
Abstract
In the treatment of B cell non-Hodgkin's lymphoma, rituximab is used in combination with different chemotherapeutics to improve its efficacy, but the mechanisms involved are not fully understood. The authors examined the mechanism by which rituximab combined with hydroxyurea or vincristine induces cell death in the human Burkitt's lymphoma Ramos cell line. Cell death was analyzed by phosphatidylserine exposure, caspase activation, and mitochondrial membrane changes. Their results indicate that the cell death initiated by the combination of rituximab and hydroxyurea is caspase-independent. In contrast, preincubation of the cells with the same concentrations of caspase inhibitors used with hydroxyurea eliminated the synergistic effect of the rituximab and vincristine combination. This was confirmed by the presence of the active fragment of caspase-3 in vincristine-treated cells. These preliminary results demonstrate that rituximab can activate different downstream signals to induce direct cell effects. Furthermore, the findings support the important role of mitochondria in the regulation of both pathways.
Introduction
Rituximab is a recombinant monoclonal antibody (mAb) constructed against the B cell antigen CD20. 1 This protein is a cell surface non-glycosylated phosphoprotein of 33–35 kDa that is expressed in a lineage-specific manner. 2,3 In normal B cells, its expression starts in the pre-B cell stage of development and continues until the antibody-producing stage. More importantly, this protein has been found on the surface of >95% of B lymphoma cells. 4,5 Rituximab is highly effective against B cell non-Hodgkin's lymphoma (NHL) and has improved the outcome of patients affected by different B cell malignancies. 6 However, 50% of B-NHL patients are unresponsive to rituximab or become resistant after therapy. 7 The killing mechanisms of rituximab have been extensively investigated. Complement-dependent cytotoxicity (CDC), antibody-dependent cell cytotoxicity (ADCC), and induction of direct cell death have all been claimed to be responsible for the efficacy of rituximab. 8 Although it is still difficult to evaluate, the contribution of each mechanism in vivo, their roles are influenced by lymphoma subtype and mode of resistance. However, in the absence of functional complement or accessory cells, rituximab can still activate intracellular signals that lead to apoptosis of lymphoma B cells. 9 –11 This notion seems acceptable because cells freshly isolated from rituximab-treated patients afflicted with chronic lymphocytic leukemia (CLL) show induction of apoptosis through activation of caspase-3, caspase-9, and PARP. 12 Although it is well established that caspases are the major drivers of rituximab-mediated apoptosis, several reports have indicated that the killing mechanism can proceed in the absence of caspases and in a highly regulated manner. 9,13 –15 Therefore, understanding the intracellular signals could permit designing new therapies that may overcome rituximab resistance.
Rituximab has been combined with chemotherapeutic agents to selectively regulate intracellular proteins that are involved in apoptotic cell death. Using this strategy, synergistic effects have been achieved by sensitizing chemotherapy-resistant malignant B cells to cell death. 10,16,17 The authors recently tested the effect of rituximab and chemotherapeutic combinations and found that rituximab interacts synergistically with vincristine and with hydroxyurea. 18 The results are interesting in view of the different mechanisms of cell death induced by these two drugs. Therefore, they made a preliminary study in the Ramos cell line of the cell death pathways involved in the synergistic effect of rituximab with hydroxyurea and with vincristine.
Materials and Methods
Antibodies and reagents
The chimeric anti-CD20 mAb rituximab (MabThera; Roche Diagnostics; stock solution 10 mg/mL) was a kind gift from the National Oncology Institute (Sabratah, Libya). All other reagents were purchased from Sigma Chemical Company, unless stated otherwise.
Cell lines
The CD20 positive Burkitt's lymphoma cell line (Ramos) was purchased from the European Collection of Cell Culture (ECACC) (Wiltshire SP4 0JK). Ramos cell lines show significant CD20 expression levels (468.4±14.0 MFI) as measured by mean fluorescence intensity in comparison to other rituximab-resistant cells.
19
Cells were maintained in RPMI-1640 medium supplemented with 10% heat-inactivated fetal bovine serum, 200 mM
Flow cytometry analysis
Before analysis, cells were manipulated into single-cell suspensions. Flow cytometry calibration, optimization, and compensation were performed using the FacsComp software (Becton Dickinson), whereas analysis and calculations were performed by CellQuest (Becton Dickinson). For each analysis, 20,000 events were collected.
Annexin V-based cell death analysis
Ramos cell lines were cultured at a density of 1×105 cells/mL and treated with 10 μg/mL rituximab alone or in combination with either 2 μM vincristine or 1 mM hydroxyurea. Following 24 and 48 hours of incubation, cell death was measured by staining with Annexin V and Propidium Iodide (PI) (BD Biosciences). Briefly, treated cells were washed in phosphate buffered saline, the supernatant was aspirated, and the cells were resuspended in Annexin V and PI incubation reagent (0.25 μg/mL Annexin V and 5 μg/mL PI in binding buffer). Cells were then incubated in the dark for 15 minutes at room temperature before analysis by flow cytometry (FacsCalibur; BD Biosciences) using CellQuest software. Annexin V has a high affinity for phosphatidylserine (PS), which is exposed on the extracellular leaflet of the cell membrane early in the apoptotic process. Using this method, three different populations of cells are distinguishable: live cells are negative for both Annexin V and PI, early apoptotic cells are positive for Annexin V and negative for PI, and late apoptotic/necrotic cells are positive for both Annexin V and PI. 20
Determination of caspase dependency of cell death
Caspase inhibitors were used to explore the caspase dependency of the cell death induced by rituximab and/or chemotherapies. Ramos cells at 1×105 cells/mL were pretreated for 30 minutes with 50 μM of either Z-VAD-FMK (pan caspase inhibitor), Z-DEVD-FMK (caspase-3 inhibitor), Z-IETD-FMK (caspase-8 inhibitor), Z-LEHD-FMK (caspase-9 inhibitor), or ZFA-FMK (caspase negative control) before treating them with rituximab and the chemotherapeutics. Following 24 and 48 hours of incubation, cell death was analyzed by using the Annexin V and PI method.
Active caspase-3 analysis
Caspase-3 was analyzed by flow cytometry using a FITC-conjugated Ab against its active fragment (BD Biosciences). Briefly, Ramos cells at 1×105 cells/mL were treated with rituximab and/or chemotherapeutic agents as described above. The cells were stained for intracellular active caspase-3 and analyzed by flow cytometry. As a negative control, the cells were stained with pure (isotype) IgG. Data were acquired by CellQuest software and the percentage of positive cells was recorded.
Mitochondrial membrane potential
To measure the mitochondrial membrane potential (ΔΨ), Ramos cells were treated as described above and stained according to the MitoProbeDiLC 1(5) Assay (Molecular Probes, Inc.). Cationic cyanine dyes accumulate in cells in response to the membrane potential changes that occur during cell death. DiLC 1(5) accumulates primarily in mitochondria with active membrane potentials. The intensity of the dye decreases when the mitochondrial membrane potential is disrupted. Carbonyl cyanide 3-chlorophenylhydrazone (CCCP), which is known to disrupt the mitochondrial membrane potential, was used as a positive control.
Statistical analysis
Values represent the means±standard error of the mean (SEM) of three separate experiments. Student's t-test was used for comparison of means and p-values<0.05 were considered significant.
Results
Caspase activation
The authors examined the caspase dependence of the observed cell death by using flow cytometry to analyze caspase-3 activation. They detected the presence of the active fragment of caspase-3 in cells treated with rituximab plus vincristine, but not in those treated with rituximab plus hydroxyurea (Fig. 1). This indicated the caspase dependence of the former combination, but not the latter. To confirm these results, they used the pan-caspase inhibitor Z-VAD. First, Z-VAD significantly reduced the percentage of Annexin V-positive cells observed after vincristine treatment from 25% to 12%, but this was not the case for hydroxyurea. Second, the pan-caspase inhibitor halved the percentage of Annexin V-positive cells seen after treatment with vincristine combined with rituximab from 63% to 31% (p<0.05). In contrast, it did not affect the percentage of death in cells treated with rituximab combined with hydroxyurea (38% and 36%, respectively) (Fig. 2). They conclude that the mechanism of cell death induced by a combination of rituximab and vincristine is caspase-dependent, whereas that of rituximab combined with hydroxyurea is caspase-independent. To determine which upstream caspases are involved in the synergistic effect of rituximab and vincristine, they treated Ramos cells with a combination of rituximab and vincristine in the absence and in the presence of specific inhibitors of caspase-8 (Z-IETD-FMK) and caspase-9 (Z-LEHD-FMK). Flow cytometric analysis (Fig. 3) showed that caspase-9 was activated by the combined treatment, but caspase-8 was not, demonstrating the involvement of caspase-9 in the killing of Ramos cells by the rituximab and vincristine combination.
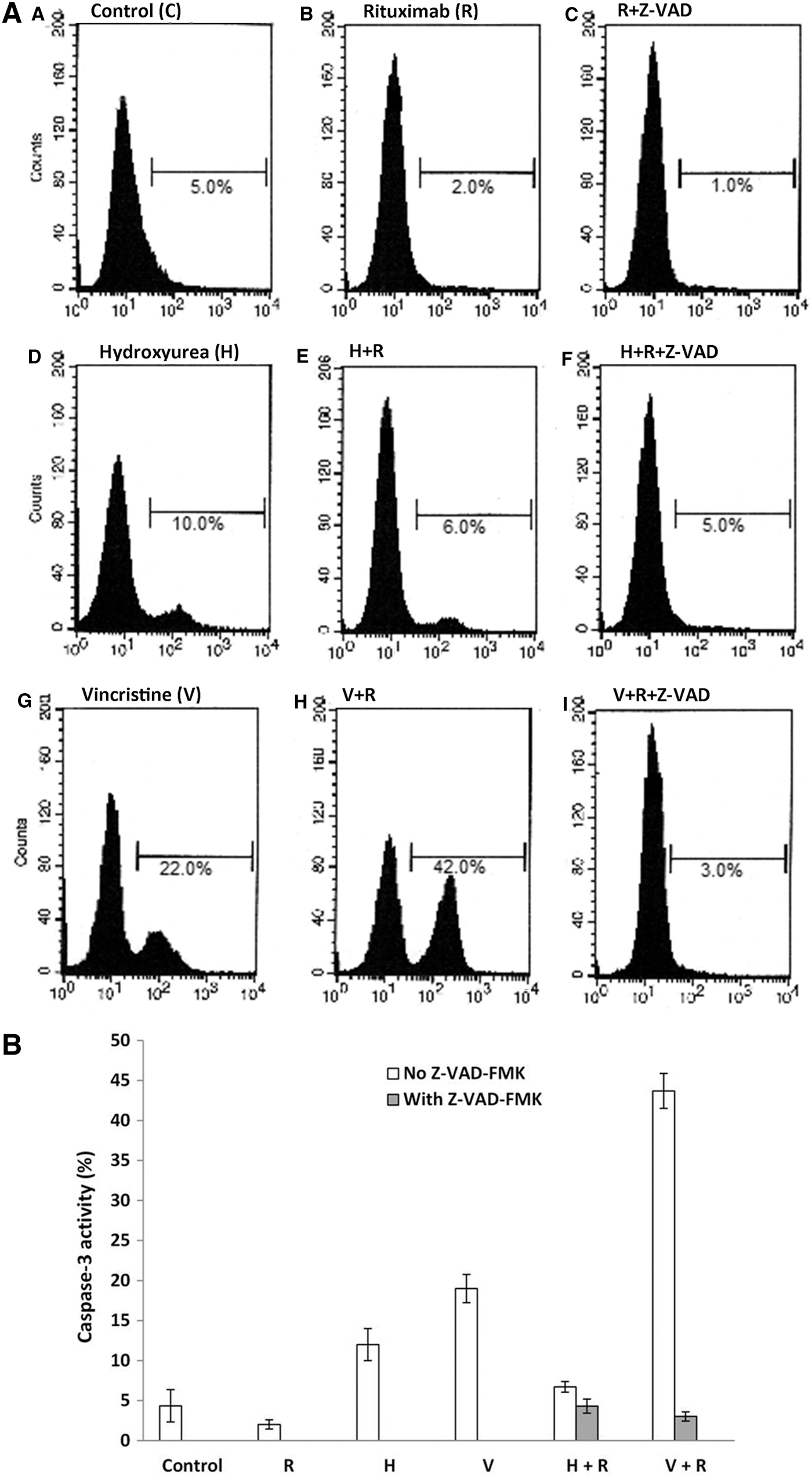
Flow cytometric analysis of caspase-3 activation by rituximab alone or in combination with chemotherapeutic agents. Ramos cells were treated with 10 μg/mL rituximab (R), 1 mM hydroxyurea (H), and 2 μM vincristine (V), each drug alone or in combinations as shown. Cells were stained with an antibody against the active caspase-3 fragment and analyzed by flow cytometry. An irrelevant antibody (C) was used as a negative control.

Combination of rituximab with either hydroxyurea or vincristine induced caspase-dependent and caspase-independent cell death, respectively. Ramos cells were pretreated with either 50 μM Z-VAD-FMK (closed bars) or appropriate vehicle (open bars) for 30 minutes before the addition of 10 μg/mL rituximab (R) alone or in combination with 1 mM hydroxyurea or 2 μM vincristine. After 48 hours, cell death was measured by flow cytometry using Annexin V. The mean percentage (±SEM) of Annexin V-positive cells is shown (n=3).

Flow cytometric analysis of the involvement of caspase-8 and caspase-9 in the apoptosis induced by the rituximab and vincristine combination. After different treatments, Ramos cells were stained with Annexin V and Propidium Iodide and analyzed by flow cytometry. Rituximab 10 μg/mL; vincristine 1 μM; caspase-8 inhibitor (Casp-8) (Z-IETD-FMK) 50 μM; caspase-9 (Casp-9) inhibitor (Z-LEHD-FMK) 50 μM.
Mitochondrial membrane changes
The authors measured the disturbance in mitochondrial membrane potential, which is an initial step in cells undergoing cell death. The percentage of disturbance in mitochondrial membrane potential following separate treatments with each drug was significantly increased (p<0.05) as compared with untreated cells (rituximab 22%, hydroxyurea 38%, vincristine 28%). When rituximab was combined with chemotherapeutic agents, the percentage of cells with a depolarized membrane increased to 51% for hydroxyurea augmentation and to 41% for vincristine addition (Fig. 4). Although treatment with rituximab combined with hydroxyurea or vincristine caused a greater disturbance in the mitochondrial membrane potential than the corresponding individual treatments, the combined effect was not significantly different from the mathematical sum of the individual treatments.
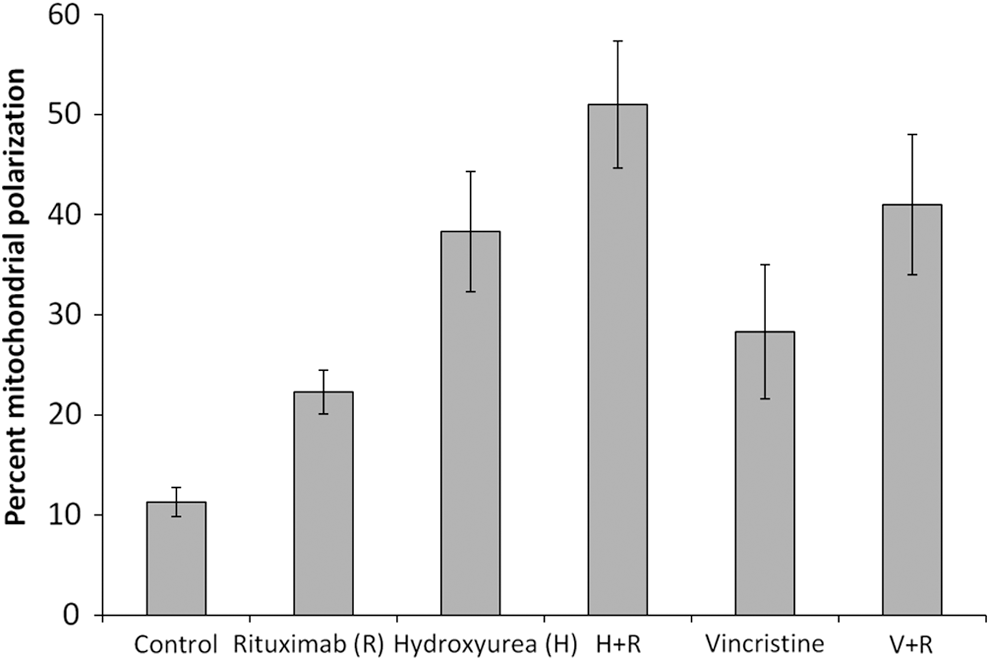
Ramos cells were incubated with 10 μg/mL rituximab (R), 1 mM hydroxyurea (H), and 2 μM vincristine (V), each drug alone or in combinations as indicated. The fraction of cells with dissipated mitochondrial transmembrane potential (Δψm) was determined by loss of staining with the fluorescent mitochondrial dye DiLC 1(5). The mean percentages (±SEM) of cells with depolarized membrane are shown (n=3).
Discussion
The clinical success of rituximab has encouraged investigators to study the mechanisms by which this Ab depletes lymphoma B cells. Although the in vivo mechanisms of action of rituximab are not fully understood, ADCC, CDC, and direct cell death are involved to some extent. 8 Accumulating data from different studies indicate that rituximab alone is not a strong inducer of cell death unless it is combined with other drugs. The authors recently demonstrated that treatment of the Ramos Burkitt's lymphoma cell line with a combination of rituximab and hydroxyurea or vincristine resulted in a synergistic effect that eliminated the majority of the tumor cells. 18 In the present study, they used Ramos cell lines to gain preliminary insights into the mechanisms of action by which rituximab increases the cytotoxicities of hydroxyurea and vincristine. In view of the current clinical use of Rituximab, human Burkitt's lymphoma cell lines are commonly used to investigate B cell depletion. They previously used a number of different Burkitt's lymphoma cell lines of type I and type III and found that Ramos type I are among the most responsive cells to rituximab, whereas type III are resistant. 9 So they excluded type III lines from their current study. Furthermore, Ramos cell lines were chosen for two other important reasons: they are Epstein-Barr Virus negative and they do not express the antiapoptotic protein Bcl-2. In addition to their direct clinical relevance, these cell lines offer the extra advantage that they can be induced to undergo classical, receptor-driven apoptosis by ligation of surface CD20, and are sensitive to both of the chemotherapeutic drugs they used. This makes it possible to directly compare the effects of rituximab and the chemotherapeutic drugs.
The chemotherapeutic drugs used in this study have different modes of action on cell cycle regulation and arrest cells in different cell cycle stages. Hydroxyurea reduces the proliferative capacity of cells and induces senescence-like changes, and eventually arrest at G1, 21,22 whereas vincristine disrupts cellular microtubules and prevents the formation of a functional spindle, resulting in the arrest of cells at mitosis. 23 However, the authors' results indicate that the difference in cell death between the chemotherapeutic drugs that act on different phases of the cell cycle is negligible, and that rituximab is very potent at inducing cell death when combined with either hydroxyurea or vincristine.
Combining rituximab with hydroxyurea induced cell death in the absence of caspase activation. This is in agreement with a recent study showing that hydroxyurea potentiates the killing effect of rituximab in a caspase-independent manner. 24 Nevertheless, rituximab plus vincristine were synergistic in Ramos cell lines although their signaling pathways are different. They examined the involvement of caspases by using the pan-caspase inhibitor Z-VAD-FMK, which blocked the synergistic effect of the rituximab+vincristine combination, but not the synergy of the rituximab+hydroxyurea combination. The fact that Z-VAD-FMK was able to block vincristine-induced apoptosis confirms its ability to cross the plasma membrane and reach the appropriate intracellular sites for caspase inhibition. In addition, vincristine-induced caspase-dependent cell death indicates that the intracellular signaling pathway leading Ramos cell lines to apoptosis is intact. These results were also confirmed by detecting the active fragment of caspase-3.
The simultaneous use of rituximab with either hydroxyurea or vincristine does not seem to alter the mechanism by which each individual chemotherapeutic drug induces cell death. Rituximab alone can kill cells in the absence of caspase activation, but it still acts synergistically with vincristine, which acts by a mechanism involving caspase activation. Interestingly, it was recently suggested that caspase-dependent and caspase-independent cell death can be coordinated in the same cells by a single upstream transcription factor through a bivalent regulatory node. 25 In addition, rituximab has been shown to block cell survival signaling pathways and sensitize cells to the effect of cytotoxic drugs. 10,17,26 Sensitization can also be mediated by the downregulation of antiapoptotic proteins. 16,26 Following sensitization, rituximab activates cells to either Fas-dependent cell death 27 or the mitochondrial killing pathway. 28 Indeed, the upstream caspase-8 activates the death receptor pathway, whereas caspase-9 activates the mitochondrial pathway. To determine which of these two upstream caspases was involved in rituximab- and vincristine-induced apoptosis, they used selective inhibitors of caspase-8 and caspase-9. The authors' results demonstrate that apoptosis induced by the rituximab+vincristine combination was completely blocked by the casapse-9 inhibitor, but not by the caspase-8 inhibitor. Activation of both caspase-9 and caspase-3 together with mitochondrial depolarization and PS exposure suggest that the rituximab and vincristine combination may activate the intrinsic pathway. In harmony with this, rituximab has been suggested to modulate intracellular proteins and induce synergistic cell death with paclitaxel (Taxol) 29 and with lenalidomide 30 using the mitochondrial pathway. Additionally, freshly isolated cells from rituximab-treated CLL patients show activation of caspase-3 and caspase-9, but not caspase-8, the activation of which is controlled by the death receptor pathway. 12 In addition to modulating intracellular proteins, rituximab as a single agent can induce direct cell death. 11,12,31,32 The authors and others have demonstrated that caspase inhibitors cannot block rituximab-induced cell death, indicating that rituximab can regulate downstream signaling and bypass caspase activation. 9,15 Therefore, the synergistic effect of rituximab and hydroxyurea, a caspase-independent inducer, 24 would imply that the intracellular events triggered by the two agents may share a common pathway that can be mutually enhanced by the addition of one drug to another. The same results have been observed when rituximab was combined with the caspase-independent cell death inducer, Milatuzumab. 33
In this context, rituximab has also been shown to sensitize cells to chemotherapeutic drugs by triggering different intracellular signals known to block various survival signaling pathways. This is achieved, in part, by downregulating the antiapoptotic Bcl-2 proteins, such as Bcl-2 and Bcl-xl (reviewed in Abulayha et al. 34 ). Since Ramos cells are Bcl-2 negative, studying Bcl-xl and its linked survival signals may give a clear understanding of the events involved in the killing mechanisms of the combinations used in this study. However, the mechanisms by which rituximab induces sensitization might be different from those involved in rituximab-induced cell death when the antibody is used as a single agent, and this has not yet been clarified.
Mitochondrial membrane potential changes have been proposed to be among the obligate events in the different apoptotic signaling pathways because mitochondrial markers relating to both caspase-dependent 15 and independent cell death 35 are involved in rituximab-induced cell death. These observations support the dominant role of mitochondrial events in the killing mechanism of rituximab. Further research is required to clearly establish a mechanistic link between the caspase-dependent and independent pathways, and to identify the most relevant apoptotic mediators involved in both systems.
The results of the authors' study extend the argument that rituximab can induce direct cell death through different downstream signals, and switching between these pathways may rely on the mode of upstream binding. This concept is supported by experiments in which the B cell receptor (BCR) in Burkitt's lymphoma cell lines was triggered by different modes of binding. The results showed that caspase-mediated apoptosis is induced following hyper crosslinking of the BCR, whereas binding of soluble Abs induces caspase-independent cell death. 36
Understanding the biochemical pathway and identifying targets that control the response to rituximab treatment may permit optimization of combination therapeutic interventions to overcome the resistant phenotypes of some NHL patients. Furthermore, the authors' in vitro findings require further studies to confirm whether these important effects also occur in the in vivo setting.
Footnotes
Acknowledgments
This work was funded by the Libyan Authority for Research, Science, and Technology, Tripoli, Libya. The authors thank Dr. Amin Bredan for his kind assistance in the preparation of the article.
Disclosure Statement
The authors declare that they have no conflict of interest.