were used to induce chemiluminescence, which was subsequently caught on Hyperfilm ECL. The intensities of the protein bands were quantified by the Sion Image analysis software (Scion Corporation, Frederick, MD).
Statistical analysis
Experiments were carried out at least in triplicate, and the results are expressed as mean±SD. Statistical analysis was performed using SPSS 16.0 (SPSS, Inc., Chicago, IL). Pairs of samples were compared using the Student's t-test. A p<0.05 was considered statistically significant.
Results
Determination of transfection effects
To test the efficiency of CLIC1 transfection, western blotting and quantitative reverse transcription–polymerase chain reaction were employed to determine the expression levels of the protein and mRNA. As shown in Figure 1, the expression of CLIC1 protein and mRNA was decreased significantly in the CLIC1 siRNA-transfected group. There was no significant difference in the expression levels of the CLIC1 protein and mRNA between the control group and the vector group, which also demonstrated that Lipofectamine and control siRNA did not affect the expression of CLIC1 protein and mRNA. Collectively, these data also demonstrated that the CLIC1 protein and mRNA were successfully suppressed in the PC-3 and DU145 cell lines.
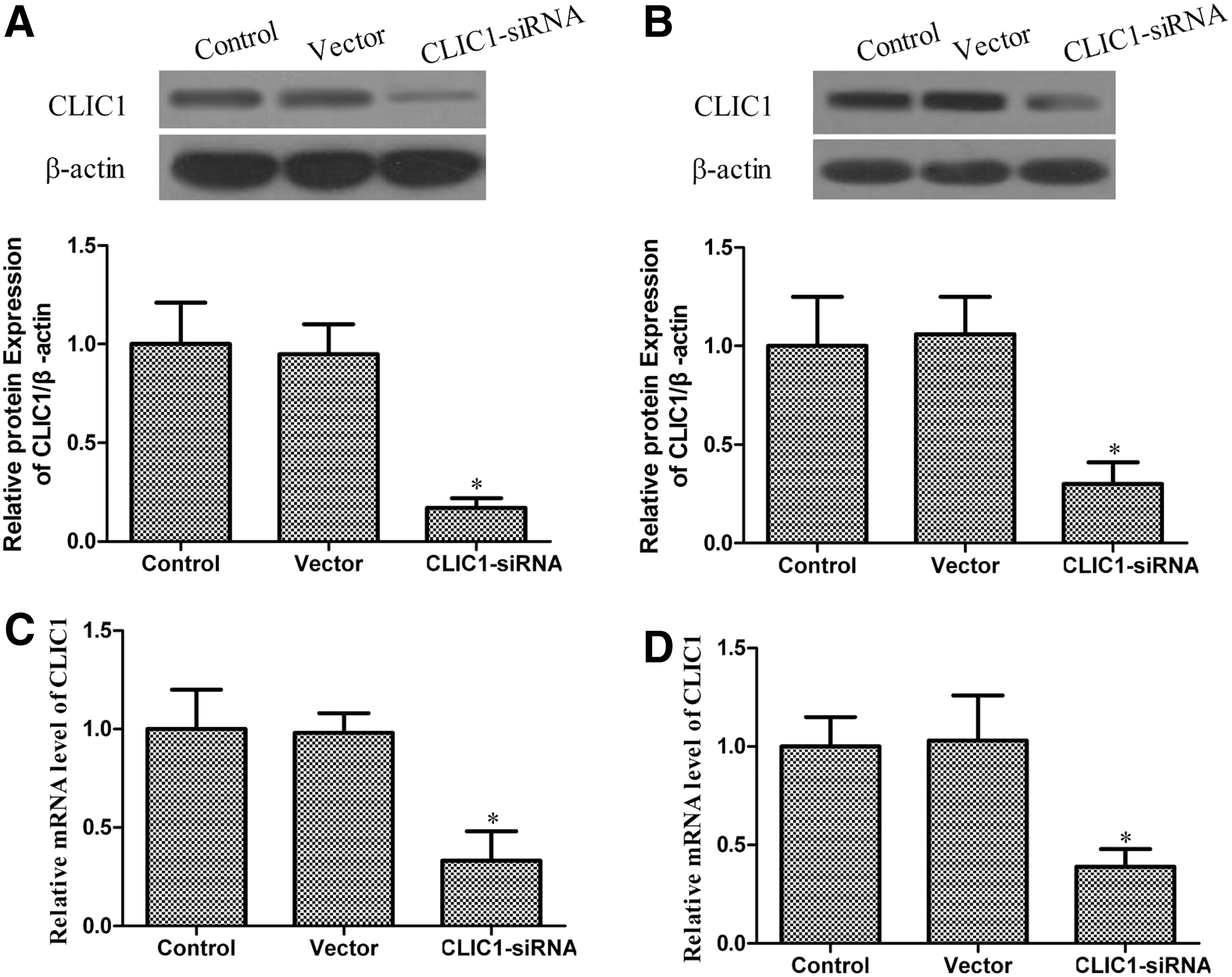
The expression of chloride intracellular channel 1 (CLIC1) in the PC-3 and DU145 cell lines. Western blot analysis of CLIC1 expression 48 hours after transfection with CLIC1 small interfering RNA (siRNA) and control siRNA. β-Actin was used as a loading control. CLIC1 mRNA expression was detected by the quantitative reverse transcription–polymerase chain reaction 48 hours after transfection with CLIC1 siRNA and control siRNA. β-Actin was used as loading control. The figure represents one of the three experiments performed independently with similar results.
The effect of CLIC1 on cell proliferation
The impact of CLIC1 on PC-3 and DU145 cell proliferation was determined using the MTT assay every 24 hours after transfection, for up to 72 hours. The obtained results revealed that the viability of the cells was, to some extent, inhibited by CLIC1 in a time-dependent manner. As shown in Figure 2, the CLIC1-transfected group grew more slowly than the control group and the vector group. These results demonstrated that the downregulation of CLIC1 could inhibit PC-3 and DU145 cell proliferation.
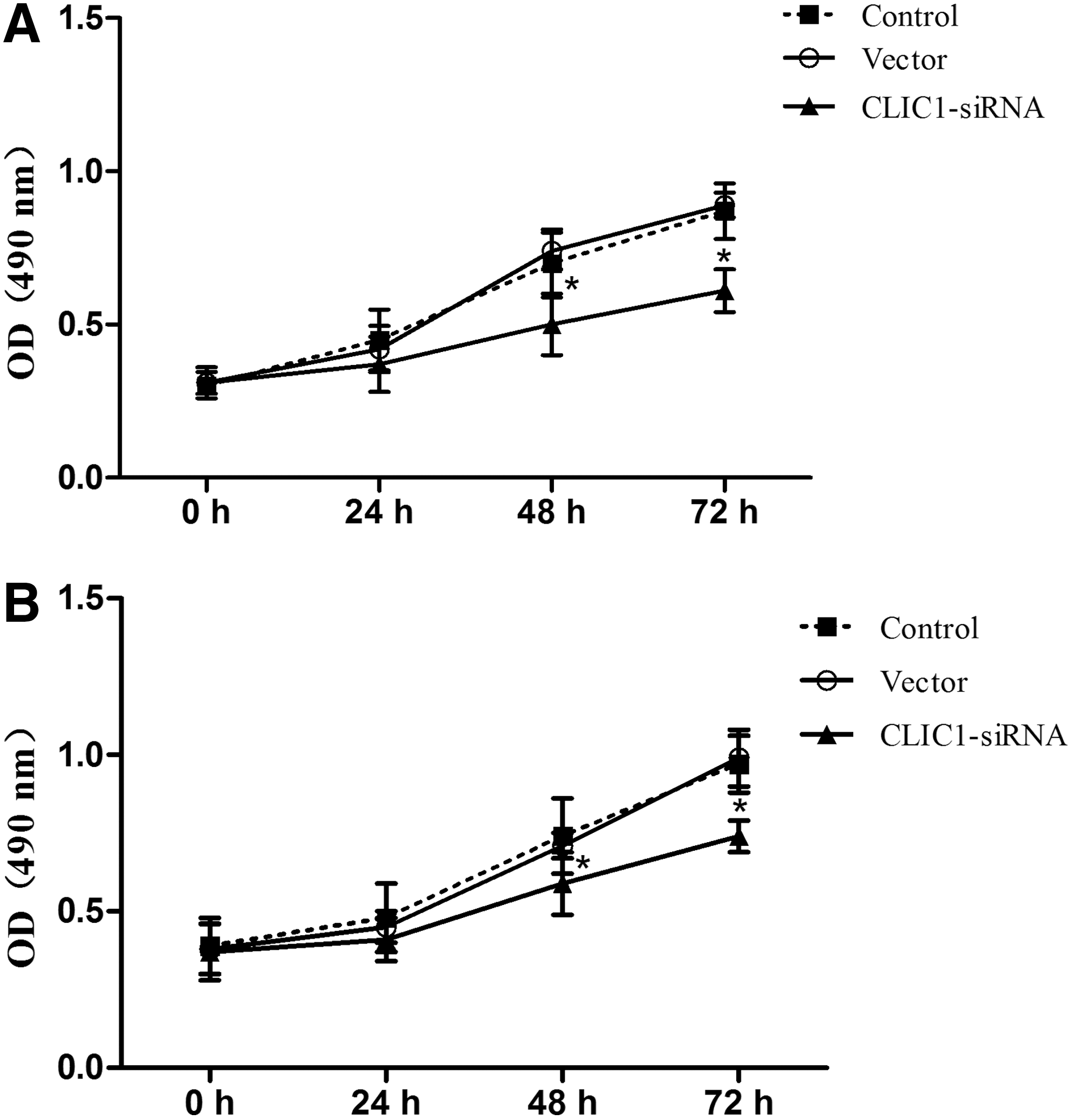
Effects of CLIC1 on PC-3 and DU145 cell proliferation. The MTT assay was conducted to assess the effect of CLIC1 on cell proliferation every 24 hours after transfection, for up to 72 hours.
The effect of CLIC1 on cell migration
The authors determined the effect of decreased CLIC1 expression on prostate cancer cell migration after 48 hours of transfection. Cancer cell migration was significantly reduced by CLIC1 siRNA according to the migration assay (Fig. 3A, B). The decreased CLIC1 expression inhibited cell migration by 27.25% (p<0.05) in PC-3 cells and 29.37% (p<0.05) in DU145 cells. Taken together, these results clearly indicate that the suppression of CLIC1 inhibited the migration ability of PC-3 and DU145 cells.

Effects of CLIC1 on PC-3 and DU145 cell migration. Cell migration was measured with the transwell assay 48 hours after CLIC1 transfection. All experiments were repeated at least thrice. All data are means±SD. (n=3) (*p<0.05 vs. control).
The effect of CLIC1 on cell apoptosis
The authors investigated whether CLIC1 could induce apoptosis in human prostate cancer PC-3 and DU145 cells. Cell apoptosis was detected through PI staining and the Annexin V method 48 hours after transfection in PC-3 and DU145 cells, followed by flow cytometry. As shown in Figure 4, there were no differences in the percentages of apoptotic cells among the CLIC1-transfected group, the control group, and the vector group in both PC-3 and DU145 cells. These data reveal that the downregulation of CLIC1 had no effect on the apoptosis in PC-3 and DU145 cells.
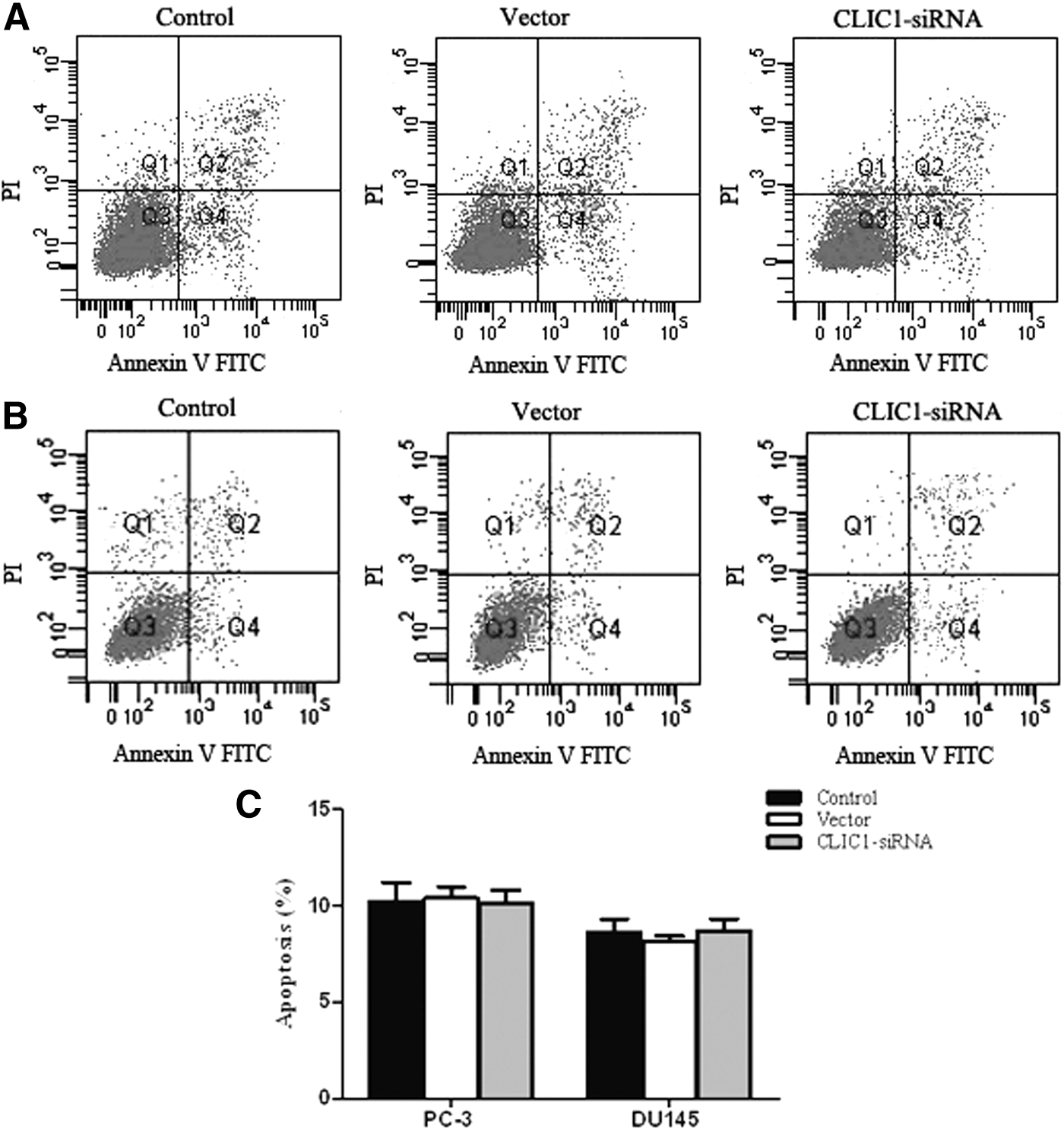
Effect of CLIC1 on PC-3 and DU145 cell apoptosis. PC-3 and DU145 cell apoptosis was detected through PI staining and the Annexin V method after 48 hours of CLIC1 siRNA transfection, followed by flow cytometry.
CLIC1 regulates the MAPK/ERK pathway, MMP-2, and MMP-9 synthesis in PC-3 cells
The ERK pathway is frequently upregulated in a variety of cancers, 19 and there is some evidence showing that mitogen-activated protein kinase (MAPK)/ERK pathways are involved in the proliferation and migration of cancer. 20 To further investigate a potential mechanism that could be involved in the growth and migration inhibition of prostate cancer cells, the expression of ERK1/2, p-ERK1/2, MMP-2, and MMP-9 was detected by western blotting. The results suggested that p-ERK1/2, MMP-2, and MMP-9 proteins were each strongly downregulated in the CLIC1 siRNA group. Furthermore, the expression of p-ERK1/2, MMP-2, and MMP-9 did not differ significantly between the control group and the vector group. The level of the ERK1/2 protein was statistically similar in all the three groups. These results are shown in Figure 5.
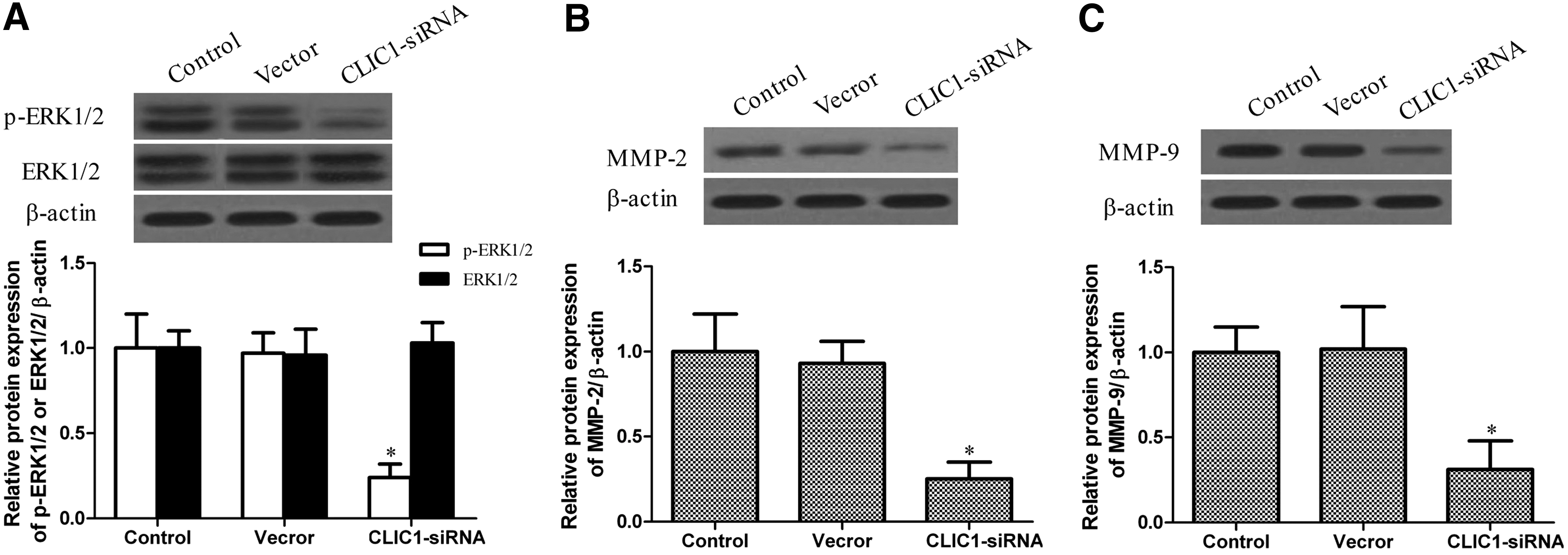
The effect of CLIC1 on extracellular signal-regulated kinase 1/2 (ERK1/2), matrix metalloproteinase (MMP)-2, and MMP-9 expression. The protein levels of phosphorylated ERK1/2 (p-ERK1/2), ERK1/2, MMP-2, and MMP-9 were determined by the western blotting analysis. β-Actin was used as a loading control. All experiments were repeated at least thrice. *p<0.05 versus control.
Discussion
Prostate cancer is one of the most commonly diagnosed malignant diseases in men and the leading cause of cancer death in men. 21,22 Currently, several surgical therapies and effective radiation can be offered for the clinical treatment of prostate cancer. However, the therapies for prostate cancer are far from satisfactory, and advanced prostate cancer remains incurable. 23 Therefore, the search for new methods of cancer therapy and prognosis is a key task for many researchers around the world.
CLIC1, the first identified human member of a growing family of organelle ion channels, is a transmembrane protein that forms a functional ion channel as a tetrameric assembly of subunits. Recently, Ummanni et al. 10 found that CLIC1 was upregulated in prostate cancer and might be a potential biomarker for prostate cancer. CLIC1 overexpression has been demonstrated in a wide variety of tumor types, including glioma 24 and colorectal cancer. 25 An elevated CLIC1 expression was strongly correlated with lymph node metastasis, perineural and lymphatic invasion, pathological staging, and poor survival. 26 However, the biological function and molecular mechanisms of CLIC1 in prostate cancer remain unclear.
In the present study, the authors used the RNA interference technology to inhibit the expression of CLIC1 and evaluated the effects of downregulating CLIC1 on proliferation, migration, and apoptosis of the PC-3 and DU145 cell lines, along with its mechanism of action. The studies showed that the knockdown of CLIC1 inhibited the proliferation of PC-3 and DU145 cells. These results suggested that CLIC1 plays a crucial role in cell growth in the PC-3 and DU145 prostate cancer lines. Some investigators 11 have found that overexpression of CLIC1 significantly increased cell motility, and the knockdown of CLIC1 markedly inhibited cell migration and invasion. In this study, the results demonstrated that the downregulation of CLIC1 grossly suppressed the migratory capacity of PC-3 and DU145 cells in vitro. These results were consistent with previous experiments. At the same time, the authors found that the knockdown of CLIC1 produced no significant change in PC-3 and DU145 cell apoptosis.
The authors further explored the molecular signaling pathways that may be involved in the proliferation and migration of the prostate cancer cell lines. It is known that MAPK/ERK can promote cancer metastasis. In the present study, the western blotting analysis showed that the MAPK/ERK pathway was suppressed in PC-3 cells in the CLIC1 siRNA group. Taken together, the findings imply that CLIC1 promotes the proliferation and mobility of prostate cancer by regulating the MAPK/ERK pathway. MMP-2 and MMP-9, members of the MMP family, are thought of as two important mediators of cancer metastases in patients with prostate carcinoma. 27 In the present study, the authors found that both MMP-2 and MMP-9 proteins were downregulated in PC-3 cells in the CLIC1 siRNA-transfected group. The results demonstrated that MMP-2 and MMP-9 play an important role in the migration of prostate cancer.
In summary, the findings show that downregulation of CLIC1 can efficiently suppress prostate cancer proliferation and migration in vitro, perhaps through the MAPK/ERK signaling pathway. However, the downregulation of the CLIC1 expression has no effect on prostate cancer cellular apoptosis. In short, CLIC1 plays an important role in regulating the biological behavior of prostate cancer cells.
Footnotes
Acknowledgment
This research was supported by the youth innovation fund projects of the First Affiliated Hospital of Zhengzhou University.
Disclosure Statement
No conflict of interests exist.