
Abstract
Collagen triple helix repeat containing-1 (CTHRC1), a secreted protein, is transiently expressed in the arterial wall in response to injury, indicating that it may contribute to vascular remodeling by limiting collagen matrix deposition and promoting cell migration. Recent studies showed that it is aberrantly upregulated in most human solid tumors, yet its role in osteosarcoma remains unclear. In this study, the authors investigated the role of CTHRC1 in human osteosarcoma tumorigenesis. The authors used lentivirus-mediated short hairpin RNA (shRNA) against CTHRC1 to limit its endogenous expression in U2OS and SW1353 cells. Interestingly, they found that depletion of CTHRC1 significantly inhibited cell proliferation and colony formation in U2OS and SW1353 cells. Flow cytometry assay showed that knockdown of CTHRC1 increased the cell percentage of G0/G1 phase, resulting in cell cycle arrest in U2OS cells. Moreover, CTHRC1 silencing induced the cell cycle arrest by a decrease in the cell percentage in G0/G1 phase and increased in G2/M phase in SW1353 cells. In addition, crystal violet staining suggested CTHRC1 silencing inhibited migration of U2OS and SW1353 cells. These results demonstrated that CTHRC1 might play an important role in osteosarcoma progression.
Introduction
Osteosarcoma is a type of cancer that produces immature bone, which is usually found at the end of long bones in children and adults. Osteosarcoma is the fifth most common malignancy and originates from mesenchymal stem cells, which also has a common origin with Ewing's sarcoma and chondrosarcoma. 1 It is characterized by malignant osteoid production and osteoblastic differentiation, 2 and its rapid growth is due to the formation of tumor osteoid tissue and bone tissue through direct or indirect channels. Currently, the therapies of osteosarcoma are mainly concentrated in multiagent chemotherapy, surgery, and radiation, which are not fully adequate for osteosarcoma treatment, 3 because many patients with initially localized disease subsequently relapse again, and have an extremely poor clinical outcome. 4
Recent studies suggest that aberrant activation of wnt signaling is associated with osteosarcoma. β-catenin, which is an important mediator of wnt signaling, is elevated in the majority of osteosarcoma tumors and may be related to the migration of osteosarcoma. 5 In addition, ectopic expression of the Wnt agonist DKK3 can restrain osteosarcoma cell SAOS2 invasion and motility. 6 Simultaneously, miR-145 is able to inhibit osteosarcoma cell proliferation and invasion by targeting ROCK1. 7 The PI3K/mTOR inhibitor also inhibits osteosarcoma cell proliferation and tumor development in vivo. 8 Other researchers indicate that numerous bone morphogenetic proteins (BMPs) and bone morphogenetic protein receptors (BMPRs) are highly expressed in osteosarcoma. 9,10 Overexpression of BMPR-II is likely to be the regulator of recurrence of malignant osteosarcoma. 11 BMI, a member of the polycomb group family of transcriptional regulators, is found overexpressed in different types of osteosarcomas; BMI-1 knockdown significantly impairs cell migration and colony formation in osteosarcoma cells. 12 However, molecular mechanisms and natural history of osteosarcoma remain unclear. Therefore, it is impending to find more effective target molecules of osteosarcoma.
Collagen triple helix repeat containing-1 (CTHRC1), a 25-kDa secreted glycoprotein, is initially found in a screen for differentially expressed genes in balloon injured versus normal rat arteries. 13 The expression of CTHRC1 in injured arteries could promote cell migration and inhibited collagen I synthesis, suggesting that CTHRC1 may be involved in the tissue repair process during vascular remodeling. 13,14 Studies on the role of CTHRC1 in human solid cancers indicate that CTHRC1 expression is highly upregulated in cancers and seems to be associated with cancer cell invasion and metastasis. 15,16 According to expression analysis of CTHRC1, abundant expression of CTHRC1 is observed in the developing skeleton in mouse embryos. 17 It has been found that CTHRC1 expresses in bone tissues in vivo and has been identified as a downstream target of bone morphogenetic protein-2 in osteochondroprogenitor-like cells, and thus, it might act as a positive regulator of osteoblastic bone formation to increase bone mass. 18 In addition, CTHRC1, which is also secreted by mature bone resorbing osteoclasts, can target stromal cells to stimulate osteogenesis, indicating that CTHRC1 is a coupling factor that regulates bone remodeling. 19 Taking into account that CTHRC1 can regulate bone formation and its critical role in many human solid tumors, the authors speculate whether CTHRC1 is involved in the progress of osteosarcoma or not.
In this present study, they constructed lentivirus-mediated short hairpin RNA (shRNA) to knock down the expression of CTHRC1 in U2OS and SW1353 cells, aiming to discover its function in osteosarcoma progression. This work suggested that depletion of CTHRC1 not only inhibited cell proliferation but also blocked the migration of U2OS and SW1353 cells. The authors also found that CTHRC1 silencing could affect the cell cycle distribution and caused cell cycle arrest at G0/G1 phase in U2OS cells and at G2/M phase in SW1353 cells. To sum up, these results indicate that CTHRC1 is a positive regulator of osteosarcoma pathogenesis.
Materials and Methods
Materials
Cell culture media for U2OS, SW1353, and 293T cells were obtained from HyClone and fetal bovine serum (FBS) was from Biowest. Lentivirus packing vectors were from Shanghai Hollybio. Lipofectamine 2000 was purchased from Invitrogen. For motility assay, the authors utilized the transwell apparatus, which was from Costar.
Cell culture
Human osteosarcoma cell lines U2OS, SW1353 and human embryonic kidney cell line 293T were purchased from the Cell Bank of Chinese Academy of Sciences (Shanghai, China). Both cells were cultured in Dulbecco's modified Eagle's medium (DMEM) (HyClone) with 10% FBS (Biowest) in 5% CO2 at 37°C. The 293T cell was mainly used to construct lentivirus packing shCTHRC1.
Construction of recombinant lentivirus
The target gene CTHRC1 (NM_138455) was searched in the website of National Center for Biotechnology Information (NCBI) to analyze the target small interfering RNA (siRNA) sequence, in which the BLAST search was used to make sure the availability of the sequence. The sequences of CTHRC1 targeted by shRNA are as follows: 5′-GCGTTGGTATTTCACATTCAACTCGAGTTGAATGTGAAATACCAACGCTTTTT-3′ (s1) and 5′-CTTGGAATGGTTCACTTAAATCTCGAGATTTAAGTGAACCATTCCAAGTTTTT-3′ (s2). Both s1 and s2 sequences were inserted into the pFH-L vector (Shanghai Hollybio), which was tagged with the green fluorescent protein (GFP). The nonsilencing control sequence (Shanghai Hollybio) was 5′-GCGGAGGGTTTGAAAGAATATCTCGAGATATTCTTTCAAACCCTCCGCTTTTTT-3′. Lentiviruses were produced by triple transfection of 80% confluent HEK293T cells with modified PFH-L plasmid and pHelper pVSVG-I and pHelper pCMVΔR8.92 plasmids (Shanghai Hollybio) using Lipofectamine 2000 (Invitrogen). Recombinant lentiviruses were harvested in a serum-free medium after 4 days, filtered, and concentrated in primed Centricon Plus-20 filter devices (Millipore). For lentivirus transfection, U2OS cells (5 × 104 cells per well) and SW1353 cells (5 × 104 cells per well) were seeded in six-well plates and infected with CTHRC1-shRNA (shCTHRC1(1)/(2)) or control shRNA (shCon) expressing lentivirus at a multiplicity of infection of 20 in U2OS cells and of 80 in SW1353 cells for 6 hours in the serum-free medium; then, the medium was replaced with the normal DMEM with 10% fetal bovine serum and cultured for 96 hours in 5% CO2 at 37°C.
RNA extraction and real-time PCR
Total RNAs from U2OS and SW1353 cells after 5 days of infection were prepared using the total RNA isolation solution (Gibco) according to the manufacturer's protocol. The cDNA synthesis reaction was done using 4 μg total RNA for each sample, using the cDNA SuperScript First-Strand Synthesis System (Invitrogen). A portion of the resultant cDNA was used for quantitative real-time reverse transcription-PCR on BioRad Connet Real-Time PCR platform with SYBR Green PCR core reagents. The primer sequences for the genes tested include the following: CTHRC1, 5′-GACCAAGGAAGCCCTGAAAT-3′ (forward) and 5′-CAACCCAGATAGCAACATCC-3′ (reverse); β-actin, 5′-GTGGACATCCGCAAAGAC-3′ (forward) and 5′-AAAGGGTGTAACGCAACTA-3′ (reverse). The amplification conditions were initial denaturation at 95°C for 1 minute followed by 40 cycles of denaturation at 95°C for 5 seconds and annealing extension of 60°C for 20 seconds. The cycle number of threshold (CT) was recorded for each reaction. The CT value of CTHRC1 was normalized to that of β-actin of the same sample.
Western blot analysis
Whole cell extracts were prepared with an ice-cold cell lysis buffer (2% mercaptoethanol, 20% glycerol, and 4% SDS in 100 mM Tris-HCl buffer, pH 6.8), and the protein concentration was determined by the enhanced BCA Protein Assay Kit (Beyotime). Equal amounts (30 μg) of protein in each lane were separated by 10% SDS-PAGE and transferred onto polyvinylidene difluoride membranes (Millipore). Membranes were blocked and then incubated with primary antibodies, rabbit anti-CTHRC1 (1:1000; ProteinTech, #16534-1-AP) and rabbit anti-GAPDH (1:10,000; ProteinTech, #10494-1-AP), overnight at 4°C. After washing with Tris buffered saline with Tween, the blots were incubated with the horse radish peroxidase-labeled anti-rabbit (1:5000; Santa Cruz, #Sc-2054) secondary antibody for 2 hours at room temperature and then visualized by the super enhanced chemiluminescence detection reagent (Applygen).
MTT assay
The authors utilized MTT plus acidic isopropanol to detect cell survival. After 96 hours of lentivirus infection, U2OS and SW1353 cells from different groups were seeded at an initial density of 2000 cells/well and 2500 cells/well separately in 96-well plates for 24 hours. Then, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) was added into each well at a final concentration of 5 mg/mL for 4 hours. Acidic isopropanol (10% SDS, 5% isopropanol, and 0.01 mol/L HCl) was then added to end the reaction and measured with an ELISA reader (Bio-Rad) at a wavelength of 595 nm. Percentage inhibition was calculated using the following formula:
Percentage growth inhibition = [(means absorbance of the shCon cells) − (means absorbance of the shCTHRC1(1)/(2) cells)] ×100/(means absorbance of the shCon cells).
Colony formation analysis
Colony formation assay was performed to detect monolayer colony formation. After 96 hours of lentivirus infection, U2OS and SW1353 cells from different groups were seeded into six-well plates at a density of 600 and 500 cells/well, respectively. All groups were fixed with methanol for 15 minutes and stained with crystals purple for 30 minutes after inoculation for 9 days. The number of visible colonies that contained more than 50 cells in each colony was counted under a microscope. Image analysis was conducted using Image-Pro® Plus Version 6.0 (Media Cybernetics).
Flow cytometry assay
To analyze the impact of knockdown CTHRC1 on cell cycle, flow cytometry was performed with propidium iodide (PI) staining. After lentivirus infection for 8 days, U2OS and SW1353 cells from different groups were seeded in a 6-cm dish at an inoculation density of 6 × 104 infected cells/well and 1 × 105 infected cells/well, respectively. The cells were cultured for 40 hours in an incubator and collected when the cell concentration reached 80%. Then, the cells were fixed with ice-cold 70% ethanol for 30 minutes at 4°C after washing with ice-cold PBS. The suspension was filtered through 400 meshes and stained with PI in the dark for 30 minutes at 4°C. The tests of DNA content were carried out independently thrice for each sample, and analysis was performed by fluorescence-activated cell sorter (Navios; Beckman Coulter). The cell proportions in the different cell cycle phase were analyzed by the software of WinMDI.
Cell migration assay
After 96 hours of lentivirus infection, U2OS and SW1353 cells from different groups were seeded in 24-well plates (4 × 104 cells/well) with a fibronectin-coated polycarbonate membrane insert (6.5 mm in diameter with 8.0 mm pores) in a transwell apparatus and cultured in DMEM. The cells were cultured in the upper chamber with the serum-free medium, and the lower chamber was filled with DMEM containing 10% FBS. After incubation for 24 hours, the cells that migrated to the bottom surface of the insert were stained by crystal violet and washed by 10% methyl alcohol. The absorbance was measured with an instrument at wavelength of 570 nm.
Statistical analysis
Statistical analysis was carried on by using SPSS19.0 (IBM). Comparisons between different groups were determined by the Student's t-test and data are expressed as mean ± SD. All experiments were performed at least three times. p Values <0.05 were considered statistically significant.
Results
Lentivirus-mediated knockdown of CTHRC1 inhibited the expression of CTHRC1
To test the speculation about the relationship of osteosarcoma and CTHRC1, which plays an important role in bone modeling or remodeling, 18 the authors used lentivirus-mediated shRNAs targeting CTHRC1 (shCTHRC1(1)/(2)) to suppress its expression in U2OS and SW1353 cells. Recombinant lentivirus infection efficiency reached more than 80% as evidenced by GFP expression 4 days after infection in both shCon and shCTHRC1 groups (Fig. 1A, E). Real-time PCR analysis showed that the mRNA levels of CTHRC1 were markedly reduced in shCTHRC1(1)- and shCTHRC1(2)-infected groups in U2OS and SW1353 cells, respectively (Fig. 1B, C, and F). Western blot analysis confirmed the silencing of CTHRC1, as only weak bands were detected in both the shCTHRC1-infected groups in U2OS cells (Fig. 1D). These data indicated that recombinant lentivirus-mediated shRNA could clearly suppress CTHRC1 expression in U2OS and SW1353 cells.

Targeted depletion of CTHRC1 by lentivirus-mediated shRNA in U2OS and SW1353 cells.
Depletion of CTHRC1 restrained proliferation of U2OS and SW1353 cells
To further explore the role of CTHRC1 in osteosarcoma proliferation, the authors performed an MTT assay to detect the effect of CTHRC1 silencing on the proliferation of U2OS and SW1353 cells. As shown in Figure 2A, the growth curve of shCTHRC1(1)-infected U2OS cells was significantly lower compared to the shCon-infected group on day 4 (2.275 ± 0.083 vs. 3.213 ± 0.125, p < 0.001) and day 5 (2.858 ± 0.020 vs. 4.787 ± 0.162, p < 0.001) after incubation. However, there was no obvious difference between the control (Con) and shCon groups. Similarly, the growth curve of shCTHRC1(2)-infected U2OS cells was also significantly dropped with an inhibitory rate of 31% on day 5, compared to the shCon-infected group (day 4: 4.752 ± 0.041 vs. 7.142 ± 0.009, p < 0.001; day 5: 5.603 ± 0.048 vs. 8.537 ± 0.079, p < 0.001) (Fig. 2B). Similar to the above results, the growth rate was significantly lowered in the shCTHRC1(1) group (day 4: 1.058 ± 0.040 vs. 2.115 ± 0.084, p < 0.001; day 5: 1.244 ± 0.051 vs. 3.102 ± 0.106, p < 0.001) and shCTHRC1(2) group (day 4: 1.210 ± 0.029 vs. 2.115 ± 0.084, p < 0.001; day 5: 1.536 ± 0.045 vs. 3.102 ± 0.106, p < 0.001) compared with shCon in SW1353 cells (Fig. 2C). These results indicated that knockdown of CTHRC1 could potently inhibit the proliferation of U2OS and SW1353 cells.
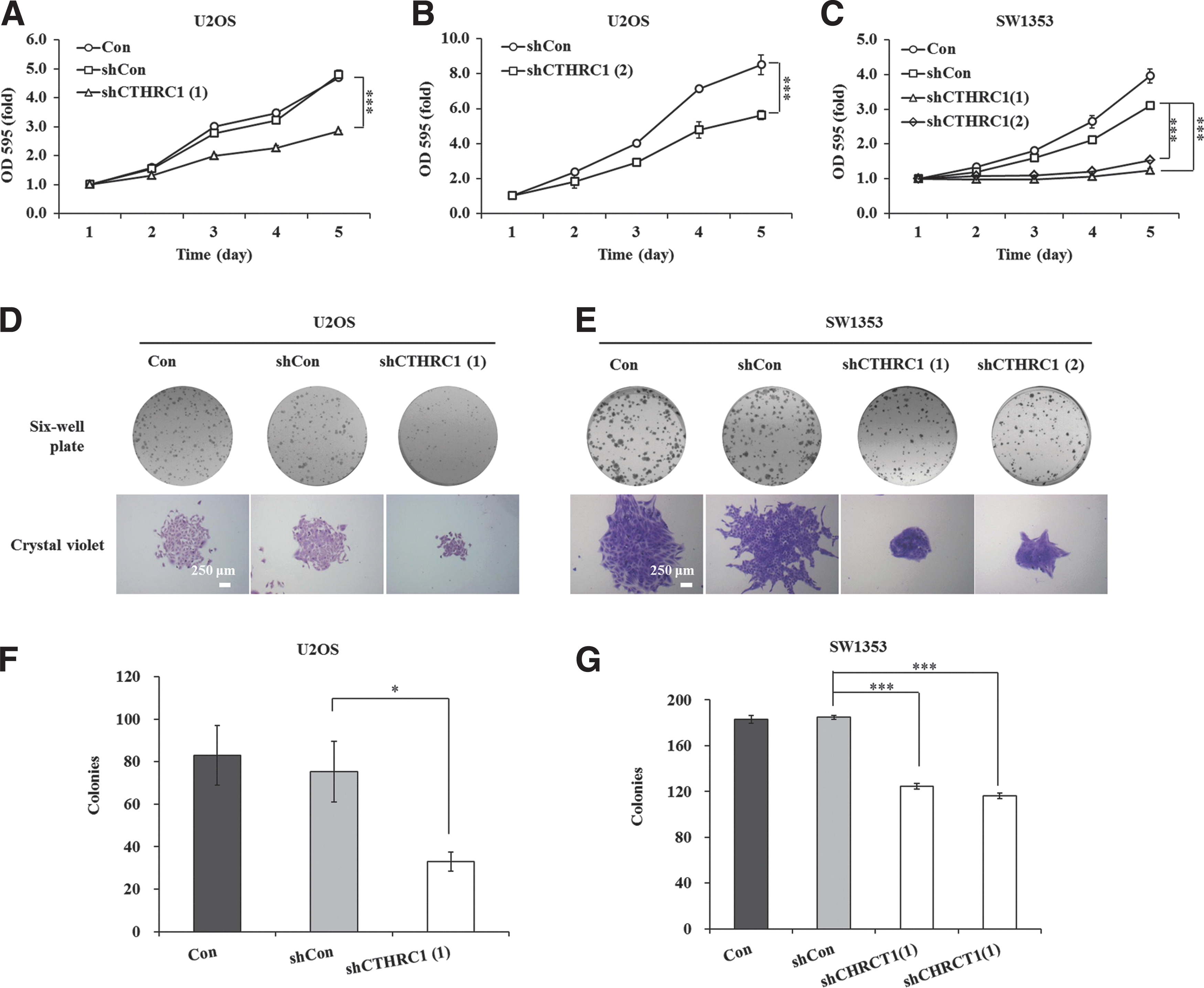
Depletion of CTHRC1 restrained cell proliferation in U2OS and SW1353 cells.
Simultaneously, colony formation assay in monolayer culture indicated that downregulation of CTHRC1 expression dramatically inhibited colony formation compared with the control groups in U2OS (Fig. 2D) and SW1353 cells (Fig. 2E). This phenotype that the number of colonies was reduced was also confirmed by colony counting in the shCTHRC1(1)-infected group compared with the shCon group (33 ± 5 vs. 75 ± 14, p < 0.05, Fig. 2F) in U2OS cells. Of course, the number of colonies were also apparently lower in the shCTHRC1(1) group (125 ± 3 vs. 185 ± 2, p < 0.001) and shCTHRC1(2) group (116 ± 3 vs. 185 ± 2, p < 0.001) than in the shCon group in SW1353 cells (Fig. 2G). However, there was no obvious difference between the control (Con) and shCon groups in U2OS and SW1353 cells. Therefore, CTHRC1 expression is detrimental to cell growth and colony formation of osteosarcoma cells and it may play a crucial role in osteosarcoma tumorigenesis.
CTHRC1 knockdown caused cell cycle arrest of U2OS and SW1353 cells
Many studies showed that cancer cells are characterized by rapid growth and division and most drug targets act on cancer cell cycle arrest. To clarify the underlying mechanisms in CTHRC1 promoting osteosarcoma cell proliferation, the authors speculated whether inhibition of CTHRC1 expression altered cell cycle in U2OS and SW1353 cells. Therefore, cell cycle progression was examined by flow cytometry in U2OS and SW1353 cells after CTHRC1 silencing (Fig. 3A, D). Intriguing, they observed an obvious increase in the number of cells in the G0/G1 phase (46.08% ± 0.02% vs. 42.01% ± 0.46%, p < 0.01), but a decrease in the number of S phase cells (35.34% ± 0.60% vs. 38.44% ± 0.40%, p < 0.01) in shCTHRC1(1)-infected group compared with the shCon group in U2OS cells (Fig. 3B). In addition, the cell percentage of sub-G1 phase, which represents cells undergoing apoptosis, also was augmented in the shCTHRC1(1)-infected group (0.54% ± 0.05% vs. 0.41% ± 0.05%, p < 0.05) (Fig. 3C). Moreover, compared to the shCon group, the cell proportion in G0/G1 phase was decreased and in G2/M phase was increased in both shCTHRC1(1) group (G0/G1 phase: 12.20% ± 0.28% vs. 23.26% ± 0.48%, p < 0.001; G2/M phase: 83.01% ± 0.96% vs. 67.18% ± 0.68%, p < 0.001) and shCTHRC1(2) group (G0/G1 phase: 17.69% ± 0.43% vs. 23.26% ± 0.48%, p < 0.001; G2/M phase: 73.78% ± 0.32% vs. 67.18% ± 0.68%, p < 0.001) in SW1353 cells (Fig. 3E). Similar to the results in U2OS cells, the cell percentage of sub-G1 phase was observed as a significant increase of cell percentage in the shCTHRC1(1)-infected group (2.47% ± 0.27% vs. 0.37% ± 0.08%, p < 0.01) and shCTHRC1(2)-infected group (2.18% ± 0.10% vs. 0.37% ± 0.08%, p < 0.01) compared with the shCon group in SW1353 cells (Fig. 3F). Hence, this study showed that downregulation of CTHRC1 could restrain cell proliferation by altering cell cycle progression and inducing apoptosis in U2OS and SW1353 cells.
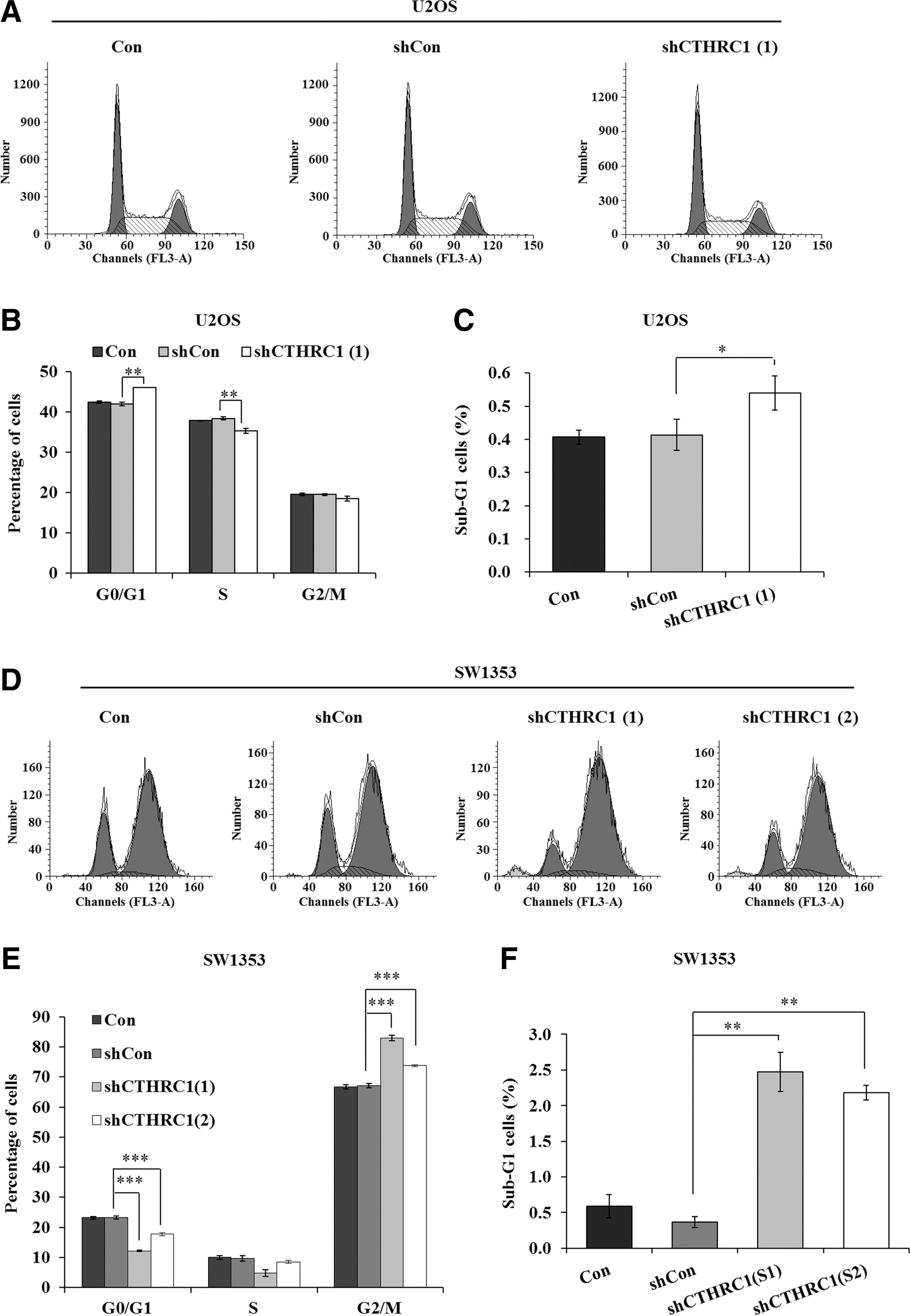
Knockdown of CTHRC1 caused cell cycle arrest and apoptosis in U2OS and SW1353 cells.
Knockdown of CTHRC1 inhibited migration of U2OS and SW1353 cells
Next, the authors further explored the effect of CTHRC1 on cell migration in U2OS and SW1353 cells using transwell migration assay (Fig. 4A, C). As shown in Figure 4B and D, the migration abilities of U2OS and SW1353 cells were significantly reduced in the shCTHRC1(1)-infected group (U2OS cells: 0.10 ± 0.002 vs. 0.14 ± 0.02, p < 0.05; SW1353 cells: 0.85 ± 0.01 vs. 1.52 ± 0.11, p < 0.01) and the shCTHRC1(2)-infected group (U2OS cells: 0.10 ± 0.02 vs. 0.14 ± 0.02, p < 0.05; SW1353 cells: 1.07 ± 0.07 vs. 1.52 ± 0.11, p < 0.01) compared with the shCon group. All in all, the above results showed that the cell migration ability was inhibited in the shCTHRC1(1)/(2)-infected group, which could indirectly indicate that the influence of CTHRC1 gene is not only on cell proliferation but also on cell migration in osteosarcoma.
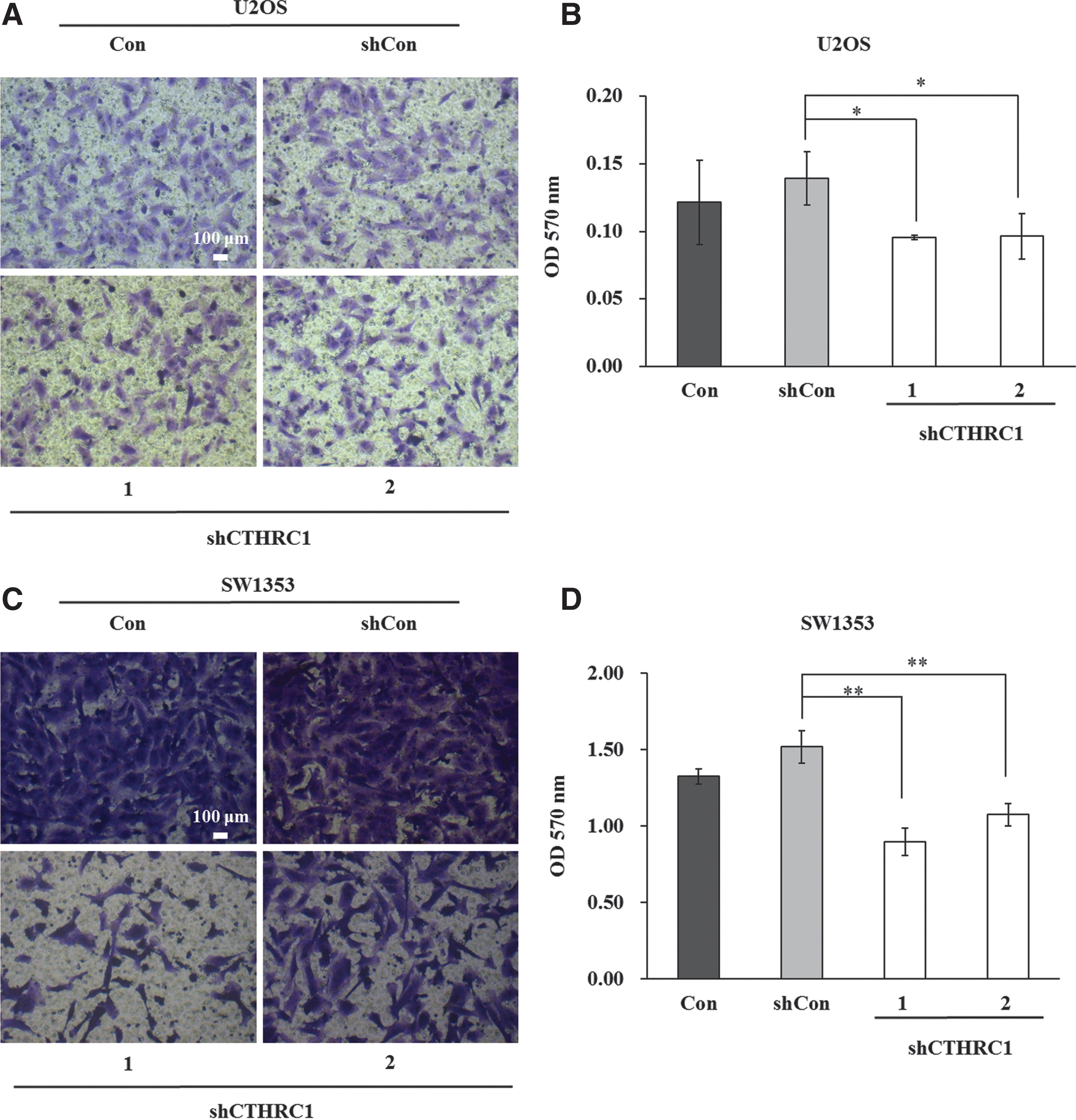
Downregulation of CTHRC1 inhibited cell migration in U2OS and SW1353 cells.
Discussion
Osteosarcoma is the most frequent primary bone sarcoma, comprising ∼20% of all bone tumors and about 5% of pediatric tumors overall. 20 Physical, chemical, and biological agents have been suggested as carcinogens for osteosarcoma. 21 In recent years, the use of surgery combined with neoadjuvant chemotherapy therapy has increased the 5-year survival rate of patients with osteosarcoma to near 50%–80%. 22,23 However, due to the rare occurrence and the heterogeneity of human osteosarcoma, the molecular research of this disease has made slow progress compared to some other cancers.
Several reports have pointed that high expression of CTHRC1 was present in many cancers, including hepatocellular carcinoma, melanoma, breast cancer, and colorectal cancer. 24,25 CTHRC1 has been shown to play important roles in breast cancer proliferation and progression. 26 In the present study, the authors first identified the relationship between CTHRC1 and osteosarcoma. The shRNAs targeting the CTHRC1 gene were transduced into the osteosarcoma cell lines U2OS and SW1353 by using lentivirus-mediated RNAi, which is an attractive tool for human cancer therapy. 27,28 Depletion of CTHRC1 successfully restrained proliferation of U2OS and SW1353 cells as measured by MTT and colony formation assays, suggesting that CTHRC1 may be a contributing factor in the growth of osteosarcoma in vitro. Moreover, flow cytometry assay was performed to detect the role of CTHRC1 in cell cycle of U2OS and SW1353 cells. The results indicated that downregulation of CTHRC1 resulted in the cell cycle arrest at G0/G1 phase in U2OS cells and at G2/M phase in SW1353 cells. The difference of cell cycle arrest in U2OS and SW1353 cells may be due to the different cell sources that U2OS cells are from a moderately differentiated sarcoma of the tibia, 29 while SW1353 cells are from a primary grade II chondrosarcoma of the right humerus obtained. 30 Meanwhile, when CTHRC1 was silenced, the number of cells in the sub-G1 phase was increased significantly in U2OS cells, indicating that knockdown of CTHRC1 might give rise to cell apoptosis in osteosarcoma. Nevertheless, the molecular mechanism of how CTHRC1 regulates osteosarcoma cell growth in osteosarcoma is poorly understood. Further study should be implemented to expound the regulatory mechanism of CTHRC1 in osteosarcoma cell growth.
Besides, CTHRC1 contributes to vascular remodeling by limiting collagen matrix deposition and promoting cell migration. 13,17 Aberrant expression of CTHRC1 is associated with cancer tissue invasion and metastasis in human solid cancers. 15 CTHRC1 has been reported to be highly expressed in invasive tumors compared with noninvasive stages of tumors, such as melanoma and breast cancer. 26 Also, suppression of CTHRC1 results in a decreased cell migration in melanoma and lung cancer cells. 15,26,31 In addition, CTHRC1 promotes pancreatic cancer progression by regulating migration and adhesion of tumor cells. 32 Similar with the results of previous reports, the authors found that depletion of CTHRC1 could also restrain migration of U2OS and SW1353 cells in this study.
In conclusion, the preliminary findings indicated that depletion of CTHRC1 could restrain proliferation and migration of U2OS and SW1353 osteosarcoma cells. Although further functional experiments are needed to clarify the role of CTHRC1 in osteosarcoma progression and to elucidate the underlying mechanisms in vitro or vivo, the results from this research already suggest that CTHRC1 silencing may be a potential therapeutic approach in osteosarcoma treatment in the future.
Footnotes
Disclosure Statement
The authors declare no conflict of interest.