
Abstract
Cancer Biotherapy and Radiopharmaceuticals
officially retracts the article entitled, “Immature Colon Carcinoma Transcript 1 Is Essential for Prostate Cancer Cell Viability and Proliferation,” by Zhijun Wang, Danfeng Xu, Yi Gao, Yushan Liu, Jizhong Ren, Yacheng Yao, Lei Yin, Jie Chen, Sishun Gan, and Xingang Cui, (Cancer Biother Radiopharm 2015;30(7):278–284; doi: 10.1089/cbr.2014.1728) due to incorrectly attributed or identified sequences. Several figures and text within the methods section are strikingly similar to those in other publications1–3 and suggests that same party prepared these papers for entirely different author sets.
The Editor and Publisher of Cancer Biotherapy and Radiopharmaceuticals are committed to preserving the scientific literature and the community it serves.
References
1. Zhang C, Zhang T, Song E, et al. Ribosomal protein S15A augments human osteosarcoma xell proliferation in vitro. Cancer Biother Radiopharm 2018;29(10):451–456; doi: 10.1089/cbr.2014.1698.
2. Wang H, Wang B, Zhu W. et al. Lentivirus-mediated knockdown of myosin VI inhibits cell proliferation of breast cancer cell. Cancer Biother Radiopharm 2015;30(8):330–335; doi: 10.1089/cbr.2014.1759.
3. Zhao S, Chen X, Wan M, et al. Tectonic 1 is a key regulator of cell proliferation in pancreatic cancer. Cancer Biother Radiopharm 2016;31(1):7–13; doi: 10.1089/cbr.2014.1778.
Introduction
In developed countries, prostate cancer is the most universally diagnosed noncutaneous cancer in males and the second leading cause of cancer-related death (after lung cancer) for men. 1 According to the recent epidemiological reports, the incidence of prostate cancer in China is increasing fast, especially in some developed regions. 2 As known, the prostate is an androgen-dependent organ; androgen signaling should go hand-in-hand with prostate cancer development and progression. 3 –7 However, after about 12–24 months of androgen-deprivation therapy, almost all tumors will develop to castration-resistant prostate cancer (CRPC) with the distant metastases and a poor prognosis in the end. 8 Due to the fact that CRPC patients are usually androgen negative and have poor prognosis and survival ratio, CRPC was investigated frequently at the present time.
The immature colon carcinoma transcript 1 (ICT1) gene was initially found in transcriptomics research between undifferentiated and differentiated human colon carcinoma cells HT29-D4 and Caco-2. 9,10 Among many differentially expressed genes, its mRNA was markedly downregulated during in vitro differentiation of HT29-D4 and Caco-2 cells; thus, the gene was named ICT1. ICT1 also named as 39S ribosomal protein L58, mitochondrial (MRP-L58), and digestion substraction-1 (DS-1). ICT1 was recently confirmed as a mitochondrial ribosomal protein (MRP). 11 As is well known, mammalian mitochondria use oxidative phosphorylation (OXPHOS) to provide more than 90% of the adenosine triphosphate used for energy by cells. In mammals, the mitochondrial genome only encodes 13 essential proteins required for OXPHOS, which are synthesized on 55S ribosomes using the specialized translational system within the mitochondrion. Mammalian mitochondrial ribosomes (55S) consist of small (28S) and large (39S) subunits. 12 ICT1 is an essential peptidyl-tRNA hydrolase (PTH) component of the mitochondrial large ribosomal subunits and regarded as a codon-independent translation release factor, which has lost stop codon specificity and directs the termination of translation in the organelle, possibly in case of abortive elongation. 13 Thus, ICT1 may be involved in the hydrolysis of peptidyl-tRNAs that have been prematurely terminated and the recycling of stalled mitochondrial ribosomes. Handa et al. indicate that the ICT1 function is indispensable to cell vitality and mitochondrial activities. 14 A recent study has shown that ICT1 may take part in the progression of lung cancer and deemed as a potential biomarker. 15 However, the role of ICT1 in prostate cancer remains unclear.
To investigate the biological function of ICT1 in prostate cancer progression, the authors established lentivirus-mediated RNA interference (RNAi) to downregulate ICT1 expression in DU145 and PC-3 cell lines, which are androgen-independent prostate cancer cells. 8 Furthermore, the effects of ICT1 knockdown on cell proliferation, colony formation, and cell cycle progression were investigated.
Materials and Methods
Cell lines and cell culture
Human prostate cancer cell lines DU145 (Cat. No. TCHu222) and PC-3 (Cat. No. TCHu158) and human embryonic kidney cell line 293T (Cat. No. GNHu17) were obtained from the Cell Bank of Chinese Academy of Sciences. DU145 and PC-3 cell lines, used as cellular models for in vitro infection experiments, were cultured in Ham's/f-12 (Cat. No. 11765-054; Gibco) supplemented with 10% fetal bovine serum (FBS) (Cat. No. S1810; Biowest) and 1% non-essential amino acids. The 293T cell line, which was used to produce replication-incompetent lentiviral particles, was cultured in Dulbecco's modified Eagle's medium (Cat. No. SH30243.01B+; Hyclone) with 10% FBS. All cell lines were maintained at 37°C in a humidified 5% CO2 atmosphere.
Construction of ICT1 shRNA lentivirus vector and virus packaging
The following stem-loop-stem oligos were synthesized. The negative control short hairpin RNA (shRNA) was 5′-GCGGAGGGTTTGAAAGAATATCTCGAGAT ATTCTTTCAAACCCTCCGCTTTTTT-3′. Two ICT1-specific shRNA sequences were 5′-GCTGTTAATGCTTGTCTATAACTCGAGTTATAGACAAGCATTAACAG CTTTTTT-3′ (S1) targeting nucleotides 721–739 and 5′-GCAGAATGTGAACAAAGTGAACTCGAGTTCACTTTGTTCACATTCTGCTTTTTT-3′ (S2) targeting nucleotides 269–287, respectively. The oligos were annealed and ligated into the pFH-L vector (Shanghai Hollybio) digested by NheI and PacI. Recombinant pFH-L plasmids were determined by DNA sequencing. Lentiviruses were generated by triple transfection of 80% confluent 293T cells with the recombinant pFH-L plasmid and pVSVG-I and pCMV△R8.92 helper plasmids (Shanghai Hollybio) using Lipofectamine 2000. Culture supernatants were harvested 3 days after transfection to extract lentiviruses expressing ICT1 shRNA [Lv-shICT1(S1)/(S2)] or control shRNA (Lv-shCon). After that, the lentiviruses were purified through ultracentrifugation, and the titer of lentiviruses was measured. DU145 and PC-3 cells were infected with the concentrated lentiviruses at a multiplicity of infection of 40 and 45, respectively.
Reverse transcription-PCR and quantitative PCR
Total cellular RNA was extracted using the TRIzol® reagent (Cat. No. 15596-018; Invitrogen). The quantity and quality of RNA samples were measured by spectrophotometry and agarose gel electrophoresis, respectively. One microgram of extracted RNA samples was reverse transcribed into cDNA following the manufacturer's protocol (Cat. No. M1705; Promega). Reverse transcription real-time quantitative PCR (RT-qPCR) was carried out by the GXD kit iqSYBR Green (Cat. No. 1708882AP; BioRad) according to the manufacturer's recommendations in the CFX96 Touch™ Real-Time PCR Detection System (BioRad). The primers sequence and the polymerase chain reaction (PCR) products size were as follows: ICT1, forward 5′-CAGCCTGGACAAGCTCTACC-3′ and reverse 5′-GGAACCTGACTTCTGCCTTG-3′ (182 bp); actin, forward 5′-GTGGACATCC GCAAAGAC-3′ and reverse 5′-AAAGGGTGTAACGCAACTA-3′ (302 bp). The PCR protocol was as follows: initial denaturation at 95°C for 60 seconds, 40 cycles of denaturation at 95°C for 5 seconds, and annealing and extension at 60°C for 20 seconds.
Western blot
Cellular samples were harvested and lysed in an ice-cold lysis buffer (50 mM Tris, 2% SDS, 5% glycerinum, 100 mM NaCl, 1 mM EDTA, pH=6.8). Total protein concentrations of the cell lysate were quantified by the BCA Protein Assay Kit (Cat. No. 23235; Pierce Biotechnology). Twenty micrograms of cellular protein lysates was resolved on 15% SDS-PAGE and transferred to PVDF membranes (Cat. No. 162-0177; BioRad). The blots were probed with rabbit anti-ICT1 (Cat. No. AP20382b, 1:1000; Abgent), rabbit anti-CDK1 (Cat. No. 19532-1-AP, 1:1000; Proteintech), mouse anti-Cyclin B1 (Cat. No. K0128-3, 1:1000; Medical & Biological Laboratories Co. Ltd.), rabbit anti-Caspase 3 (Cat. No. 9661, 1:500; Cell Signaling Technology), rabbit anti-poly ADP-ribose polymerase (PARP) (Cat. No. 9542, 1:1000; Cell Signaling Technology), rabbit anti-Bax (Cat. No. 2774, 1:500; Cell Signaling Technology), rabbit anti-Bcl-2 (Cat. No. 2876, 1:500; Cell Signaling Technology), and rabbit anti-GAPDH (Cat. No. 10494-1-AP, 1:80000; Proteintech) and successively incubated with horseradish peroxidase (HRP)-conjugated goat anti-rabbit and goat anti-mouse IgG antibodies (Cat. Nos. SC-2054 and SC-2005, 1:5000; Santa Cruz). Signals were detected using the ECL-PLUS/Kit (Cat. No. RPN2132; Amersham). The blots were probed with GAPDH as control for loading. Bands on the X-ray films were quantified by Quantity One software (BioRad).
MTT assay
The effect of ICT1 on cell viability was determined based on cell growth curves using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) colorimetric assay as described previously. 16 Briefly, after 4 days of lentivirus infection, cells were seeded in 96-well plates at a density of 3×103 cells per well and allowed to attach overnight. Cell viability was evaluated daily within 5 days. At each indicated time `point, 10 μL of MTT solution (5 mg/mL in phosphate-buffered saline [PBS]) was added to the cells and incubated for 4 hours at 37°C. The medium was removed and 100 μL of solubilization solution (0.01 M HCl, 10% SDS, 5% isopropanol) was added to each well and incubated overnight at 37°C. The optical density of the plates was measured using the Epoch Microplate Spectrophotometer (Biotek) absorbance at 595 nm.
Colony formation assay
The effect of ICT1 on cell proliferation was determined based on colony numbers using the colony formation assay as described previously. 16 Briefly, after 4 days of lentivirus infection, DU145 cells were seeded in six-well plates at a density of 200 cells per well. The medium was replaced every 3 days. After 8 days of culture, cells were treated with PBS containing 4% paraformaldehyde for 30 minutes at room temperature. The fixed cells were stained with crystal violet (Cat. No. C0121; Beyotime) for 10 minutes, then gently washed with water, and air-dried. The number of colonies (>50 cells per colony) was counted under a microscope.
Cell cycle analysis by flow cytometry
Cell cycle distribution of DU145 cells was analyzed by flow cytometry with propidium iodide staining. Briefly, after 4 days of lentivirus infection, DU145 cells were seeded in 6-cm dishes at a density of 2×106 cells per dish. After 40 hours of culture, cells were washed thrice with PBS, trypsinized, and collected by centrifugation before resuspension in PBS containing 10% FBS. Precooled 70% ethanol was added dropwise to the cell suspensions. Samples were kept overnight at 4°C. Then, the cells were treated with 50 μg/mL propidium iodide and 50 μg/mL RNase at room temperature in the dark for 1 hour. Finally, the suspension was filtered through a 50-mm nylon mesh, and a total of 10,000 stained cells were subjected to flow cytometric analysis (FACSCalibur; BD Biosciences). Data were analyzed using the ModFit software (Verity Software House).
Apoptosis analysis by flow cytometry
Cell cycle distribution of DU145 cells was analyzed by flow cytometry with Annexin V-APC/7-AAD double staining. Briefly, after 4 days of lentivirus infection, DU145 cells were seeded in 6-cm dishes at a density of 1×106 cells per dish. After 48 hours of culture, the cells were collected and subjected to Annexin V-APC/7-AAD double staining according to the manufacturer's instruction (KeyGEN Biotech). Flow cytometry analysis was performed as mentioned above.
Statistical analysis
All experiments were repeated in triplicate at least, and the results from separate experiments are presented as mean±standard deviation (SD). Significance of the differences was determined by Student's t test. The p<0.05 was defined as a significant difference.
Results
Lentivirus-mediated shRNA suppressed ICT1 expression in DU145 and PC-3 cells
To explore the role of ICT1 in human prostate cancer, lentivirus-mediated shRNA was used to silence ICT1 expression. The concentration dose of Lv-shICT1 used was 1E+10 TU/mL. As shown in Figure 1A and D, after 4 days of lentivirus infection, over 90% and 80% of DU145 cells were green fluorescent protein positive in Lv-shICT1(S1) and Lv-shICT1(S2) groups, respectively, under fluorescence microscopy, indicating a successful infection efficiency. To further evaluate the knockdown efficiency, RT-qPCR and western blot assays were performed to detect ICT1 expression in DU145 cells. As shown in Figure 1B and E, the mRNA level of ICT1 was significantly reduced by 97.1% in the Lv-shICT1(S1) group and 82.9% in the Lv-shICT1(S2) group, respectively. However, no knockdown effect was observed following Lv-shCon infection. Moreover, the protein level of ICT1 was decreased by Lv-shICT1(S1) and Lv-shICT1(S2), respectively (Fig. 1C and F). In addition, the expression of ICT1 was obviously decreased in PC-3 cells after Lv-shICT1(S1) infection (Supplementary Fig. S1A and B; Supplementary Data are available online at
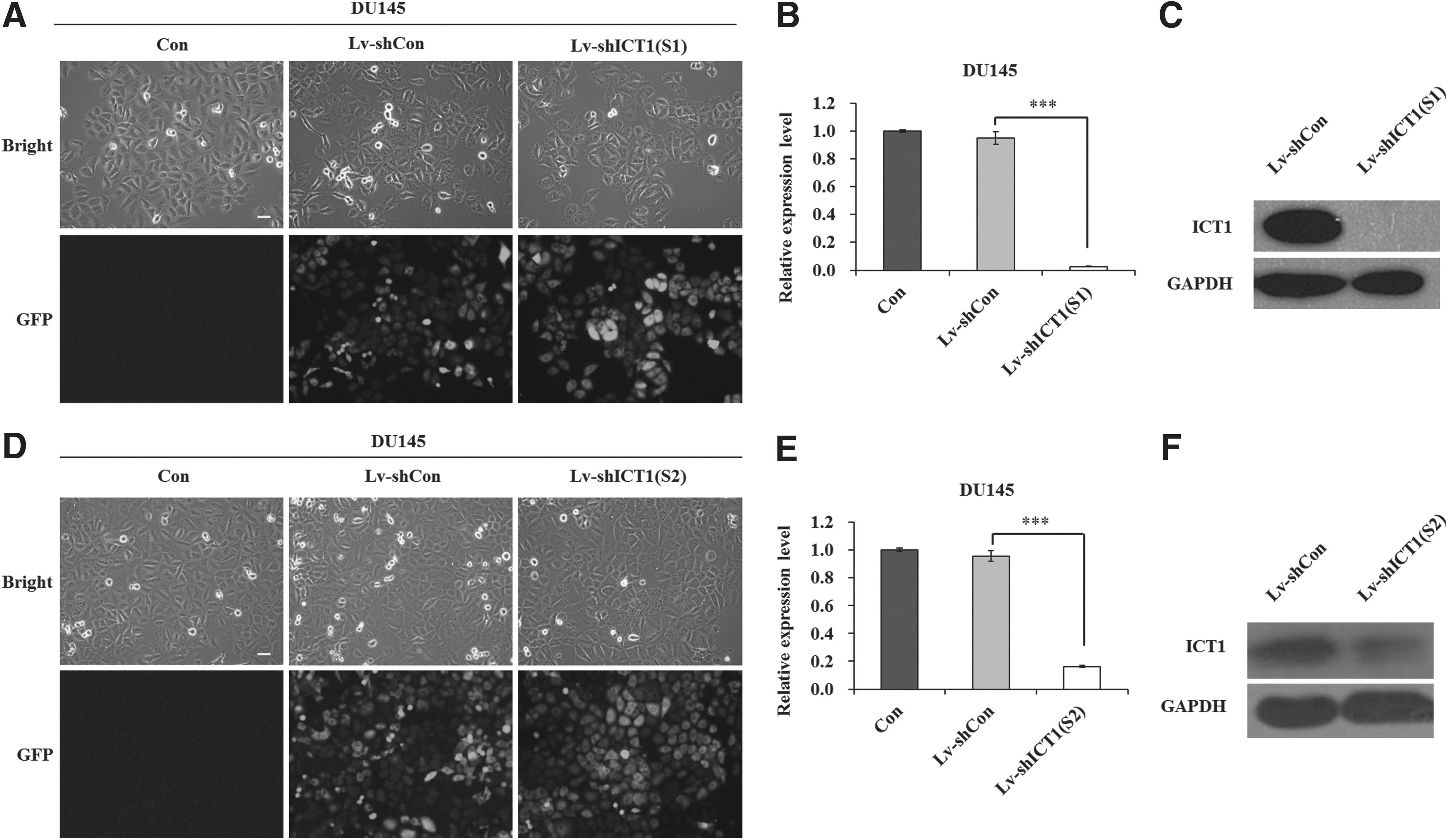
Silencing of immature colon carcinoma transcript 1 (ICT1) expression in DU145 cells by Lv-shICT1(S1)/(S2).
Knockdown of ICT1 suppressed viability of DU145 and PC-3 cells
To ask whether ICT1 plays a role in cell viability, the MTT assay was performed in DU145 and PC-3 cells. As shown in Figure 2A and B, the number of viable cells was much fewer in both Lv-shICT1(S1) and Lv-shICT1(S2) groups (p<0.001) than in Lv-shCon and Con groups. Moreover, the growth curve of PC-3 cells was also markedly dropped in response to ICT1 knockdown (Supplementary Fig. S1C). These results indicated that knockdown of ICT1 suppressed viability of DU145 and PC-3 cells.
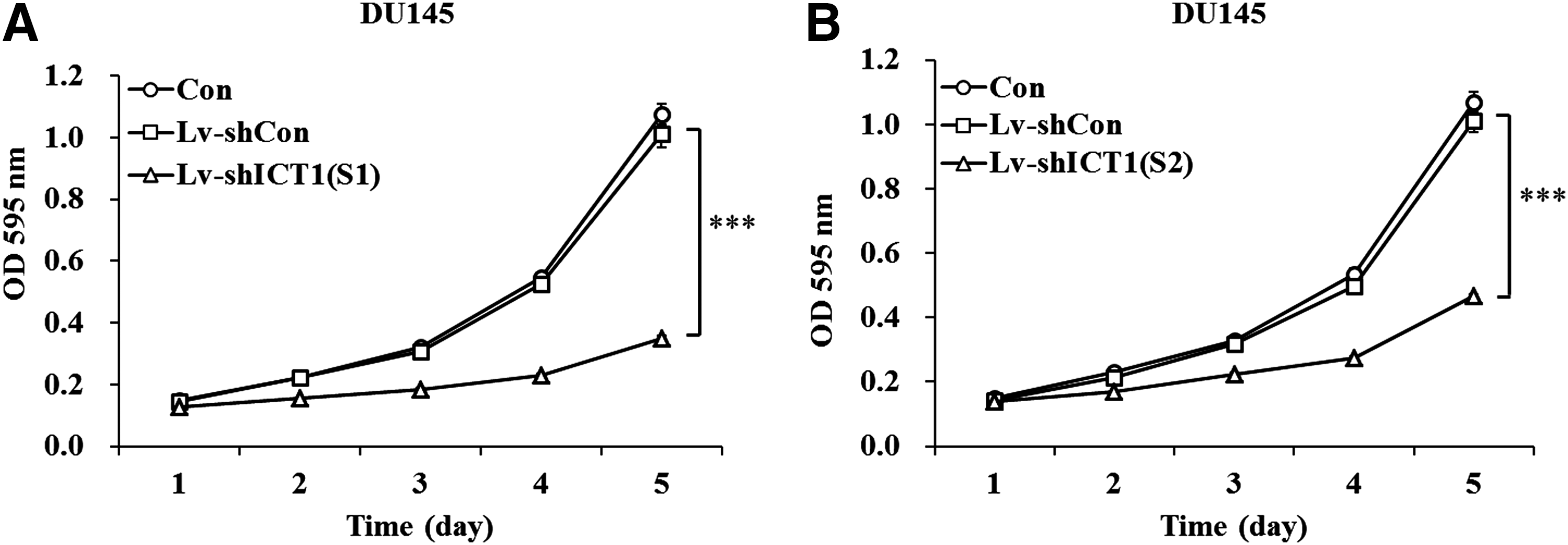
Lv-shICT1(S1)/(S2) inhibited the viability of DU145 cells.
Knockdown of ICT1 impaired colony formation in DU145 cells
Colony formation assay was performed to gain an additional insight into the effect of ICT1 on cell proliferation. As shown in Figure 3A and B, the number of colonies was much fewer in both Lv-shICT1(S1) and Lv-shICT1(S2) groups (p<0.001) than in Lv-shCon and Con groups. Representative images of colony size and colony numbers are shown in Figure 3C and D. Smaller and fewer colonies could be seen in Lv-shICT1(S1) and Lv-shICT1(S2) groups. However, no difference was observed in Lv-shCon and Con groups. These results indicated that knockdown of ICT1 suppressed proliferation of DU145 prostate cancer cells.

Lv-shICT1(S1)/(S2) impaired the proliferation of DU145 cells.
Knockdown of ICT1 blocked cell cycle progression in DU145 cells
It was known that cell proliferation depends on cell cycle progress. Thus, the authors analyzed cell cycle by FACS to explore the underlying mechanisms. As shown in Figure 4A and B, the cell cycle distribution in Lv-shCon and Con groups differed from that in the Lv-shICT1(S1) group, in which more cells progressed into the G2/M phase, indicating the inhibition of cell division. Thus, ICT1 could regulate cell proliferation through cell cycle control. Moreover, knockdown of ICT1 resulted in a remarkable increase in the sub-G1 phase in DU145 cells (Fig. 4C), suggesting that knockdown of ICT1 might induce cell apoptosis. In addition, Lv-shICT1(S2) was also confirmed to block cell cycle progression of DU145 cells (Supplementary Fig. S2A and B).
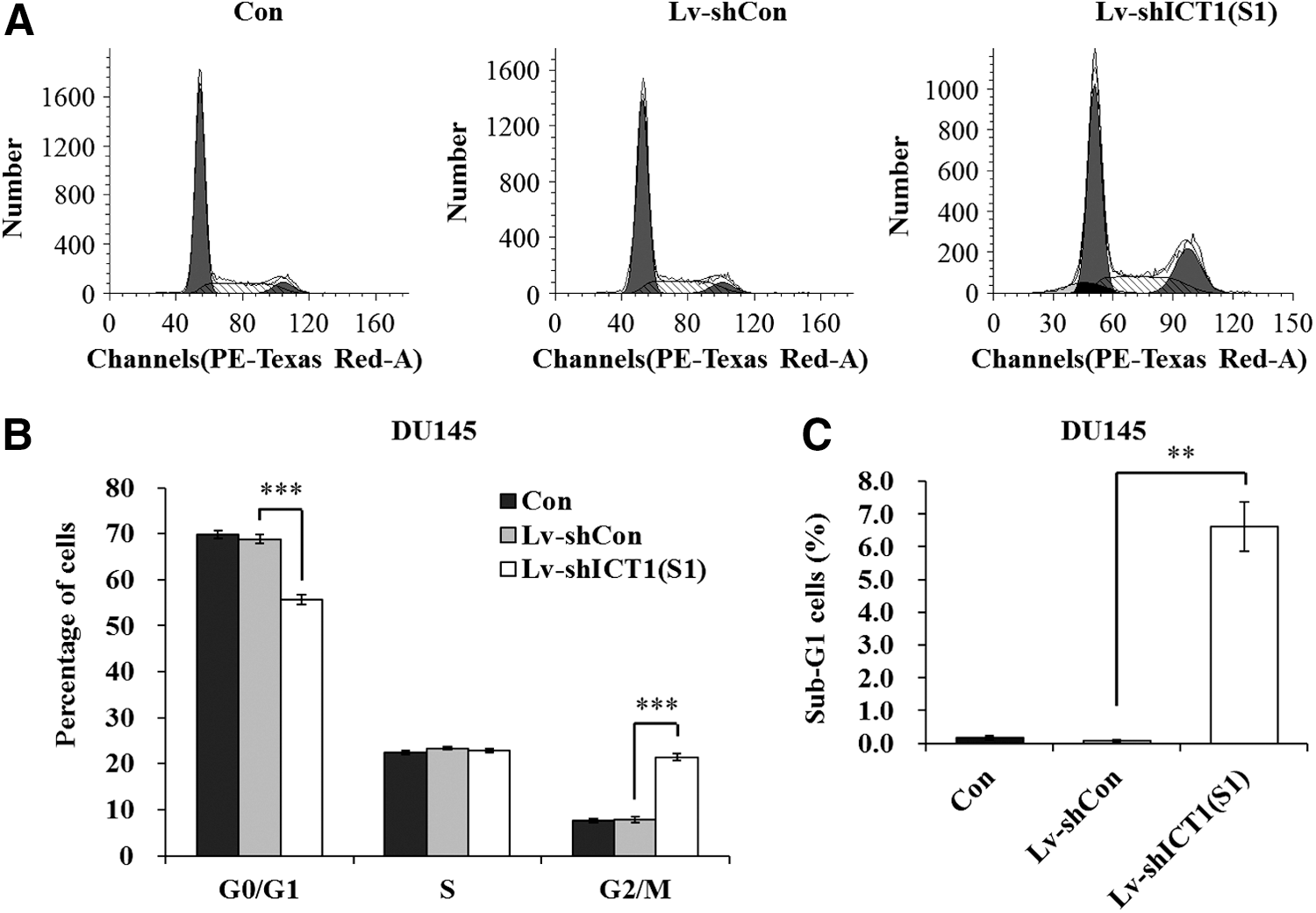
Lv-shICT1(S1) blocked the cell cycle progression in DU145 cells.
Knockdown of ICT1 induced apoptosis in DU145 cells
To verify the effect of ICT1 on cell apoptosis, the authors applied Annexin V-APC/7-AAD double staining on DU145 cells following Lv-shICT1(S1) infection (Fig. 5A). Annexin V-APC versus 7-AAD plots from the gated cells showed the populations corresponding to viable (Annexin V−/AAD−), necrotic (Annexin V−/7-AAD+), early apoptotic (Annexin V+/7-AAD−), and late apoptotic (Annexin V+/7-AAD+) cells. Knockdown of ICT1 increased late apoptotic cells by 2.4-fold, compared to the Lv-shCon group (Fig. 5B). These results indicated that knockdown of ICT1 could suppress the growth of prostate cancer cells through cell cycle arrest and apoptosis.
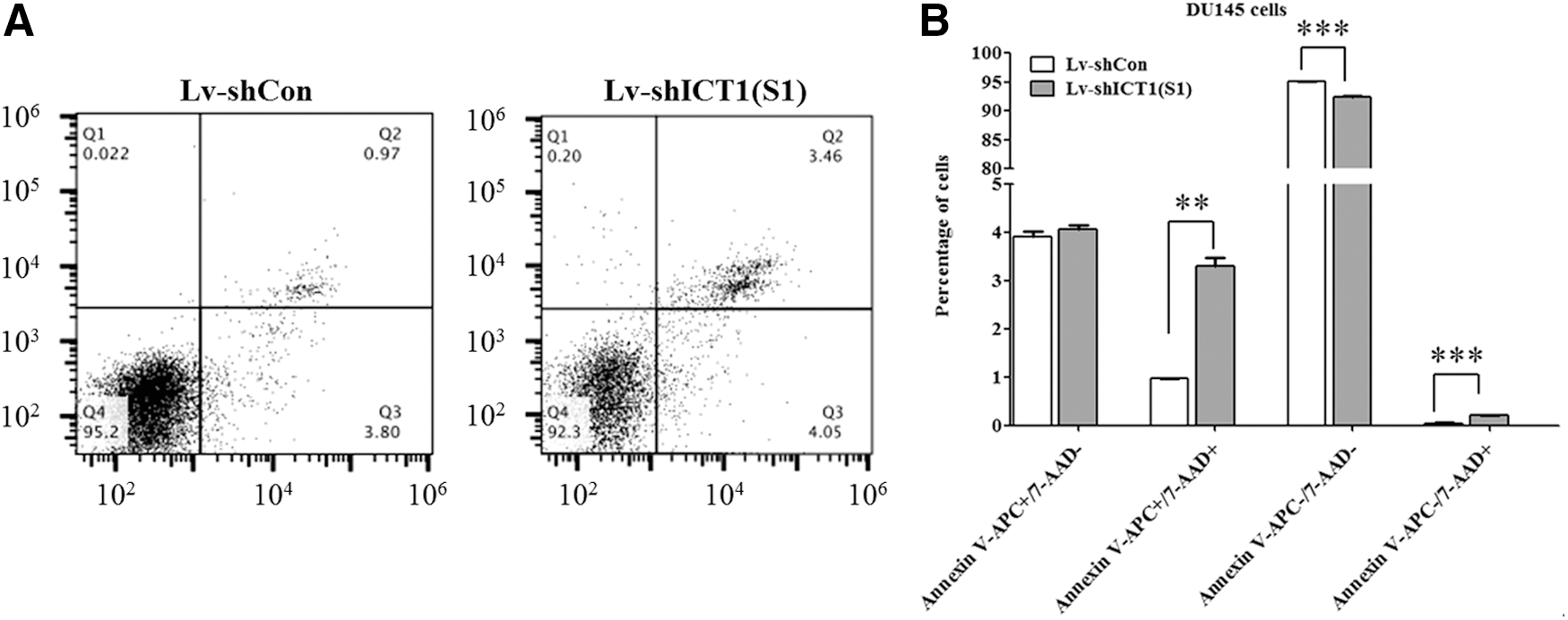
Lv-shICT1(S1) induced cell apoptosis in DU145 cells.
Knockdown of ICT1 regulated cell cycle and apoptotic markers
Furthermore, the authors detected the expression alterations of some cell cycle regulators and apoptotic markers. As shown in Figure 6, the expression levels of CDK1 and Cyclin B1, associated with G2-M transition, were decreased in the Lv-shICT1(S1) group. Besides, the expression levels of cleaved-PARP, caspase 3, and Bax were increased, while the antiapoptotic protein Bcl-2 was downregulated in DU145 cells following Lv-shICT1(S1) infection. Hence, knockdown of ICT1 induced a strong proapoptotic effect in prostate cancer cells.
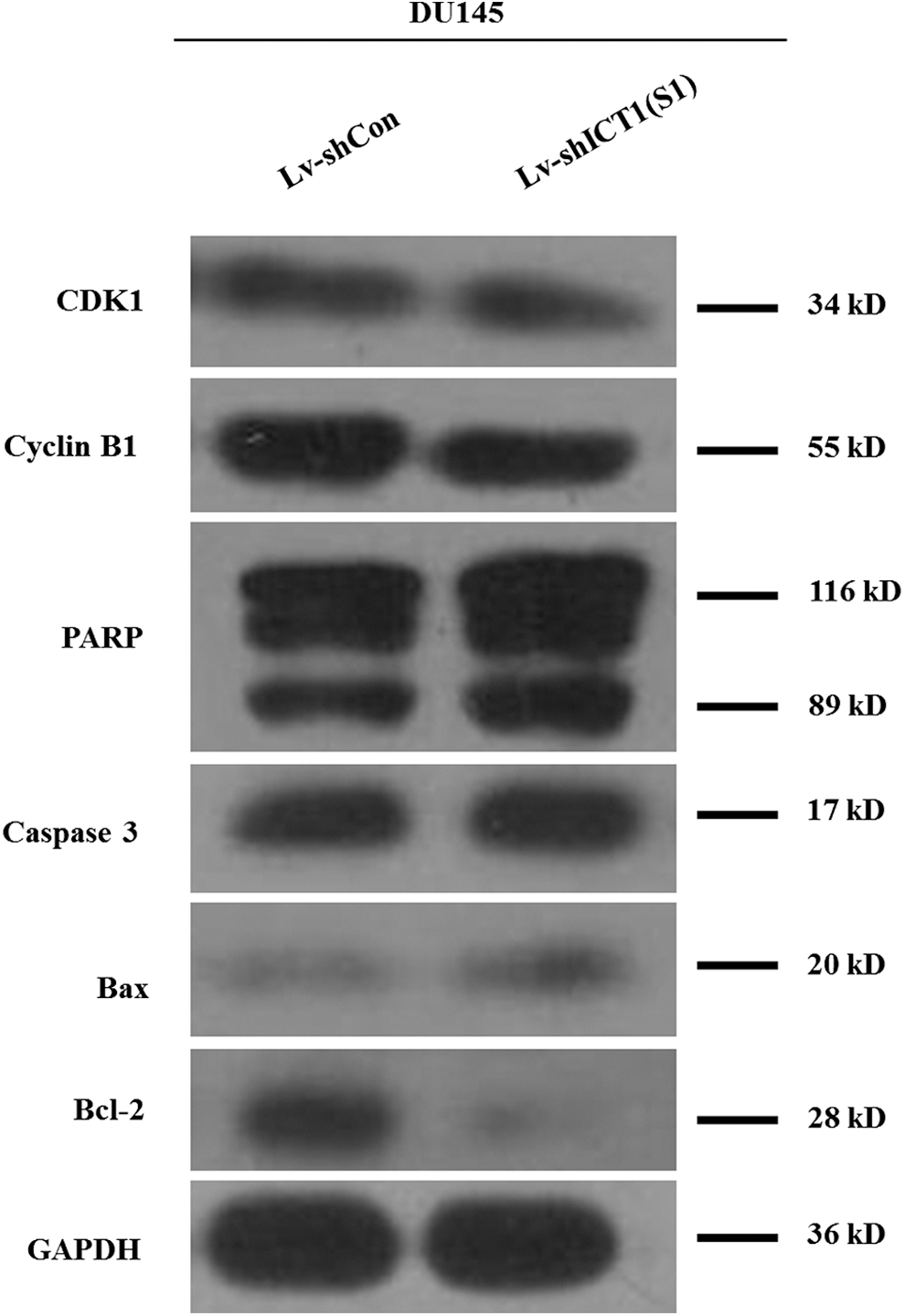
Knockdown of ICT1 regulated cell cycle regulators and apoptotic markers. Western blot analysis of CDK1, Cyclin B1, PARP, caspase 3, Bax, and Bcl-2 expression in DU145 cells following Lv-shICT1(S1) infection.
Discussion
Prostate cancer, the second leading cause of cancer death among men in the United States, affects one in seven men at some point in their lifetime. Nearly 1 in 36 men die from prostate cancer in the United States annually. More recently, ICT1 was found to be overexpressed in various kinds of cancer cells and involved in tumor progression. 14,15,17 In this study, the authors provided new evidence that ICT1 plays an essential role in cell growth and survival in prostate cancer. They selected two different ICT1 shRNAs to specifically block its endogenous expression in human androgen-independent prostate cancer cell lines DU145 and PC-3. Decreased ICT1 expression by either ICT1 shRNA-expressing lentivirus significantly inhibited cell viability and proliferation. The more efficient shRNA-targeting ICT1 was used for mechanism study.
Uncontrolled proliferation is one of the hallmarks of the cancer cell 18 that most often results from genetic alterations and/or the inactivation of master regulators of the cell cycle. 19 Compared to controls, ICT1-silenced cells were more inclined to redistribute in the G2/M phase, leading to cell cycle arrest. CDK1 and Cyclin B1 are key regulators for G2-M transition during the cell cycle. Cyclin B1 is the M-phase promoting factor and is essential for the initiation of mitosis. 20 The authors found that the expression levels of CDK1 and Cyclin B1 were decreased in ICT1-silenced cells. It is reasonable to speculate that the mechanisms of ICT1 knockdown restricting G2-M transition may be partly through suppression of CDK1 and Cyclin B1.
Flow cytometry also showed a noticeable accumulation of cells in the sub-G1 phase after ICT1 knockdown. Furthermore, Annexin V-APC/7-AAD double staining confirmed that knockdown of ICT1 increased late apoptotic cells. PARP is an abundant DNA-binding enzyme that detects and signals DNA strand breaks. 21 The presence of cleaved PARP is one of the most used diagnostic tools for the detection of apoptosis in many cell types. 22 When DU145 cells were infected with ICT1 shRNA-expressing lentivirus, a significant proteolytic cleavage of PARP was detected using western blot, indicating a proapoptotic role for ICT1 knockdown. Apoptosis is a type of programed cell death that is caspase dependent. 23 In this study, the expression of caspase 3 was obviously increased in ICT1-silenced cells. In addition, knockdown of ICT1 increased the expression of proapoptotic mitochondria protein Bax and conversely decreased the antiapoptotic mitochondria protein Bcl-2 in DU145 cells. Therefore, the authors could conclude that the growth inhibition by ICT1 silencing in prostate cancer was probably due to the induction of mitochondrial-related apoptosis. These results are supported by the previous study, which demonstrated that the lack of ICT1 in HeLa cells inhibited cell proliferation, which is due to cell cycle arrest and apoptotic cell death. 14
Human mitochondria are ubiquitous organelles essential for cell viability. Correct maintenance of mitochondrial transcription and translation system is essential for cell viability. Mitochondrial dysfunction by limiting the availability of glucose 24 or by genetic alterations 25 results in cell proliferation inhibition and apoptosis. ICT1 is identified as one of four members of the prokaryotic/mitochondrial release factor family. Moreover, depletion of ICT1 by RNAi resulted in a reduction of mitochondrial protein synthesis, leading to a loss of cell viability as well as mitochondrial dysfunction. 13,14 It will be interesting to investigate whether ICT1 knockdown affects mitochondrial activities in prostate cancer cells.
In conclusion, this study highlights the crucial role of ICT1 in promoting prostate cancer cell proliferation in vitro. The depletion of ICT1 by lentivirus-mediated shRNA or small molecular inhibitor may provide a novel therapeutic approach for the treatment of prostate cancer. Further investigations will help us illuminate the potential value of ICT1-targeted therapy in vivo.
Footnotes
Disclosure Statement
The authors declare no conflicts of interest.