
Abstract
Virotherapy represents a promising new approach for treating cancer. Here the authors have analyzed the effect of ultraviolet-inactivated Sendai virus strain Tianjin (UV-Tianjin) on human breast cancer MCF-7 cells in vitro and in vivo. In vitro, UV-Tianjin inhibited the proliferation of MCF-7, MDA-MB-231, and T47D breast cancer cell lines, although MCF-7 cells were most susceptible to UV-Tianjin treatment. Hoechst staining and flow cytometric analysis of UV-Tianjin-treated MCF-7 cells revealed that UV-Tianjin induced apoptosis in a dose-dependent manner. Moreover, UV-Tianjin treatment resulted in reductions in the mitochondria membrane potential of MCF-7 cells and regulated the levels and activities of Bcl-2, Bax, cyt c, caspases, Fas, and Fas ligand (FasL). In vivo, UV-Tianjin inhibited the growth of MCF-7 tumors in nude mice and increased tumor cell apoptosis compared with saline-treated controls. In addition, the percentage of tumor cells positive for cleaved versions of caspase-7, caspase-8, and caspase-9 was higher in UV-Tianjin-treated tumors than in saline-treated controls. In summary, UV-Tianjin exhibited the antitumor activity in human breast cancer MCF-7 cells both in vitro and in vivo. The UV-Tianjin treatment seemed to induce apoptosis by activating both the mitochondrial and death receptor apoptotic pathways.
Introduction
Breast cancer is one of the most common malignancies and its incidence is increasing worldwide. 1 The most common treatment for breast cancer is surgery, followed by postoperative adjuvant chemotherapy. However, chemotherapy is associated with a variety of adverse side effects, and some cancer cells are drug resistant. 2,3 Therefore, it is important to develop new antitumor drugs with lower levels of toxicity and fewer side effects.
Virotherapy represents a promising new approach for treating cancer. 4,5 Some viruses have been used for cancer treatment as they can selectively replicate in cancer cells. These include Newcastle disease virus, vesicular stomatitis virus, and measles virus. 6 –8 However, recent studies have shown that ultraviolet-inactivated, replication-defective Sendai virus [also known as the hemagglutinating virus of Japan envelope (HVJ-E)] induces strong antitumor immune responses by activating cytotoxic T lymphocytes and natural killer cells and suppressing regulatory T cells. 9 –11 Furthermore, HVJ-E induces apoptosis in castration-resistant human prostate cancer cells and human glioblastomas in a dose-dependent manner, without affecting nonmalignant cells. 12,13 The molecular mechanisms by which HVJ-E induces cancer cell apoptosis have not been fully elucidated.
Sendai virus strain Tianjin was isolated from the lungs of a marmoset, 14 and the authors have shown that the ultraviolet-inactivated Tianjin strain (UV-Tianjin) suppresses the growth of colon carcinomas in mice by inducing an antitumor immune response and tumor cell apoptosis. 15 In addition, UV-Tianjin induces apoptosis of rat glioma cells both in vitro and in vivo. 16 In the present study, it is demonstrated that UV-Tianjin inhibited the proliferation of the human breast cancer cell lines MCF-7, MDA-MB-231, and T47D. Interestingly, MCF-7 cells, which are deficient for caspase-3, were more sensitive to UV-Tianjin treatment than the other two cell lines. The authors also showed that UV-Tianjin induced apoptosis of MCF-7 cells through both the mitochondrial and death receptor pathways. Therefore, UV-Tianjin is a promising therapeutic agent for treating human breast cancer.
Materials and Methods
Cells, viruses, and mice
Human breast cancer cell lines MCF-7, MDA-MB-231, and T47D were obtained from the Cell Bank of Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). MCF-7 cells were propagated in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (FBS; HyClone Laboratories, Tianjin, China) at 37°C in 5% CO2. T47D cells were maintained in the RPMI 1640 medium supplemented with 10% FBS at 37°C in 5% CO2. MDA-MB-231 cells were maintained in Leibovitz's L-15 medium containing 10% FBS at 37°C in non-CO2 conditions. The Sendai virus strain Tianjin (GenBank: EF679198) was propagated in chorioallantoic fluid from 9- to 11-day-old chicken eggs. Viruses were purified by centrifugation and then inactivated by UV irradiation (99 mJ/cm2), as described. 17 Inactivated viruses could not replicate, but their capacity for viral fusion remained intact. Five- to 6-week-old female BALB/c nude mice were purchased from Beijing HFK Bioscience (Beijing, China).
Ethics statement
All animals were handled according to the China Guidelines for the Care and Use of Laboratory Animals and with the ARRIVE guidelines. 18 Animals were housed in pathogen-free conditions, and all experimental manipulations involving mice were performed under sterile conditions in a laminar flow hood. Intratumoral injections were performed on anesthetized mice, and all efforts were made to minimize suffering.
Reagents and antibodies
The authors purchased MTT from Sigma-Aldrich (Shanghai, China). Hoechst 33342 and caspase-3/-7, caspase-8, and caspase-9 colorimetric assay kits were purchased from KeyGen Biotech (Nanjing, China). The Annexin V-FITC/PI apoptosis detection kit was purchased from Biovision (Wuhan, China). The JC-10 MMP assay kit was purchased from Beijing Fanbo Biochemicals (Beijing, China). The caspase inhibitors Z-VAD-FMK (pan caspase), Z-LEHD-FMK (caspase-9 specific), and Z-IETD-FMK (caspase-8 specific) were purchased from Sigma-Aldrich. Antibodies specific for caspase-7 (9492), caspase-8 (9746), and caspase-9 (9502) were obtained from Cell Signaling Technology (Shanghai, China). Antibodies specific for Bcl-2 (sc-509), Bax (sc-20067), cyt c (sc-13561), Fas (sc-1023), FasL (sc-6237), and β-actin (sc-8432) were purchased from Santa Cruz Biotechnology (Beijing, China). For immunohistochemistry, antibodies specific for caspase-7 (ab52297), caspase-8 (ab25901), and caspase-9 (ab129749) were purchased from Abcam Biotechnology (Beijing, China). In situ apoptosis detection kits were purchased from Promega (Beijing, China).
Cell viability assay
The MTT assay was performed, as described, 19 with modifications. Briefly, human breast cancer cells (2.0×105 cells/mL) were seeded in a volume of 100 μL into 96-cell culture plates. After 24 hours, UV-Tianjin [multiplicity of infection (MOI): 5–200] was added to the monolayer, and cells were cultured for an additional 24 hours. The authors then added 10 μL of MTT [5 mg/mL in phosphate-buffered saline (PBS)] to each well, and cells were cultured for an additional 4 hours. After the MTT-containing medium was removed, 100 μL of DMSO was added to dissolve the formazan and the culture plates were shaken for 10 minutes at room temperature. Finally, the optical density (OD) of each well was measured using a microplate reader (Bio-Rad, Hercules, CA) at 490 nm. The following formula was used to calculate cell viability: viability (%)=(sample OD490−blank OD490)/(control OD490−blank OD490)×100%.
Hoechst staining
MCF-7 cells were seeded onto coverslips in six-well plates at a density of 2.5×105 cells per well. After 24 hours, cells were treated with UV-Tianjin at various MOIs (10, 20, or 40) for 24 hours. The cells were then washed with PBS and incubated with 10 μL Hoechst 33342 in 1 mL PBS for 15 minutes. After rinsing with PBS, the cells were analyzed using fluorescence microscopy (Nikon Eclipse E600, Tokyo, Japan). Apoptotic nuclei were identified by condensed chromatin, contiguous to the nuclear membrane, as well as nuclear fragmentation of condensed chromatin.
Flow cytometric analysis of apoptosis
Apoptotic cells were examined using an Annexin V-FITC/PI apoptosis detection kit. Briefly, cells were treated with various doses of UV-Tianjin (MOI: 10, 20, or 40) for 24 hours and then 5×105 cells were harvested using trypsinization, washed twice with PBS, and resuspended in 500 μL of binding buffer. Cell suspensions were then incubated with 5 μL of Annexin V-FITC and 5 μL of propidium iodide (PI) for 10 minutes at room temperature in the dark and then immediately evaluated using flow cytometry (FACScan; BD Biosciences, San Jose, CA). For caspase inhibitor assays, cells were pretreated with 20 μM Z-VAD-FMK, Z-LEHD-FMK, or Z-IETD-FMK for 2 hours and then treated with UV-Tianjin (MOI 40) for an additional 24 hours. The extent of apoptosis was then determined using flow cytometry, as described above.
Caspase activity assay
The caspase activity was determined using caspase-3/-7, caspase-8, or caspase-9 colorimetric assay kits. Briefly, MCF-7 cells were cultured in six-well plates overnight and then treated with UV-Tianjin (MOI: 10, 20, or 40) for 24 hours. The cells were collected, washed in PBS, and placed in 100 μL of lysis buffer for 30 minutes on ice. After centrifugation, supernatants containing 150 μg of protein were incubated with 200 μM caspase-3/-7 (Ac-DEVD-pNA), caspase-8 (Ac-IETD-pNA), or caspase-9 (Ac-LEHD-pNA) substrates in a reaction buffer for 4 hours at 37°C. Levels of released pNA were measured using a microplate reader (405 nm).
Flow cytometric analysis of mitochondria membrane potential
MCF-7 cells (2.5×105/well) were plated in six-well plates. After an overnight incubation, cells were treated with UV-Tianjin (MOI: 5, 10, or 20) for 24 hours or 50 μM carbonyl cyanide 3-chlorophenylhydrazone (CCCP) for 1 hour. Cell densities were adjusted to 1×106 cells/mL, and then cells were incubated with JC-10 dye (final concentration 20 μg/mL) for 30 minutes at 37°C. After rinsing twice with the JC-10 staining buffer, red and green fluorescence was measured using flow cytometry. A decrease in the red:green fluorescence ratio indicates a loss in mitochondria membrane potential (MMP). Quantitative analysis was conducted using CellQuest analysis software.
Western blot analysis
Western blot analysis was performed, as described, 20 with modifications. MCF-7 cells were treated with UV-Tianjin (MOI: 10 or 40) for 24 hours and then harvested using a cell scraper. Cell pellets were resuspended in 100 μL of RIPA buffer [1% NP-40, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS), 150 mM NaCl, 10 mM sodium phosphate, pH 7.2, 2 mM EDTA, 50 mM sodium fluoride, 0.2 mM sodium vanadate] containing protease inhibitor cocktail and lysed for 30 minutes on ice. Following the bicinchoninic acid assay, about 40 μg of total protein from each lysate was separated using SDS–polyacrylamide gel electrophoresis (PAGE). Proteins were then transferred to a PVDF membrane. Each membrane was then blocked overnight at 4°C with 5% nonfat dry milk in TBST (20 mM Tris-HCl, pH 7.6, 150 mM NaCl, 0.05% Tween-20), incubated with a primary antibody for 1 hour at room temperature in 5% nonfat dry milk/TBST, washed thrice with TBST, and incubated with a horseradish peroxidase-linked secondary antibody in 5% nonfat dry milk/TBST for 1 hour at room temperature. After washing with TBST, protein bands were visualized on X-ray film using 3,3′,5,5′-tetramethylbenzidine staining. The developed blots were subjected to densitometric analysis by ImageJ 1.43 software (NIH, Bethesda, MD) using β-actin as an internal control.
In vivo antitumor effect
MCF-7 cells (2×106) were resuspended in 100 μL of PBS and intradermally injected into the back of the mice. When the tumor was 4–6 mm in diameter, mice were treated with intratumor injections of UV-Tianjin (2.5×108 particles in a total volume of 100 μL) or 100 μL of saline (control). The injections were administered on days 10, 13, and 16. Tumor volumes were measured in a blinded manner using slide calipers and the following formula: tumor volume (mm3)=length×(width)2/2. Mice were sacrificed 21 days post-treatment, and tumors were removed and weighed.
TUNEL assay
Tumor tissues from mice treated with UV-Tianjin or saline (n=10/group) were fixed in 10% neutral buffered formalin in PBS and embedded in paraffin. Apoptotic cells were identified using a TUNEL apoptosis assay kit. Three discontinuous sections were selected from every specimen. Images of the sections were obtained using a fluorescence microscope, and apoptotic cells had dark brown nuclei when viewed with a light microscope. The number of apoptotic cells per section was counted in 10 random high-power microscopic fields (400×). Researchers performed these counts in a blinded manner. The apoptotic index was calculated by dividing the number of TUNEL-positive cells by the total number of cells. 21
Immunohistochemical analysis
The immunohistochemical analyses were performed on the different sections of the same tumors with TUNEL assay. Thin sections were stained with specific antibodies in combination with secondary antibodies. A cell was considered positive if it had brown granules in the cytoplasm. The number of positive cells was counted in 10 microscopic fields (400×) that were chosen at random. The percentage of positive cells (positive cells/total cells×100%) was then calculated. Three discontinuous sections from each specimen were analyzed, and the three resulting indexes were averaged.
Statistical analyses
Data represent the mean±standard deviation from at least three independent experiments. Statistical analyses were performed using SPSS 18.0 for Windows (SPSS, Chicago, IL). Statistical comparisons were performed using one-way analysis of variance, and further post hoc testing was conducted using Prism 5 software (GraphPad, San Diego, CA). A p-value <0.05 was considered statistically significant.
Results
UV-Tianjin inhibits the growth of human breast cancer cells
When the human breast cancer cell lines MCF-7, MDA-MB-231, and T47D were treated with UV-Tianjin at various MOI for 24 hours, cell viability was suppressed in a dose-dependent manner (Fig. 1A). However, susceptibility to UV-Tianjin treatment varied, as MCF-7 cells, which are caspase-3 deficient, were most sensitive. A microscopic examination revealed that, for cells treated with UV-Tianjin at MOI 40, MCF-7 cells had more extensive cell fusions than MDA-MB-231 or T47D cells (Fig. 1B).
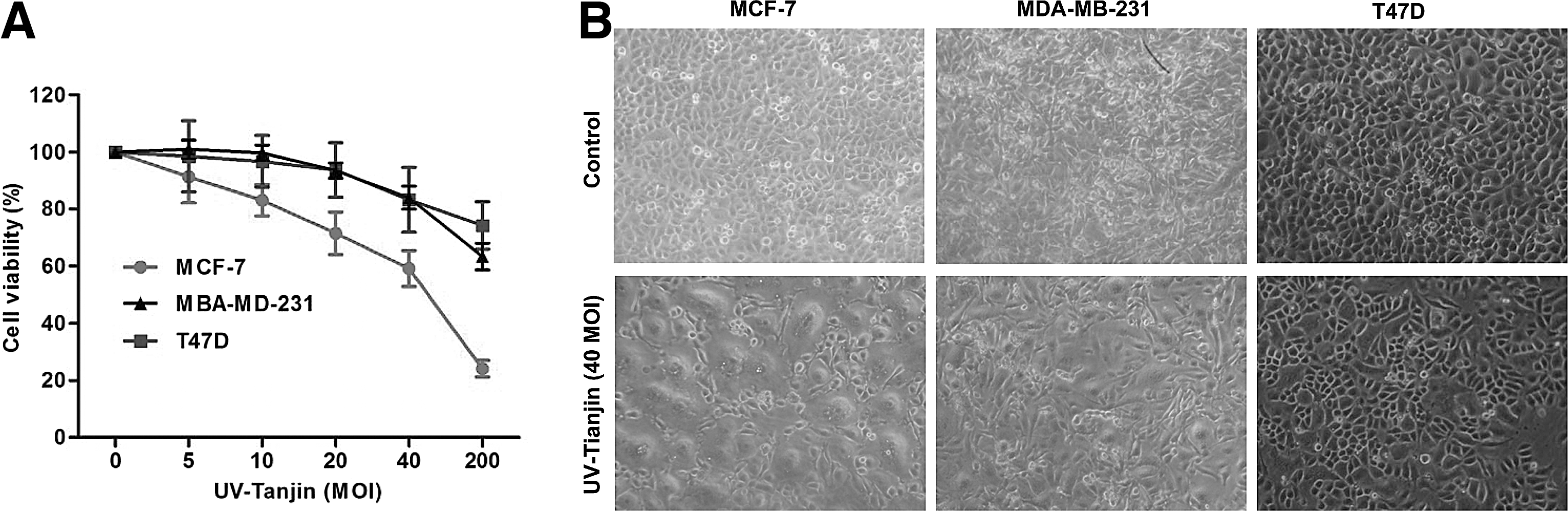
Effect of UV-Tianjin on the viability of MCF-7, MDA-MB-231, and T47D cells.
UV-Tianjin induces apoptosis of MCF-7 cells
Given that MCF-7 cells were most sensitive to UV-Tianjin treatment, the authors focused their analysis on this cell line. To determine whether UV-Tianjin-mediated growth inhibition resulted from apoptosis, they stained MCF-7 cells with Hoechst 33342. Following UV-Tianjin treatment, nuclei were often condensed, which indicates apoptosis. Moreover, there was a dose-dependent increase in apoptosis when MCF-7 cells were treated with UV-Tianjin (Fig. 2A). The authors next used flow cytometry to estimate the number of apoptotic cells resulting from UV-Tianjin treatment (24 hours). UV-Tianjin MOIs of 10, 20, and 40 resulted in 37.3%±5.1%, 43.6%±3.2%, and 58.6%±4.3% apoptotic cells (early apoptosis+late apoptosis), respectively. A significant difference in the number of apoptotic cells was observed between treated and untreated MCF-7 cells [p<0.01; Fig. 2B(a–d), C]. Taken together, these results showed that UV-Tianjin effectively initiated apoptosis in MCF-7 cells.
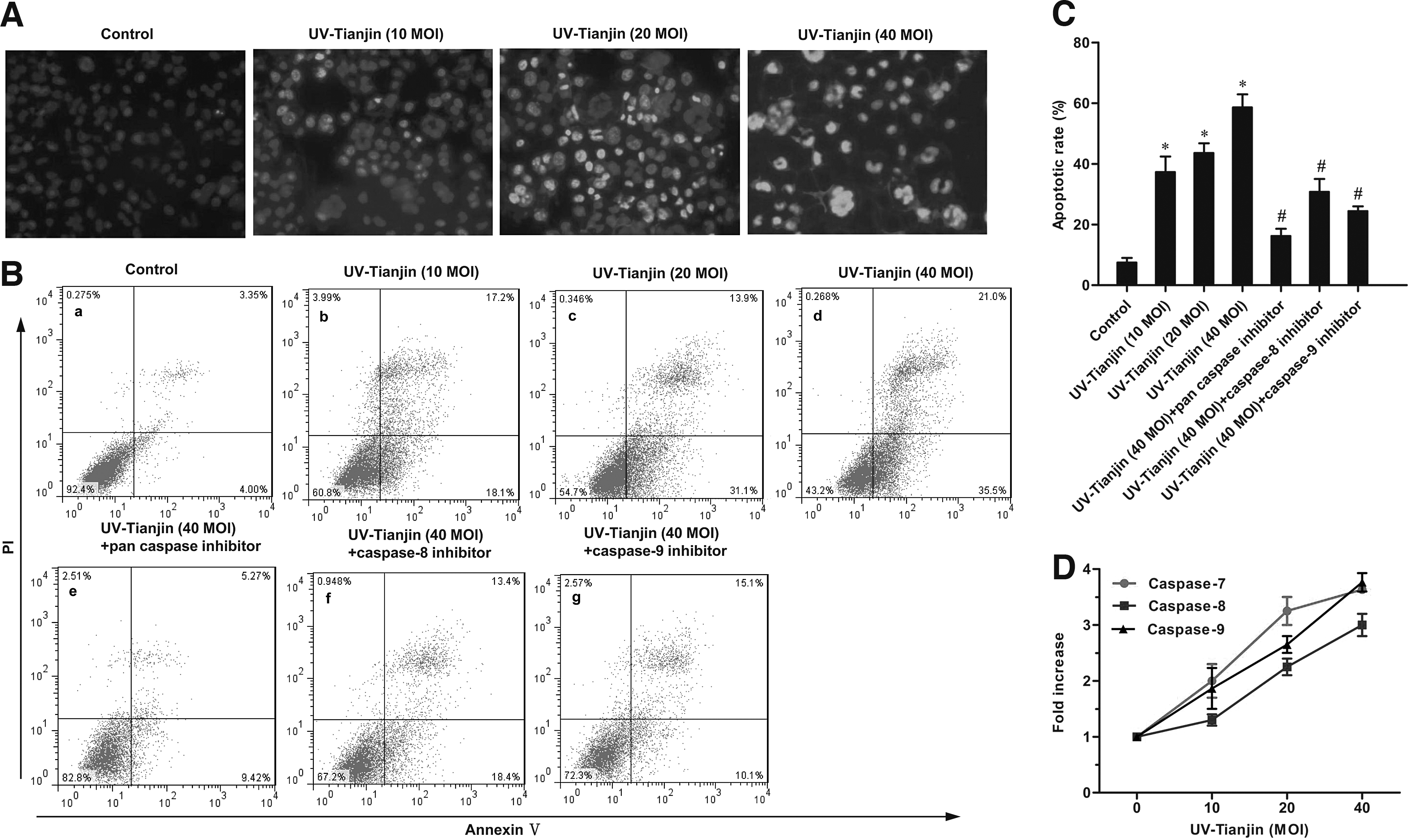
UV-Tianjin induces apoptosis of MCF-7 cells.
UV-Tianjin-induced apoptosis is caspase dependent in MCF-7 cells
To investigate the mechanisms by which UV-Tianjin induced apoptosis of MCF-7 cells, the authors used colorimetric methods to analyze caspase-7, caspase-8, and caspase-9 activities. Activities associated with all three caspases increased in a dose-dependent manner following 24 hours of UV-Tianjin treatment (Fig. 2D). To determine which caspase was most relevant to UV-Tianjin-induced apoptosis, pharmacological inhibitors of caspases were added to the cultures. When cells were pretreated with Z-VAD-FMK (a pan-caspase inhibitor), Z-LEHD-FMK (a caspase-9-specific inhibitor), or Z-IETD-FMK (a caspase-8-specific inhibitor) for 2 hours, UV-Tianjin-induced apoptosis was inhibited to different degrees [Fig. 2B(e–g)]. The caspase-8 and caspase-9 proteases are essential components of the extrinsic and intrinsic apoptotic pathways, respectively, whereas caspase-7 functions as a downstream effector of both caspase-8 and caspase-9. As such, these data suggested that UV-Tianjin-induced apoptosis is caspase dependent and may involve both the endogenous mitochondrial pathway and the exogenous death receptor pathway.
Both the mitochondrial and death receptor pathways play a role in UV-Tianjin-induced apoptosis of MCF-7 cells
To investigate whether the mitochondrial apoptotic pathway was involved in UV-Tianjin-induced apoptosis, the dye JC-10 was used to examine changes in the MMP. CCCP, which disrupts MMP, was used as a positive control. MCF-7 cells were treated with different doses of UV-Tianjin (MOI: 5, 10, or 20) for 24 hours or 50 μM CCCP for 1 hour. Flow cytometry data suggested that UV-Tianjin disrupted the MMP (Fig. 3A, B). In this study, the level of MMP at 40 MOI is similar to that at 20 MOI. Hence, the lower MOI (5 MOI) was used. The fact that UV-Tianjin downregulated the MMP in a dose-dependent manner indicated that mitochondria played an important role in UV-Tianjin-induced apoptosis.
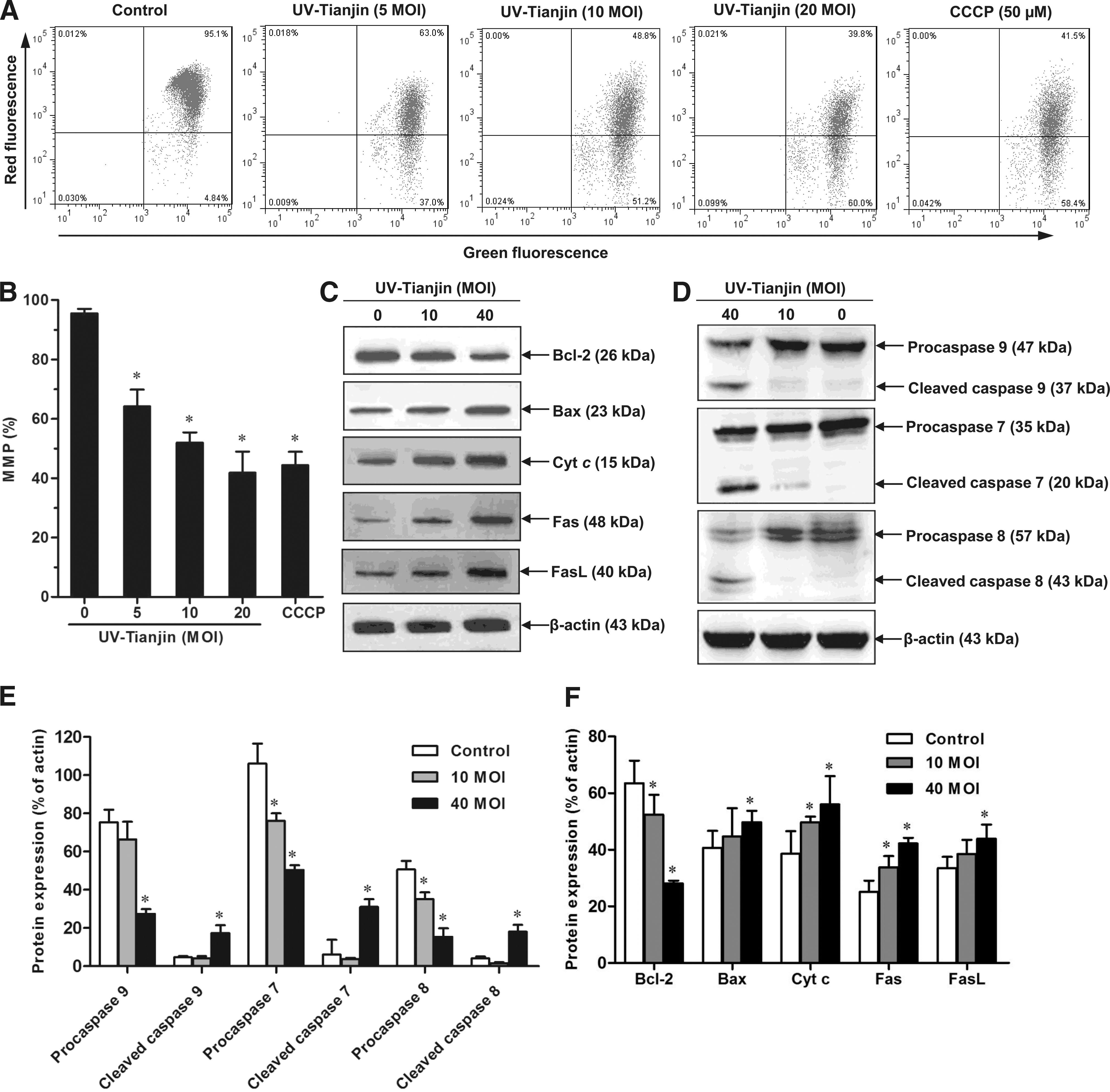
Effects of UV-Tianjin on the mitochondria membrane potential (MMP) and the expression of apoptosis-related proteins.
The authors next examined cytoplasmic levels of Bax, Bcl-2, cyt c, caspase-9, and caspase-7 with western blotting. As the dose of UV-Tianjin was increased, Bcl-2 was downregulated, whereas levels of Bax, cyt c, cleaved caspase-9, and cleaved caspase-7 were upregulated (Fig. 3C–F). Therefore, UV-Tianjin affected the MMP, which promoted changes in Bax and Bcl-2 levels and the release of cyt c. This activated caspase-9, which subsequently activated caspase-7 and led to the apoptosis of MCF-7 cells.
To determine whether the death receptor pathway was involved in UV-Tianjin-induced apoptosis, the authors next examined the effect of UV-Tianjin on Fas, Fas ligand (FasL), and the downstream caspase-8. Cells exposed to UV-Tianjin at MOI 40 exhibited increases in Fas, FasL, and cleaved caspase-8 compared with controls and cells treated with MOI 10 of UV-Tianjin (Fig. 3C–F).
UV-Tianjin inhibits tumor growth in vivo by inducing apoptosis
The effect of UV-Tianjin on tumor growth was further analyzed in tumor-bearing mice. Consistent with these in vitro studies, intratumor injection of UV-Tianjin inhibited MCF-7 tumor growth in vivo (p<0.01; Fig. 4A, B). At 21 days post-treatment, the average weight of UV-Tianjin-treated tumors was significantly less than tumors treated with saline (p<0.01; Fig. 4C).
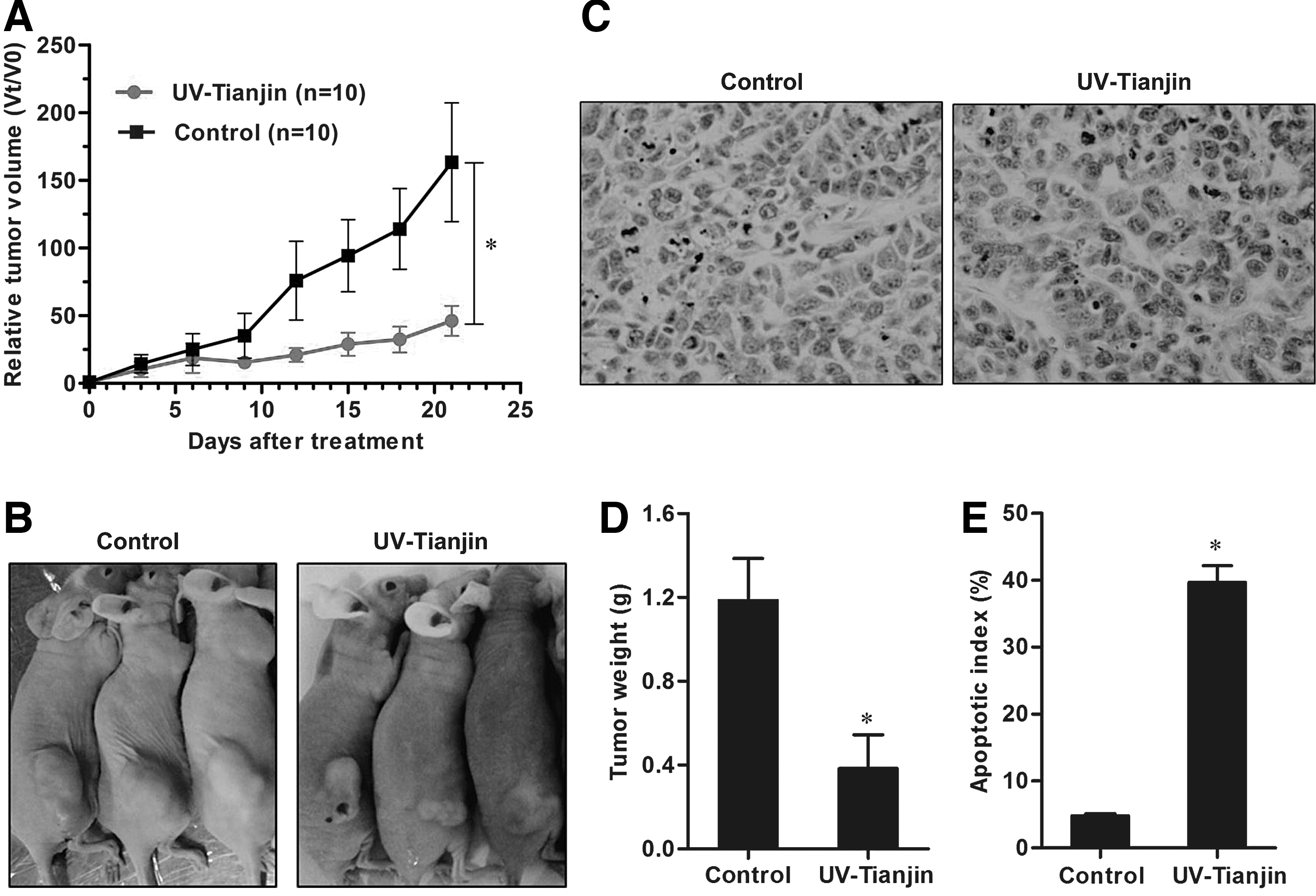
Antitumor effects of UV-Tianjin on breast cancer cell in vivo.
To determine whether UV-Tianjin elicited apoptosis in vivo, in situ TUNEL staining was performed on sectioned tumors. UV-Tianjin-treated tumors had significantly more apoptotic cells, that is, cells with dark brown nuclei, than control tumors (Fig. 4D). The apoptotic index of the UV-Tianjin-treated group was significantly higher than the control (p<0.01; Fig. 4E). These results further suggested that increases in tumor cell apoptosis may explain why UV-Tianjin had a strong antitumor effect in a model of human breast cancer.
UV-Tianjin activates caspase-7, caspase-8, and caspase-9 in tumor tissues
Immunohistochemical analyses were performed to assess caspase-7, caspase-8, and caspase-9 activation in tumors treated with UV-Tianjin or saline. The authors observed strong activation of these caspases in UV-Tianjin-treated tumors (Fig. 5A). In contrast, minimal caspase activation was observed in control mice (Fig. 5B). These results confirmed that UV-Tianjin induced apoptotic pathways mediated by caspase-7, caspase-8, and caspase-9.
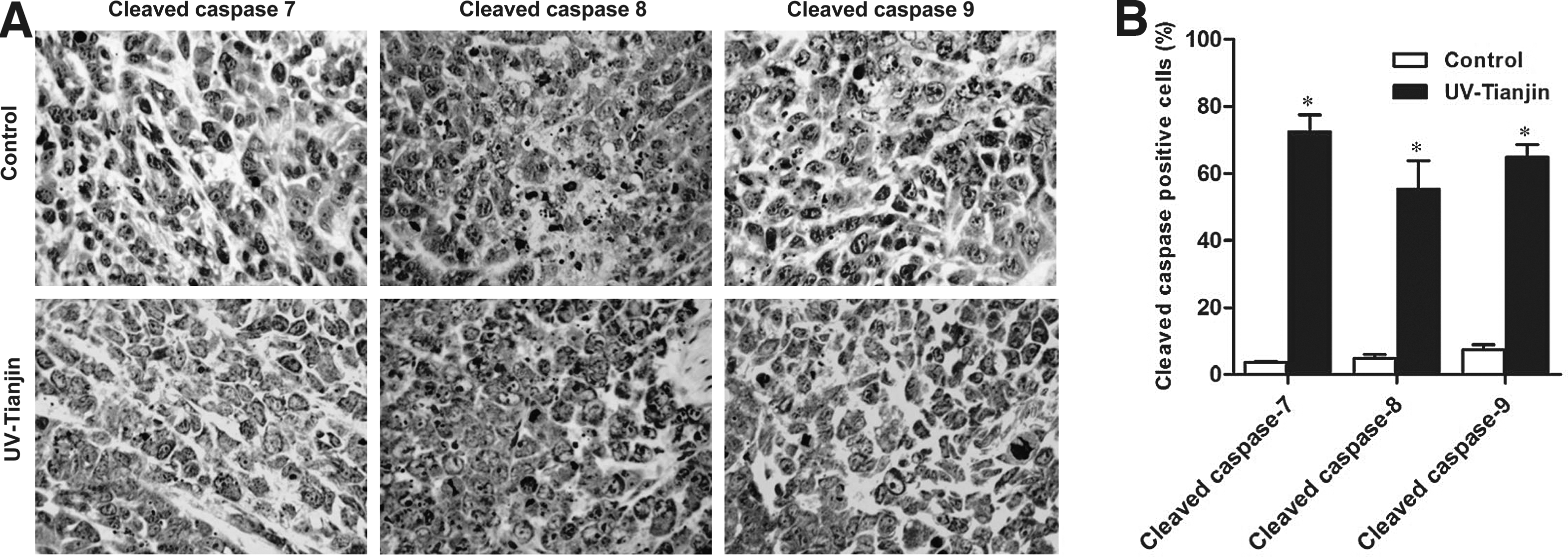
Analysis of caspase-7, caspase-8, and caspase-9 activation in tumor tissues using immunohistochemistry.
Discussion
Apoptosis is a common phenomenon in tumor cells playing an important role in both the genesis and progression of tumors. 22 Attempts to develop new antitumor drugs have recently focused on the induction of apoptosis to inhibit tumor growth. Previous studies have demonstrated that certain viruses inhibit tumor cell proliferation by inducing apoptosis. 23 –25 In this study, the authors demonstrated that UV-Tianjin inhibited the growth of the human breast cancer cell lines MCF-7, MBA-MD-231, and T47D. MCF-7 cells, which are caspase-3 deficient, were most sensitive to UV-Tianjin treatment. Hoechst staining and flow cytometric analysis revealed that UV-Tianjin induced MCF-7 apoptosis in a dose-dependent manner in vitro. In vivo, UV-Tianjin inhibited the growth of xenograft tumors in nude mice and significantly increased the number of TUNEL-positive tumor cells compared with controls. Thus, UV-Tianjin induced apoptosis of breast cancer MCF-7 cells in vitro and in vivo.
There are two major signaling pathways that mediate apoptosis: the extrinsic pathway, which involves death receptors, and the intrinsic pathway, which involves mitochondria. In this study, it is demonstrated that UV-Tianjin treatment decreased the MMP in a dose-dependent manner. Bcl-2 and related proteins are key regulators of the mitochondrial apoptotic pathway. 26,27 Bcl-2 binds to the outer membrane of mitochondria and inhibits apoptosis by preventing the release of cyt c. The proapoptotic protein Bax permeabilizes the mitochondrial membrane in response to cellular stress. 28 This triggers the release of cyt c into the cytosol, which associates with apoptotic peptidase activating factor 1 to form a caspase-9-activating protein complex called the apoptosome. Caspase-9 then activates downstream effector caspases, including caspase-3 and caspase-7, which cleave a number of cytoskeletal and nuclear proteins, such as poly (ADP-ribose) polymerase 1, and ultimately cause cell death. 29 In addition, caspase-7 is involved in the apoptosis of caspase-3-deficient MCF-7 cells. 30 In this study, the authors examined the effect of UV-Tianjin on Bax, Bcl-2, cyt c, caspase-7, and caspase-9. UV-Tianjin increased levels of Bax and cytoplasmic cyt c but downregulated Bcl-2. Moreover, UV-Tianjin treatment (MOI 40) induced the cleavage of procaspase-9 and procaspase-7 in MCF-7 cells. These results indicated that UV-Tianjin-induced apoptosis of MCF-7 cells was closely associated with the mitochondrial cell death pathway.
To determine whether the death receptor pathway was also involved in UV-Tianjin-induced apoptosis, the authors measured enzymatic activities of Fas, FasL, and caspase-8 with western blotting. Fas is a member of the tumor necrosis factor receptor superfamily and generates an apoptotic signal when bound by FasL, which localizes to the surface of adjacent cells. Once FasL binds Fas, Fas associated through death domain is then recruited to form the death-inducing signaling complex; formation of this complex leads to activation of the initiator caspase-8, which cleaves and activates effector caspases, such as caspase-3 or caspase-7, causing cell death. 31 These results showed that levels of Fas, FasL, and cleaved caspase-8 increased following UV-Tianjin treatment at MOI 40. Thus, the authors concluded that UV-Tianjin induced the apoptosis of MCF-7 cells through both extrinsic and intrinsic apoptotic pathways. More research is required to confirm these results and this research will provide a reference for biotherapy of breast cancer.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (No. 81172168).
Disclosure Statement
The authors declare that they have no competing interests.