
Abstract
Cancer Biotherapy and Radiopharmaceuticals (CBR)
is officially retracting the published article by Jiapeng Xu, Weimin Wang, Zhenxin Zhu, Ziran Wei, Dejun Yang, and Qingping Cai entitled, “Tumor Protein D52-Like 2 Accelerates Gastric Cancer Cell Proliferation In Vitro,” Cancer Biother Radiopharm June 2015, 30(3): 111–116; DOI: 10.1089/cbr.2014.1766
The editors of CBR received a letter from an investigator raising concerns regarding this published paper. The investigator claims that this study employed the 5′-GTAG… shRNA sequence as a “negative control”, despite the fact that according to Blastn analyses, this sequence shows homology to the human NOB1 gene. The investigator's concern is that since the article did not employ a non-targeting lentiviral construct as described in the study, it most likely invalidates the experimental results reported in the article. The authors of the article were afforded the opportunity to respond, and they disagreed with the concerns. However, the editors have decided to retract the article based on dispute and the serious nature of the plausibility of the raised concerns.
Introduction
Gastric cancer or stomach cancer, referring to cancer arising from any part of the stomach, is one of the most devastating human cancers, which causes over 700,000 deaths worldwide per year. Sixty-five percent to 80% of gastric cancers are developed from Helicobacter pylori infection. 1,2 The mechanism by which H. pylori induces gastric cancer potentially involves chronic inflammation, or the action of H. pylori virulence factors. 3 Other risk factors, including smoking, dietary factors, iodine deficiency, genetic factors, have also been involved. Surgery remains the only curative therapy for gastric cancer. Unlike other tumors, gastric cancer is difficult to cure because few symptoms emerge in early stage, the disease is usually advanced when the diagnosis is made. 4 The 5-year survival rate for gastric cancer is reported to be less than 10%. 5,6 The poor prognosis of gastric cancer generally results from the fact that the tumor has often metastasized by the time of discovery and the fact that most people with the condition are elderly (median age is between 70 and 75 years) at presentation. In recent years, antisense therapy has arisen as a promising therapy for many diseases, including cancer. Once a gene is identified as an indispensable factor special in tumor cell proliferation, this therapeutic strategy is considered to be efficient to turn this gene off, leading to the death of cancer cell. Therefore, the development of target genes is an urgent need for the treatment of gastric cancer.
The tumor protein D52 is frequently overexpressed in several carcinomas and has been identified as a B cell differentiation marker. D52-like genes are also differentially expressed in particular hematological malignancies, where transcript or protein levels may reflect cellular proliferative or differentiative status. TPD52L2, alternatively named as TPD54, DKFZp686A1765, or HCCR-binding protein 2, is a member of the tumor protein D52-like family. 7 These proteins are characterized by an N-terminal coiled-coil motif that is used to form homo- and heteromeric complexes with other tumor protein D52-like proteins, 8 as well as N- and C-terminally located PEST sequences, which have been linked with regulating protein stability. 9 Alternatively spliced transcript variants encoding multiple isoforms hD52 (TPD52, PrLZ), hD53 (TPD52L1), hD54 (TPD52L2), hD55 (TPD52L3/NYD-SP25) have been observed for this gene. 10,11 hD52, as well as its two homologues hD53 and hD54, have been identified to be overexpressed in human breast carcinoma. 8 Moreover, hD52 and hD54 transcripts were frequently detected in acute lymphoblastic leukemia (ALL) and acute myelocytic leukemia, where they were frequently coexpressed. 12 Nakamura et al. identified TPD52L2 associated with tumor recurrence of pancreatic cancers, which has shown its clinical significance as a prognostic biomarker. 13 Recently, TPD52L2 was also shown to mediate the positive regulation of hABCF3 on cell proliferation of human liver cancer cell lines in vitro through the interaction with hABCF3. 14 Previous evidence suggests TPD52L2 as an oncogenic protein, just as its name. However, the pathological function of TPD52L2 in other tumors like gastric cancer remains unclear.
Thus, in the present study, the authors utilized lentivirus-mediated knockdown of TPD52L2 to explore its role in gastric cancer cell proliferation and underlying mechanisms.
Materials and Methods
Cell culture
Human gastric cancer cell lines MKN-28, SGC-7901, MGC80-3, and AGS and human embryonic kidney cell line 293T (HEK293T) were obtained from the Cell Bank of Shanghai Institute of Cell Biology, Chinese Academy of Sciences (Shanghai, China). MKN-28 and SGC-7901 cells were maintained in RPMI1640 (Hyclone) supplemented with 10% fetal bovine serum (FBS; Hyclone). AGS cells were cultured in F-12 (Sigma) with 10% FBS. MGC80-3 and HEK293T cells were maintained in Dulbecco's modified Eagle's medium (Hyclone) supplemented with 10% FBS. All cell lines were cultured at 37°C in a humidified atmosphere of 5% CO2.
Construction of recombinant lentivirus and gene silencing
The short hairpin RNA (shRNA) sequence (5′-GCGGAGGGTTTGAAAGAATATCTCGAGATATTCTTTCAAACCCTCCGCTTTTTT-3′) was selected for human TPD52L2 gene (NM_199360) after screening to validate potential shRNA and it was inserted into the lentiviral expression vector pFH-L (Shanghai Hollybio), which contains a reporter gene GFP. Scrambled shRNA (5′-CTAGCCCGG CCAAGGAAGTGCAATTGCATACTCGAGTATGCAATTGCACTTCCTTGGTTTTTTGTTAAT-3′) was used as a negative control. Recombinant lentivirus was generated by triple transfection of 80% confluent HEK293T cells with modified pFH-L vector and packing plasmids pVSVG-I and pCMVΔR8.92 (Shanghai Hollybio) using Lipofectamine 2000 (Invitrogen). For lentivirus infection, cells were seeded into six-well plates and transfected with TPD52L2 shRNA (Lv-shTPD52L2) or control shRNA (Lv-shCon) expressing lentivirus at a multiplicity of infection (MOI) of 6.
RNA extraction and real-time quantitative PCR
After 5 days infection, total RNA was isolated from MGC80-3 cells using TRIzol reagent (Gibco) according to the manufacturer's instruction. Five micrograms total isolated RNA was used to synthesize the first strand of cDNA using the SuperScript II RT 200 U/mL (Invitrogen). TPD52L2 mRNA expression was evaluated by real-time quantitative PCR (RT-qPCR) on the Bio-Rad Connect Real-Time PCR platform with SYBR Green PCR Core Reagents (reaction system: 2× SYBR premix Ex Taq 10 μL, forward and reverse primers [2.5 μM] 0.8 μL, cDNA 5 μL, ddH2O 4.2 μL). After initial denaturation at 95°C for 1 minute, a total of 40 cycles (denaturation 95°C, 5 seconds; annealing extension of 60°C, 20 seconds) proceeded, and absorbance was obtained at the extension stage. β-actin was used as the internal reference control. The PCR primers were as following: TPD52L2-F: 5′-TTCACAGGCAGGACAGAAGA-3′, TPD52L2-R: 5′-TTGAAGGTCGCAGAGTTCCT-3′; β-actin-F: 5′-GTGGACATCCGCAAAGAC-3′, β-actin-R: 5′-AAAGGGTGTAACGCAACTA-3′. Data were analyzed using 2−ΔΔCt statistical method. Results were presented as CT values, which were defined as the threshold PCR cycle number at which an amplified product is first detected. The average CT was calculated for both TPD52L2 and β-actin, and ΔCT was determined as the ratio of the mean of the triplicate CT values for TPD52L2 to the mean of the triplicate CT values for β-actin. Tests were performed in triplicate for each sample.
Western blot analysis
Cultured cells were washed with ice-cold phosphate buffered saline (PBS) and then lysed in 2× sodium dodecyl sulfate (SDS) sample buffer (100 mM Tris-HCl [pH 6.8], 10 mM EDTA, 4% SDS, 10% glycine) for 1 hour at 4°C. The lysates were clarified by centrifugation at 13,000 g for 30 minutes at 4°C and the supernatants were employed for further analysis. The total protein concentration was estimated using the BCA protein assay kit. Protein samples (30 μg) were loaded and electrophoresed in a 10% SDS–polyacrylamide gel electrophoresis (SDS-PAGE) at 50 V for 3 hours, and subsequently transferred to polyvinylidene difluoride (PVDF) membranes (Millipore) at 300 mA for 1.5 hours. After being blocked with Tris buffered saline Tween-20 (TBST) (20 mM Tris [pH 7.6], 150 mM NaCl, 0.01% Tween-20) containing 5% (m/v) defatted dry milk for 1 hour at room temperature, membranes were probed with (goat anti-TPD52L2 [#SAB2501053, dilution 1:500; Sigma], rabbit anti-poly ADP-ribose polymerase (PARP) [#9542, dilution 1:1000; Cell Signaling Technology] or rabbit anti-GAPDH [#10494-1-AP, dilution 1:60,000; Proteintech Group, Inc.]) primary antibody overnight at 4°C. After washing by TBST for three times, the membrane was incubated with horseradish peroxidase (HRP)-labeled donkey anti-goat HRP (#A0181, dilution 1:5000; Beyotime) or goat anti-rabbit HRP (#Sc-2054, dilution 1:5000; Santa Cruz) secondary antibody at room temperature for 2 hours. The membranes were analyzed using the Super ECL Detection Reagent and film was developed in a dark room.
MTT cell viability assay
Briefly, cells from different groups (Lv-shTPD52L2, Lv-shCon, Con) were seeded in 96-well plates at a density of 2000 cells per well. At indicated time points, 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) was added into each well at a final concentration of 5 mg/mL for 4 hours. Acidic isopropanol (10% SDS, 5% isopropanol, and 0.01 M HCl) was then added to stop the reaction and measured with an ELISA reader (Bio-Rad) at a wavelength of 595 nm. Viability of cells was calculated from theoretical absorbance. Tests were performed in triplicate for each sample.
Colony formation assay
To assay monolayer colony formation, MGC80-3 cells from different groups (Lv-shTPD52L2, Lv-shCon, Con) after 96 hours of infection were plated into six-well plates at a density of 500 cells per well. After 6 days of culture at 37°C, cells were washed with PBS and fixed with 4% paraformaldehyde for 30 minutes at room temperature. The fixed cells were then stained with Crystals Purple for 10 minutes, washed with double-distilled water and air dried. The number of colonies was counted using light/fluorescence microscopy. Tests were performed in triplicate for each sample.
Cell cycle analysis
Cell cycle analysis was carried out by Propidium Iodide (PI) staining method using flow cytometry. Briefly, after 3 days of infection at an MOI of 6, MGC80-3 cells were seeded in 6-cm dishes (200,000 cells/dish) and cultured for 40 hours at 37°C. MGC80-3 cells in each dish were harvested at a cell confluence of 80%, followed by washing with ice-cold PBS and fixing with 70% cold alcohol. After centrifugation, cells were resuspended in PI/RNase/PBS (100 μg/mL PI and 10 μg/mL RNase A) solution and incubated in the dark for 30 minutes at room temperature. The suspension was filtered through a 50-mm nylon mesh, and the DNA content of stained nuclei was analyzed by PI staining. PI uptake was evaluated by Fluorescence Activated Cell Sorting on flow cytometer (FACS Calibur; BD Biosciences). Tests were performed in triplicate for each sample.
Statistical analyses
All statistical analyses were performed using the SPSS13.0 software. The differences between groups were compared using Student's t-test, and data were expressed as mean±standard deviation of three independent experiments. Statistically significant difference was accepted at p<0.05.
Results
Lentivirus-mediated RNAi inhibits TPD52L2 expression in gastric cancer cells
To explore the role of TPD52L2 in gastric cancer, the expression levels of TPD52L2 protein were firstly detected in different gastric cancer cell lines by western blot. As shown in Figure 1A, TPD52L2 was widely expressed in most gastric cancer cell lines SGC-7901, MGC80-3, and AGS, but little in MKN-28. The authors tend to knock down the TPD52L2 expression through lentivirus-mediated RNAi technique. The efficiencies of lentivirus infection were significantly high (>80%) in all these four cell lines as indicated by reporter gene GFP expression (Fig. 1B and Supplementary Fig. S1A–C; Supplementary Data are available online at
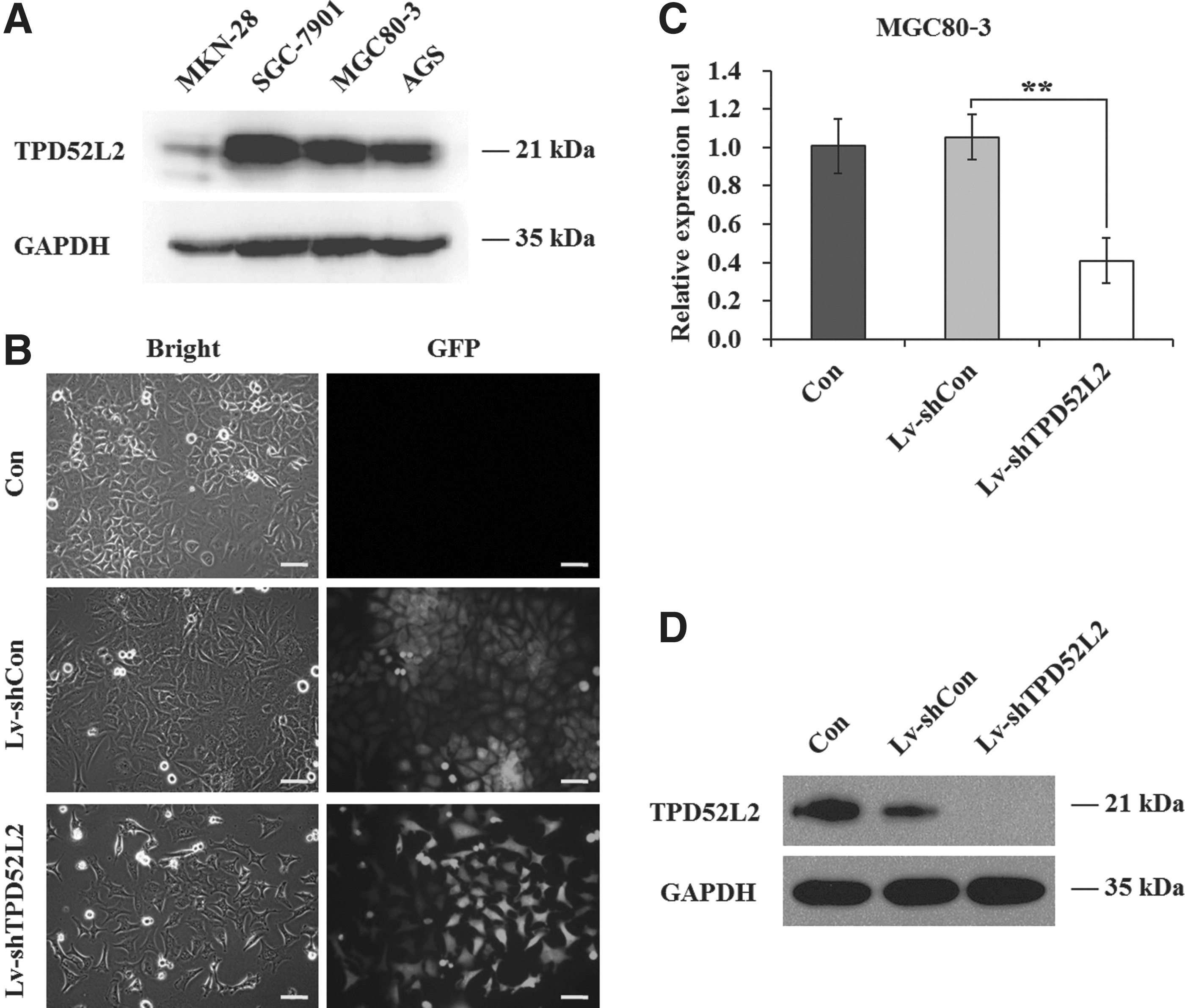
Lentivirus-mediated knockdown of tumor protein D52-like 2 (TPD52L2) in MGC80-3 cells.
Downregulation of TPD52L2 leads to cell proliferation inhibition in gastric cancer cells
TPD52L2 is extensively expressed in most of the human tissues; it was primarily described as a positive regulator in cell proliferation. To elucidate the effect of TPD52L2 on gastric cancer malignant proliferation, the authors examined the cell viability and proliferation of Lv-shTPD52L2-infected cells by MTT and colony formation assays. As shown in Figure 2A, the proliferation rate after 4 or 5 days was strongly inhibited in TPD52L2 knockdown cells, respectively (0.521±0.020 vs. 1.000±0.029, p<0.001; 0.612±0.012 vs. 1.650±0.052, p<0.001). Whereas there was no significant difference between the Lv-shCon and Con groups, suggesting the specific inhibition of cell proliferation in Lv-shTPD52L2-infected MGC80-3 cells. The proliferation was also remarkably decreased in Lv-shTPD52L2-infected SGC-7901 cells (p<0.001, Supplementary Fig. S1D), indicating that knockdown of TPD52L2 could significantly reduce the proliferation of gastric cancer cells. However, the inhibitory effect of Lv-shTPD52L2 on cell proliferation was weaker in MKN-28 and AGS cells with low TPD52L2 expression (Supplementary Fig. S1E, F).
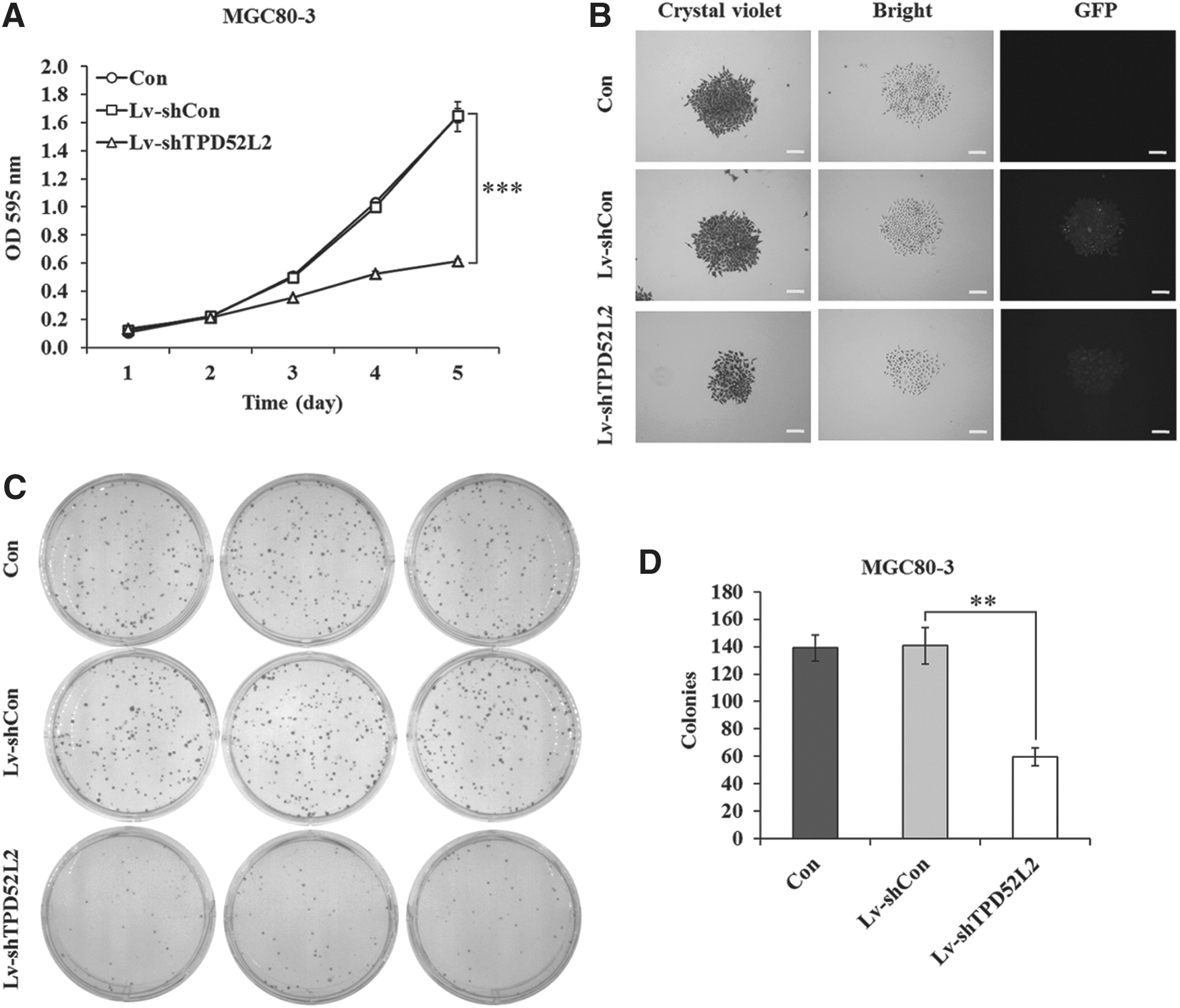
TPD52L2 knockdown inhibits cell proliferation and colony formation in MGC80-3 cells.
The authors further tested cell growth through colony formation analysis. Lv-shTPD52L2 group could strongly suppress the size of single colony (Fig. 2B) and the number of colonies (Fig. 2C, D). Their results demonstrate that TPD52L2 is essential for gastric cancer cell viability and proliferation.
Downregulation of TPD52L2 induces cell cycle arrest as well as apoptosis in MGC80-3 cells
A disregulation of the cell cycle components may lead to tumor formation. As extensively accepted, cancer cells always exert characteristic uncontrolled cell cycle. Many clinically used drugs for cancer therapy specially targeted cell cycle arrest, and this was considered as a promising lead to cancer therapeutics. The authors tended to test whether TPD52L2 knockdown could influence cell cycle distribution of MGC80-3 cells through FACS assay. They found that TPD52L2-silenced cells were blocked in the G0/G1 phase, compared with the Lv-shCon group (63.870%±1.215% vs. 52.720%±0.411%, p<0.01) (Fig. 3A, B). The cell populations of S and G2/M phases in the Lv-shTPD52L2 group were concomitantly reduced. Moreover, TPD52L2 knockdown in MGC80-3 cells also led to a significant accumulation in the sub-G1 phase representing apoptotic cells (Fig. 3C). Furthermore, knockdown of TPD52L2 caused an increase in the cleaved-PARP expression (Fig. 3D). Their data suggest that the knockdown of TPD52L2 in MGC80-3 cells leads to cell cycle arrest and apoptosis, which may contribute to gastric cancer cell proliferation inhibition.

Downregulation of TPD52L2 induces cell cycle arrest as well as apoptosis.
Discussion
TPD52L2, is commonly described as a protein involved in tumorigenesis, according to its name. TPD52L2 has been shown to be involved in several cancers, including breast cancer, ALL, and pancreatic cancer. However, the molecular basis for its oncogenic role is entirely unknown. For this purpose, they constructed shRNA-loaded lentivirus to knock down TPD52L2 expression in gastric cancer cells, and found that TPD52L2 silencing resulted in a remarkable alleviation in cell proliferation and colony formation ability. Regulation of the cell cycle involves processes crucial to the survival of a cell, including the detection and repair of genetic damage as well as the prevention of uncontrolled cell division. 15 Lv-shTPD52L2-infected cells were blocked in the G0/G1 phase, which is marked by the use of 20 amino acids to form millions of proteins and later on enzymes that are required in S phase, 16 suggesting the possible role of TPD52L2 in DNA synthesis preparation. Moreover, a significant accumulation of cells in the sub-G1 phase was observed after TPD52L2 knockdown, which is suggestive of apoptosis. 17 Furthermore, knockdown of TPD52L2 in MGC80-3 cells caused a cleavage of PARP, which is one of the most used diagnostic tools for the detection of apoptosis in many cell types expression. 18 Their study revealed that TPD52L2 is essential for gastric cancer cell proliferation, probably through regulation of cell cycle and apoptosis. This work was supported by a recent report showing that knockdown of TPD52L2 inhibited glioma cell proliferation and induced G0/G1 phase cell cycle arrest. 19
Current treatment of gastric cancer, including surgery, chemotherapy, and radiation has proved not much effective clinically. In recent years, antisense therapy has arisen as a promising therapy for many diseases. Its target was identified as the key genetic disorder responsible for particular diseases development. As of 2012, some 40 antisense oligonucleotides and small interfering RNAs (siRNAs) were in clinical trials, including over 20 in advanced clinical trials (phase II or III). 20 Many inappropriately expressed genes are reported in gastric cancer patients, including tumor suppressor genes and oncogenes. The replacement of inactivated tumor suppressor genes like p53 has emerged as an attractive treatment option, both alone and combined with conventional chemotherapy. 21 Approaches for the downregulation of tumor genes in gastric cancer have also been investigated and may be of potential clinical value in the gene therapy of gastric cancer. 22
Our work has illustrated that TPD52L2 is indispensable in gastric cancer proliferation. If this was further confirmed from in vivo research, antisense therapy targeting TPD52L2 will be promising. Furthermore, which process is exactly responsible for these events modulated by TPD52L2 is still to be investigated in a subsequent study.
Conclusions
In this study, the authors provide new evidence that TPD52L2 plays a critical role in gastric cancer growth in vitro. Their findings suggest that antisense therapy targeting TPD52L2 in gastric cancer may be of clinical value.
Footnotes
Acknowledgment
This work was supported by a grant from the Miaopu Project of the Second Military Medical University (No. 2014QN17).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.