
Abstract
The idea of multiple differentiation capacity of glioma stem cells and progentior cells (GSCPs) has been accepted by most of the researchers, but the effect of local environment on the differentiation of GSCPs is unclear. GSCPs SU2 and CM-Dil-stained C6 cells (C6-Dil) were injected into the brain of GFP transgenic nude mice. The xenografts were sectioned. Morphological changes of tumor cells that resided in the choroid plexus, molecular markers expression, and the relationship between the original tumor cells and host cells were studied carefully. The tumorigenicity rate was 40/40 (100%) in all of the inoculated nude mice. Cell morphology and molecular expression of neoplasm settled in the choroid plexus showed that choroidcarcinoma derived from GSCPs was developed. These results showed that GSCPs may have the multiple differentiation capacity, which can be induced by the local environment of host brain as NSCs, and cell fusion may play an important role in the transformation.
Introduction
The concept of cancer stem cells (CSCs) has been around for many years, 1,2 although little attention was paid to them at the beginning. Now, there are compelling evidences that CSCs do exist in most malignant tumors, including gliomas. Targeting on CSCs in glioma is very important in curing malignant glioma, because glioma stem cells and progenitor cells (GSCPs) may be the seed for tumor initiation and progression, and target therapy against these cells may hopefully eradicate glioma. 3 –5 However, the anti-CSC therapies are still unpractical at present, partly because of the insufficient investigation of the differentiation features of GSCPs.
Against this backdrop, it is of importance to investigate the features of GSCPs further. As known, GSCPs were called neural stem-like cells because they both share the same marker, CD1336; later, Singh reported that CD133+ glioma cells other than CD133− glioma cells were the only tumor initiation cells in glioma and were called GSCPs. 7 However, the authors have reported that CD133− cells differentiated from CD133+ cells could retrodifferentiate to CD133+ cells, 8 and the daughter cells of CD133− cells in the C6 cell line could be a mixture of these two kinds of cells. 9 Moreover, researchers have confirmed that a small subset of CD133− glioblastoma cells can undergo asymmetrical cell divisions, differentiate, and maintain infinite growth potential as well. Accordingly, these cells were called CD133− GSCPs, specific for a novel subtype of CSCs. 10
The different reports mentioned above may be due to the different derivations of experimental specimens and conditions in different laboratories, but it was deduced that the various characteristics of GSCPs may mainly depend on their multidirectional differentiation potential and the direction of differentiation is decided by the surrounding microecosystem. 11 The authors called this phenomenon as differentiation mimicry of GSCPs, which is similar to that of NSCs. In this study, they injected GSCPs into the brain of nude mice and investigated the mimicry growth characteristics of GSCPs when they settled in different tissues of host brain. It is very interesting that GSCPs formed tumor in the choroid plexus, because as far as one knows, none such case was reported before. The mimicry growth characteristics of GSCPs reflect some unusual features of GSCPs, which will be discussed in the text.
Materials and Methods
Reagents and equipments
The mouse anti-Nestin monoclonal antibody and mouse anti-GFAP monoclonal antibody (BD biosciences Co.); mouse anti-Ki67 antibody, mouse anti-Cytokeratin monoclonal antibody, mouse anti-S100 antibody, and mouse anti-Vimentin antibody (Santa Cruz, CA); Micro-drill 18000-17 (Fine Science Tools) and stereotaxic instrument (HuaiBei Zhenghua bio-instruments Co.).
Cell culture
This study was approved by the Medical Review Board of Soochow University Medical School. C6 cells were purchased from the Shanghai Cell Bank. GSCPs SU212 was established by this laboratory. C6 cells were cultured in 10% fetal bovine serum and SU2 cells were cultured as described previously. 12 In brief, the cells were cultured in serum-free DMEM/F-12 containing human recombinant N2 (20 ng/mL; Invitrogen), EGF (20 ng/mL; Invitrogen), bFGF (20 ng/mL; Gibco), and plated at a density of 3×106 live cells. Cells were fed by changing half of the medium every 3 days and were enzymatically dissociated into single cells for passage when they were large in size and high in density.
Animals
Four- to six-week-old male and female GFP nude mice at an average weight of 25 g were provided by the Center for Experimental Animals, Soochow University (certificate No. SY X K (Su) 2007-0035). All the animals were bred and maintained in the Specific Pathogen Free Animal Care Facility, Nasal1000 grade. The National Institutes of Health guidelines for the care and use of laboratory animals were followed in all animal procedures.
C6 staining by CM-Dil
C6 cells were cultured as confluent monolayers and washed thrice with phosphate-buffered saline (PBS) (4 minutes each) before staining. A staining medium was prepared by adding 5 μL CM-Dil to 1 mL DMEM/F12 medium. Then, C6 cells were incubated with the staining medium for 30 minutes. Thirty minutes later, the staining medium was removed and cells were washed again with PBS thrice (4 minutes each). The stinging cells (C6-Dil) were collected and injected into the brain of GFP nude mice.
Orthotopic implantation
As previously described, 12 mice were anesthetized by intraperitoneal injection of 10% chloral hydrate (200 mg/kg). In 30 mice, burr holes were made at the right side of the cranium 2.5 mm away from the midline and 1 mm anterior to bregma. In another 10 mice, bore holes were made 1 mm away from the midline and 6 mm posterior to bregma. Spheres of C6-Dil cells or GSCPs were enzymatically dissociated into single cells and resuspended in serum-free DMEM/F12. Fifteen microliters of cell suspension (∼2×105 cells) was injected into the right caudate nucleus or cerebellum at a depth of 3.5 and 3 mm, respectively, over 10 minutes by using a 20-μL Hamilton syringe assisted with a stereotaxic apparatus.
Immunohistochemistry
Mice were sacrificed for morphological studies when they became cachectic. The xenogeneic graft glioma samples were fixed in 4% phosphate-buffered formaldehyde, processed into paraffin blocks, sectioned, and stained with hematoxylin and eosin (HE) by routine histopathological procedures. The overview of tumor mass and its relationship with adjacent host brain structures were observed with naked eyes or a low power lens.
For immunohistochemical analysis, the endogenous peroxidase was quenched with 0.3% H2O2 for 30 minutes. The sections were washed with PBS for 5 minutes and incubated in diluted normal blocking serum for 20 minutes (VECTASTAIN Elite ABC kit). The sections were washed thrice with PBS, 5 minutes for each, and followed by incubation with a primary antibody (Nestin 1:100, Ki67 1:100, Cytokeratin 1:100, GFAP 1:100, S100 1:100, and Vimentin 1:100) at room temperature for 3 hours.
After washing thrice with PBS, 5 minutes for each, the sections were incubated with a biotinylated secondary antibody for 0.5 hour (VECTASTAIN Elite ABC kit) and followed by three 5-minute washes with PBS. The sections were incubated with the VECTASTAIN Elite ABC reagent for 30 minutes and then washed thrice with PBS for 5 minutes. 3,3′-Diaminobenzidine (vector SK-4100), mixed with distilled water, buffer, and H2O2 solution was applied to brain sections until staining was optimal, as determined by light microscopy. Then, the sections were washed in tap water, dehydrated, cleared with xylene and coverslipped with neutral balsam mountant.
Immunofluorescence tracing of C6-Dil cells
C6-Dil cells were injected into the right caudate nucleus of GFP mice brain. Mouse brain tissues were fixed with 4% phosphate-buffered paraformaldehyde, dehydrated with 20% sucrose solution, and cut into 5 μm coronal sections with a cryostat. Sections were washed thrice with PBS, mounted with the Vectashield medium (Vector laboratories), and coverslipped. Results were viewed using a confocal laser scanning microscope (Leica TCS SP2).
Cell counting
To determine the percentage of positive labeling cells, the authors first counted the number of tumor cells of three different areas randomly under a microscope (400×) and then got an average number for every area. The same method was applied to get an average number of positive cells (Nestin positive, GFAP positive, Ki-67 positive, or Cytokeratin positive). The final number of positive cells was expressed as a percentage of total tumor cells. An investigator who was blinded to the experimental conditions performed cell counting.
Statistical analysis
For comparison of multiple means, analysis of variance followed by the Student–Newman–Keuls analysis was performed. All values are expressed as mean±SE. Statistical significance (p<0.05 for all analyses) was assessed using GraphPad Prism 4 software.
Results
Morphological differences of tumors derived from GSCPs homing at different sites
GCSPs were injected into different anatomic sites of nude mice. HE staining of xenograft showed that tumor cells derived from GSCPs rapidly grew in the injected site at the beginning and then invaded diffusely into the surrounding structures, including the lateral ventricle, third ventricle, subarachnoid space, parenchyma of the contralateral cerebral hemisphere, and even the leptomeningeal.
It is rather intriguing that the morphology of tumor cells varied from site to site. Tumor cells proliferated easily after injection into the caudate nucleus of mouse brain. They were fusiform or asteroid and manifested a combination of extensive cytoplastic and nuclear pleomorphism. They arranged tightly into a small cluster and disseminated broadly, consistent with the morphosis of glioblastoma (Fig. 1E, F). A solid tumor in the choroid plexus had marked nuclear pleomorphism, increased cellular density, and blurring of mammillae with poorly structured sheets of tumor cells, pathologically similar to choroid plexus carcinoma (Fig. 1G, H). Tumor along the ventricle wall was pathologically similar to the ependymal tumor, and tumor cells there had atypical pseudorosettes in which obvious nuclear pleomorphism and megakaryocytes were observed clearly (Fig. 1I, G). Tumor cells that invaded into leptomeningeal were surrounded by a bulk of rhabdoid cells, analogic to the morphological structure of meningioma (Fig. 1K, L). The results indicated that the morphology of tumor derived from GSCPs could be affected by the microecosystem around which they grow in.

Differentiation mimicry of CSCs in different parasitic location.
Migration of GSCPs and morphological evidence of choroid plexus neoplasm formation
The migration pathway of GSCPs in the progression of choroid plexus neoplasm formation was delineated according to the mouse stereotaxic atlas. 13 The results showed a part of GSCPs injected into the caudate nucleus first invaded into the CA area of hippocampus and then disseminated to choroid plexus of the third ventricle (Figs. 1C and 2A–D); sometimes, however, GSCPs could directly invade into the lateral ventricle and grow along the choroid plexus there (Figs. 1D and 2E–H); GSCPs were also injected into the cerebellum to see whether they could invade into the choroid plexus of the fourth ventricle and form a tumor there. As expected, choroid plexus neoplasms were seen in the fourth ventricle (Fig. 2L, M).

Tumor evolution in choroid plexus after invasion of CSCs.
Morphologically, there were two kinds of tumors formed in the choroid plexus, the gigantic type that occupied the whole ventricle and the hydrocephalus type that filled with overflowed cerebrospinal fluid (CSF) (Fig. 2E–H). Hydrocephalus formation in the latter may be due to the CSF overdose secretion or blockage of the CSF pathway by a tumor mass. In addition, the authors found that GSCPs in choroid plexus occupied the domain of host cells step by step. For example, the transition region between cancer and noncancer could be seen in the early stage (Fig. 2I–M); however, at the advanced stage, the mouse choroid plexus was completely replaced and formed by tumor cells (Fig. 2E–H). These results demonstrated the role of tissue remolding of GSCPs in host tissues, but the underlying mechanism is still waiting to unveil.
To further investigate the interaction between GSCPs and host cells, C6 cells, which were mostly glioma stem and progenitor cells, 9 were stained by CM-Dil. C6-Dil cells were injected into the brain of GFP transgenic nude mouse bred in our own laboratory. 14 As expected, choroidal tumor was formed as SU2 did. The results showed that there were at least three kinds of cells seen in the choroidal tumor, including C6-Dil cells (red), the host cells (green), and the fusion cells (yellow) (Fig. 3). These data suggested that host cell-like tumor cells might be derived from the fusion cells of the original tumor cells and the host cells.

Choroid plexus tumor developed from C6-Dil cells after injection of these cells into the brain of GFP transgenic nude mouse.
Markers expression of choroid plexus neoplasms
To investigate the biological phenotype of tumor cells, the expression of cell surface markers, including Nestin, Ki67, GFAP, S100, and Vimentin, was detected. Compared to the normal monolayer choroid plexus cells of normal mice brain, the cells of choroid plexus in tumor-bearing mice were in disorder and multilayered with obvious nuclear pleomorphism. Furthermore, the tumor cells strongly expressed Nestin and Ki67, while normal choroid plexus cells did not express Nestin and expressed Ki67 moderately (Fig. 4A, B). Secreting CSF, choroid plexus cells are endowed with the potential to self-renew, and hence, it is reasonable to find Ki67-positive cells in normal choroid plexus. However, when CSCs settled down in choroid plexus, the number of Nestin-positive and Ki67-positive cells was increased dramatically. Various morphological changes and disordered arrangement of the cells in choroid plexus prompt the formation of tumor in the choroid plexus. Moreover, based on the separate criteria of WHO (2007), 15 Cytokeratins are expressed by most choroid plexus carcinomas, while GFAP is present in about 20% of cases (Fig. 4C, D). In the current study, tumor cells in the choroid plexus were positive for both Cytokeratins and GFAP, while normal cells were negative for these markers (Fig. 4C, D). In addition, tumor cells were negative for both S100 and Vimentin (Fig. 4E, F). Taken together, it was certain that tumors derived from GSCPs, growing in the choroid plexus, were choroidcarcinoma other than malignant glioma.
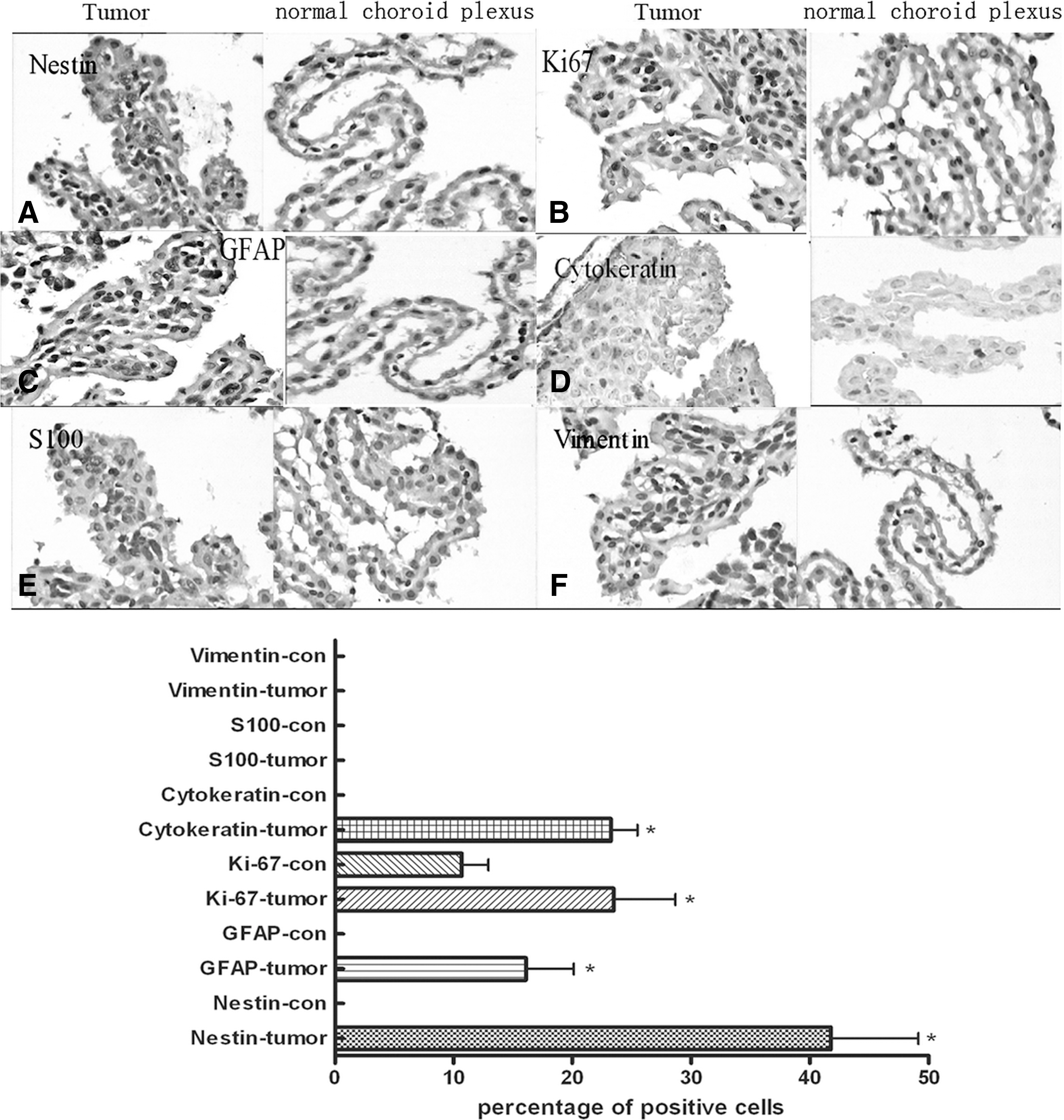
Immunohistochemistry staining of choroid plexus tumor (400×).
Discussion
Growing evidence has demonstrated that NSCs could migrate to a distant impaired area, settle down, and differentiate into corresponding cells of this area. This potential of NSCs is called differentiation mimicry. 16,17 GSCPs and NSCs are similar in many aspects, but whether GSCPs have the potential of differentiation mimicry is unclear. As mentioned previously, 12 GSCPs had the potentials of retrodifferentiation and widespread dissemination. In this study, the authors found that GSCPs spread widely to different regions after being injected into the mice caudate nucleus, and tumor cell morphology varied from site to site. For example, the appearance of tumor cells in leptomeningeal and choroid plexus was completely different from that of tumor cells in the caudate nucleus and gray matter. This study offered the preliminary evidence to support that the GSCPs could generate different tumor types, mostly depending on the local microecosystem where they resided. It seems that tumors from different areas could be divided into, at least, malignant astrocytoma, malignant ependymoma, malignant meningioma, and choroidcarcinoma. If tumor-bearing mice were induced by carcinogenic agents or gene modification, it could be sure that the animal had got polytypic tumors. 18 –20 This differentiation mimicry phenomenon was actually inconsistent and even contradicted with the popular idea that GSCPs are featured by replicating their parent tumor. However, the authors think that the differentiation mimicry maybe more correct to reflect the characteristics of the tumor stem cells, as it reflects both the plastic nature of stem cell and the impact of the microenvironment. It will be more acceptable if the related mechanism could be well elucidated in a further study.
Transdifferentiation of NSCs into vascular endothelial cells has been reported by Oishi, 21 and the same transdifferentiation of CSCs called vasculogenic mimicry may exist for the finding of tumor cell-derived endothelial cells. 22 Actually, both of these findings concerning transdifferentiation could be regarded as special differentiation mimicry of NSCs and CSCs when subjected to a vascular microecosystem. In contrast to the normal differentiation mimicry that occurred within the same embryonic layer, transdifferentiation of vasculogenic mimicry took place between the ectoderm and mesoblastema. Theoretically, transdifferentiation between different embryonic layers is far more complicated, but the differentiation mimicry of CSCs into choroid plexus tumor may be easier because it was a transformation between the same embryonic layers, and thus, the key point was whether the tumor derived from CSCs was really choroid plexus tumor. First, morphologically, as Figure 2 shows, choroid plexus tumor cells derived from SU2 or C6 had marked nuclear pleomorphism, increased cellular density, blurring of mammillae with poorly structured sheets of tumor cells, and pathologically similar to choroid plexus carcinoma. The GFP transgenic nude mouse provides the opportunity for us to investigate the relationship between tumor cells and host cells in vivo. C6-Dil cells, which were red seen under a fluorescence microscope, were injected into the brain of GFP transgenic nude mouse. The resulted tumor showed that in addition to the original tumor cells (red) and host cells (green), there were fusion cells of tumor cells and host cells, which suggested that GSCPs may obtain the ability of mimicry differentiation into host cell-like cells by cell fusion with the local host cells; second, immunohistochemically, as Figure 4 shows, tumor cells overexpressed Nestin and Ki67 indicating that the majority of tumor cells were immature and highly proliferative. In addition, tumor cells were positive for Cytokeratin and GFAP, both of which are special markers for choroid plexus tumors. Although the human choroid plexus tumor often expressed S100 and Vimentin, the authors did not find positive expressions of these markers in these transplanted models. These results were coincident with those in Shimomoto's report. 23 Based on the separate criteria of WHO (2007), 14 and taken together, all the results proved the differentiation of GSCPs to choroidcarcinoma.
The progression of cancerogenesis in choroid plexus will help us to further understand the differentiation mimicry of GSCPs. As Figures 2 and 3 shows, choroidcarcinoma formed gradually after the migration of GSCPs to the choroid plexus. However, it was still unclear whether this progression was different from that of other cancerogenesis such as the formation of spontaneous tumor, carcinogenic agent-induced tumor, and gene remolding-induced tumor. In this study, the authors found that GSCPs in the gray matter or near the blood vessels had the ability to fuse with the host cells (Fig. 3), but whether this fusion effect has impact on the differentiation of GSCPs needs to be observed in future investigations. Besides, it was a very complicated tissue remolding process for CSCs to differentiate to choroidcarcinoma, and cell fusion may be only one of the complicated mechanisms. To further investigate the morphodifferentiation mechanisms of differentiation mimicry of GSCPs, more studies should be focused on the microecosystem where GSCPs are located. As known, the destiny of NSCs is decided by the local microecosystem provided by their niche. 24 –26 For CSCs, their niche is aberrant with abnormal vessels 27 –29 and whether the niche plays the similar role of NSCs remained unclear. At least, in this test, the authors showed a formation of choroidcarcinoma after injecting of GSCPs into nude mouse brain in vivo. The underlying mechanism prompts that this process may be the cell fusion between GSCPs and the choroid plexus cell, which was induced by the local tumor microecosystem. The relationships between CSCs and their niche should be further investigated in the future.
Footnotes
Acknowledgments
This work was funded by grants from the Natural Science Foundation of China (No. 81302196; 81172400; 81071766; 81472739), the Natural Science Foundation of Jiangsu Province (No. BK2011341), and the Natural Science Foundation of Suzhou (No. SYS201307).
Disclosure Statement
No conflicting financial interests exist.