
Abstract
Cancer Biotherapy and Radiopharmaceuticals
officially retracts the article entitled, “Tectonic 1 Is a Key Regulator of Cell Proliferation in Pancreatic Cancer,” by Shiyong Zhao, Xuedong Chen, Ming Wan, Xingming Jiang, Chunlong Li, Yunfu Cui, and Pengcheng Kang, (Cancer Biother Radiopharm 2016;31(1):7–13; doi: 10.1089/cbr.2014.1778) due to incorrectly attributed or identified sequences. Several figures and text within the methods section are strikingly similar to those in other publications1–3 and suggests that same party prepared these papers for entirely different author sets.
The Editor and Publisher of Cancer Biotherapy and Radiopharmaceuticals are committed to preserving the scientific literature and the community it serves.
References
1. Wang Z, Xu D, Gao Y, et al. Immature colon carcinoma transcript 1 is essential for prostate cancer cell viability and proliferation. Cancer Biother Radiopharm 2015;30(7):278–284; doi: 10.1089/cbr.2014.1728.
2. Zhang C, Zhang T, Song E, et al. Ribosomal protein S15A augments human osteosarcoma xell proliferation in vitro. Cancer Biother Radiopharm 2018;29(10):451–456; doi: 10.1089/cbr.2014.1698.
3. Wang H, Wang B, Zhu W, et al. Lentivirus-nediated knockdown of myosin VI inhibits cell proliferation of breast cancer cell. Cancer Biother Radiopharm 2015;30(8):330–335; doi: 10.1089/cbr.2014.1759.
Introduction
The pancreas is a glandular organ in the digestive system and endocrine system of vertebrates. It functions as an endocrine gland producing several important hormones, including insulin, glucagon, somatostatin, and pancreatic polypeptide, which circulate in the blood. The pancreas is also a digestive organ, secreting pancreatic juice containing digestive enzymes that assist digestion and absorption of nutrients in the small intestine. 1 Dysfunction of pancreas is always disastrous.
Pancreatic cancer is considered as one of the most vicious human cancers. In 2012, pancreatic cancer caused 330,000 deaths globally, the seventh most common cause of death due to cancer. Symptoms that are specific enough to suspect pancreatic cancer often do not appear until the disease is already in an advanced stage. By the time of diagnosis, the cancer has usually spread to other parts of the body. This is the cause of difficulties in clinical treatment. 2,3 Thus, diagnosis in the early stage is extremely important for following treatment. CA19-9 (carbohydrate antigen 19.9) is a tumor marker that is frequently elevated in pancreatic cancer, but lacks sensitivity and specificity. 4,5 More specific biomarkers remain to be identified.
Tectonic 1 (TCTN1) belongs to a family of signal-sequence-containing secreted and transmembrane proteins evolutionarily conserved among eukaryotes. 6,7 Multiple transcript variants encoding different isoforms have been found for this gene. TCTN1 is essential for preventing diffusion of transmembrane proteins between the cilia and plasma membranes, through forming a tectonic-like complex localized at the transition zone of primary cilia. 8 As a regulator of the Hedgehog (Hh) pathway, TCTN1 is required for both activation and inhibition of the Hh pathway in the patterning of the neural tube. The Hh pathway is a signaling pathway that transmits information to embryonic cells required for proper development. 9 However, its role in tumor progression has received little attention.
In this study, the authors utilized a lentivirus system to knock down TCTN1 expression in PACN-1 pancreatic cancer cells, tending to illustrate the interplay between TCTN1 and pancreatic cancer development.
Materials and Methods
Cell culture
Human pancreatic carcinoma, epithelial-like cell line PANC-1, ASPC-1, BXPC-3, and human embryonic kidney cell line HEK293T were obtained from Cell Bank of Shanghai Institute of Cell Biology, Chinese Academy of Sciences (Shanghai, China). HEK293T was employed for lentivirus package. Both cells were cultured in Dulbecco's modified Eagle's medium (SH30243.01B+; Hyclone) with 10% fetal bovine serum (S1810; Biowest). All cell lines were incubated at 37°C with a humidified atmosphere (95% air, 5% CO2).
Construction of recombinant lentivirus
Recombinant lentiviral vector was constructed as described. 10 The short hairpin RNA (shRNA) sequence (5′-GCTCAGATGCATCAGTTCCTTCTCGAGAAGGAACTGATGCATCTGAGCTTTTTT-3′) specific for TCTN1 knockdown was inserted into the lentiviral expression vector pFH-L (Shanghai Hollybio), which contains a green fluorescent protein (GFP). The shRNA-containing scrambled candidate sequence of TCTN1 complementary DNA (cDNA) (5′-GCGGAGGGTTTGAAAGAATATCTCGAGATATTCTTTCAAACCCTCCGCTTTTTT-3′) was used as control. shRNA was generated by annealing the synthetic DNA oligonucleotide primers and inserted into lentiviral expression vector pFH-L (Shanghai Hollybio), which contains a GFP gene as a reporter gene activated by CMV promoter.
Recombinant lentiviral vector was produced by cotransfection of 50% confluence HEK293T with reconstructive vector pFH-L and packing plasmids, pVSVG-I and pCMVΔR8.92 (Shanghai Hollybio), mixing Lipofectamine 2000. Lentiviral particles were harvested after 2 days, centrifuged to get rid of cell debris, and filtered through 0.45-μm filters.
Recombinant lentivirus transfection in PANC-1 cells
For lentivirus infection, PANC-1 cells were seeded at 5 × 104 cells per well into a six-well culture plate. When grown to the density of 30% confluence, cells were infected with lentivirus-containing sh-TCTN1 (Lv-shTCTN1) or control lentivirus (Lv-shCon) at a multiplicity of infection of 75. After 96 hours of infection, cells were harvested and transduction efficiency was evaluated by GFP-positive cells.
RNA extraction and quantitative real-time polymerase chain reaction analysis
After 6 days of infection, total RNA was isolated from PANC-1 cells. Five micrograms RNA from total isolated RNA was used to synthesize the first strand of cDNA using SuperScript II RT 200 U/mL (Invitrogen). TCTN1 messenger RNA (mRNA) expression was evaluated by RT-qPCR on the BioRad Connet Real-Time PCR platform with SYBR Green PCR core reagents (reaction system: 2× SYBR premix ex taq 10 μL, forward and reverse primers (2.5 μM) 0.8 μL, cDNA 5 μL, ddH2O 4.2 μL). After initial denaturation at 95°C for 1 minute, a total of 40 cycles (denaturation 95°C, 5 seconds; annealing extension at 60°C, 20 seconds) proceeded, and the absorbance was obtained at the extension stage. Actin was used as the internal reference control. The polymerase chain reaction (PCR) primers were as follows: TCTN1-F: CCTTTGCGTGAATGTTGTTC, TCTN1-R: AGAGGGACTGGCTGGGTATT; actin-F: GTGGACATCCGCAAAGAC, actin-R: AAAGGGTGTAACGCAACTA. Data were analyzed using the 2−ΔΔCt statistical method. Results are presented as CT values, which were defined as the threshold PCR cycle number at which an amplified product is first detected. The average CT was calculated for both TCTN1 and actin, and ΔCT was determined as the ratio of the mean of the triplicate CT values for TCTN1 to the mean of the triplicate CT values for actin.
Western blot analysis
PANC-1 cells were collected at 5 days postlentivirus infection. After incubation in RIPA buffer, the cell lysis was centrifuged at 15,000 g at 4°C for 10 minutes. The supernatant was boiled after adding 2× sodium dodecyl sulfate (SDS) sample buffer (100 mM Tris-Hcl [pH 6.8], 10 mM EDTA, 4% SDS, 10% Glycine). Proteins were separated by 10% SDS-polyacrylamide gel electrophoresis (PAGE) gels and electrotransferred onto polyvinylidene difluoride membranes (Millipore). After blocking with 5% milk, the membranes were probed overnight with primary antibodies, rabbit anti-TCTN1 (1:5000, Ab-326; Proteintech), rabbit anti-Bax (1:1000, No. 2774; Cell Signaling), rabbit anti-Bcl-2 (1:1000, Ab-184; Proteintech), mouse anti-cdc2 (1:1000, No. 9116; Cell Signaling), rabbit anti-p21 (1:1000, No. 2947; Cell Signaling), rabbit anti-p27 (1:1000, No. 3686; Cell Signaling), rabbit anti-PARP (1:1000, No. 9542; Cell Signaling), and mouse anti-β-actin (1:50,000, Ab-55; Proteintech). The membrane was washed and then incubated with horseradish peroxidase-conjugated secondary goat anti-rabbit antibodies (1:5000, SC-2054; Santa Cruz) for 1 hour. Target proteins were detected with enhanced chemiluminescence reagents (Amersham) and photographed.
MTT cell viability assay
PANC-1 cells from different groups were seeded at an initial density of 2 × 103 cells/mL in 96-well plates for different times as the experiment required. After 96 hours of incubation, 3-(4, 5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) stocked at 5 mg/mL in phosphate-buffered saline (PBS) (pH 7.4) was added into each well at a final concentration of 100 μg/mL for 4 hours. Acidic isopropanol (10% SDS, 5% isopropanol, and 0.01 M HCl) was then added to stop the reaction and an enzyme-linked immunosorbent assay reader was used to measure the absorbance (Bio-Rad) at a wavelength of 595 nm. Viability of cells was expressed relative to theoretical absorbance.
Colony formation
To assay monolayer colony formation, 500 PANC-1 cells from different groups after 96 hours of infection were plated in six-well plates. After 7 days, cells were fixed with methanol and stained with crystal purple. Cells were indicated by the number of colonies.
Cell cycle analysis
The cell cycle distribution was analyzed using flow cytometry with propidium iodide (PI) staining. In brief, PANC-1 cells infected with the lentivirus constructs for 5 days were seeded in 6-cm dishes (2 × 105 cells/dish) and cultured for 40 hours at 37°C, respectively. On reaching a confluence of 80%, cells were harvested after trypsinization, washed with PBS, and fixed with 70% cold ethanol. The fixed samples were collected by centrifugation and resuspended in PI/RNase/PBS (100 μg/mL PI and 10 μg/mL RNase A) solution for 30 minutes at room temperature in the dark. The suspension was filtered through a 50-mm nylon mesh, and the DNA content of stained nuclei by PI was analyzed by a flow cytometer (FACS Calibur; BD Biosciences).
Apoptosis analysis by flow cytometry
PANC-1 cells were divided into two groups with different treatments (Lv-shTCTN1 and Lv-shCon). After 72 hours of incubation, cells were washed and reseeded in 6-cm dishes at a density of 80,000 cells/dish before analysis. After 24 hours, the cells were collected and Annexin V-APC/7-AAD double staining was performed according to the manufacturer's instructions (KeyGEN Biotech). Flow cytometry analysis was carried out on the FACS Caliber II sorter and Cell Quest FACS system (BD Biosciences).
Statistical analysis
Statistical analysis was carried on by using SPSS19.0 (IBM). The differences between groups were compared using Student's t-test, and data are expressed as mean ± standard deviation. Statistically significant difference was accepted as p < 0.05. Each experiment was performed in triplicate.
Results
Lentivirus-mediated RNA interference downregulated TCTN1 expression
As shown in Figure 1A, TCTN1 was highly expressed in three pancreatic cancer cells. The development of lentiviral systems to mediate RNA interference (RNAi) in primary neurons has facilitated physiological and biomedical research. 10 –12 Thus, the authors selected a 19 nt sequence specific for TCTN1 to construct lentivirus-expressed shRNA to downregulate TCTN1 expression in PANC-1 cells. The infection efficiency was significantly high as evidenced by the GFP reporter gene (Fig. 1B), indicating that shRNA directed by lentivirus was highly expressed in cells. Quantitative real-time PCR showed that TCTN1 mRNA was remarkably decreased in PANC-1 cells, attaining a knocking down efficiency as high as 93.8% (Fig. 1C). Therefore, the authors confirmed that lentivirus-mediated RNAi successfully knocked down TCTN1 expression in PANC-1 cells.
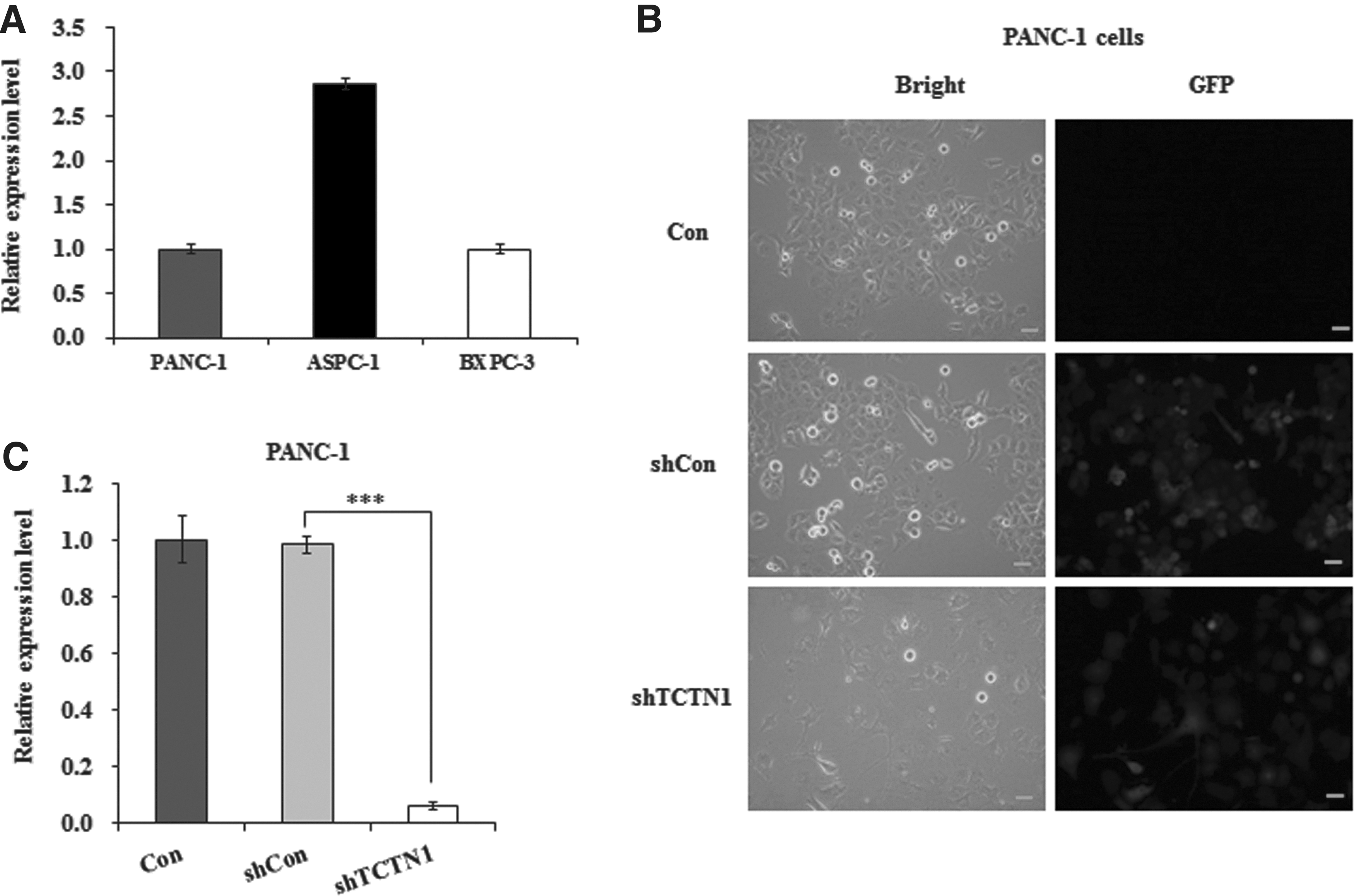
Knockdown of TCTN1 through lentivirus-mediated RNA interference.
Decreased TCTN1 by lentivirus-mediated knockdown leads to cell viability inhibition in PANC-1
To elucidate the effect of TCTN1 on malignant phenotype in pancreatic cancer, the authors examined the cell proliferation rate of Lv-shTCTN1-infected PANC-1 cells by MTT assay at indicated times. Compared with controls, the growth rate after 4 or 5 days was strongly inhibited in TCTN1 knockdown cells, respectively (0.235 ± 0.005 vs. 0.711 ± 0.038, p < 0.001; 0.221 ± 0.007 vs. 1.065 ± 0.061, p < 0.001) (Fig. 2A), while there was no significant difference between Lv-shCon and Con groups, suggesting that the specific inhibition of cell proliferation was attributable to specific knockdown of TCTN1.
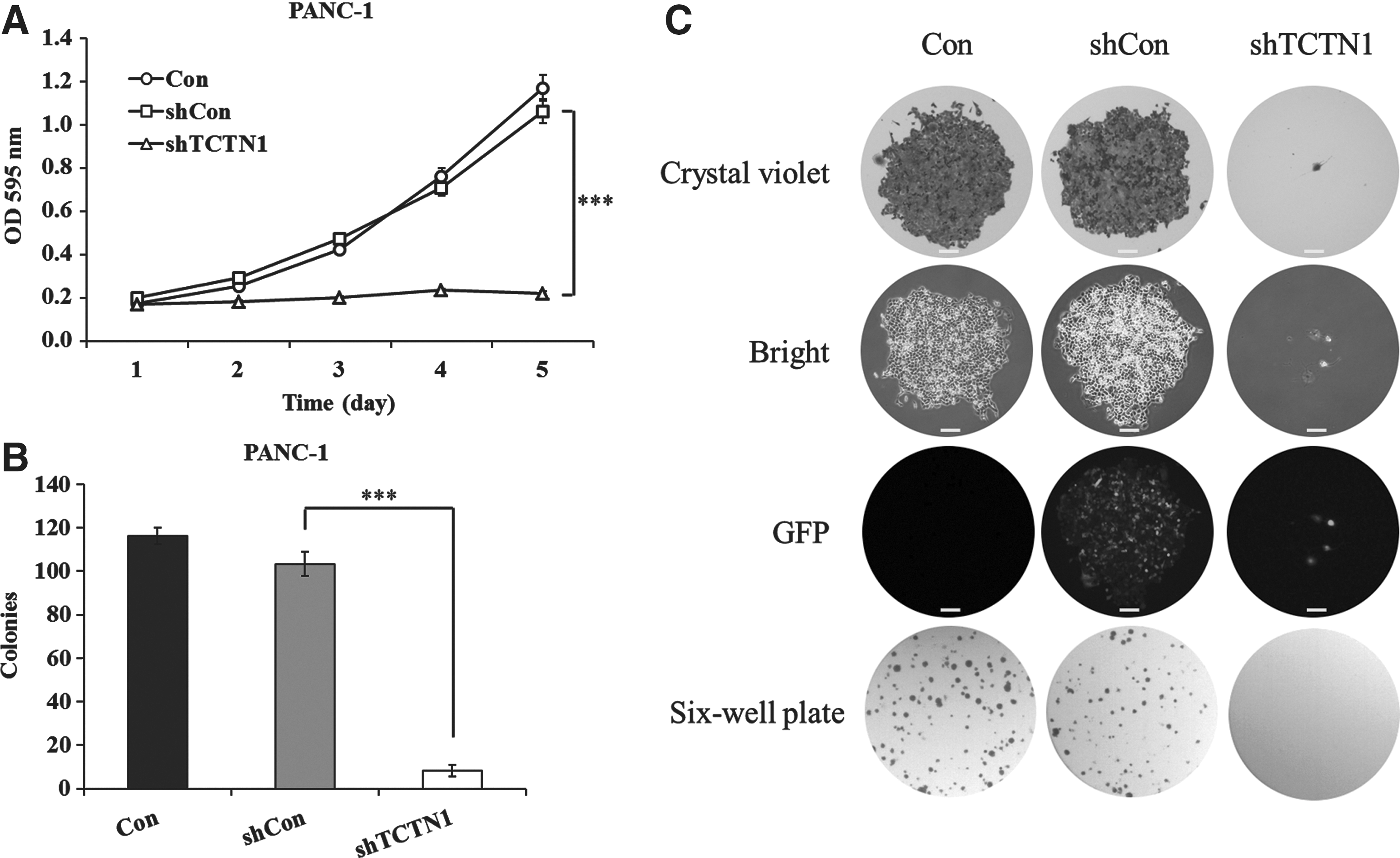
Effects of TCTN1 knockdown on PANC-1 cell proliferation and colony formation.
Meanwhile, the authors performed colony formation assay in monolayer culture. Images recorded by the fluorescence microscope showed a significant decrease in growth in knockdown cells (Fig. 2B). Statistical analysis of the number of colonies of PANC-1 cells revealed that the number of surviving colonies in Lv-shTCTN1-infected cells was much fewer than those of the Lv-shCon group (p < 0.001) (Fig. 2C). The data demonstrated that knockdown of TCTN1 in pancreatic cancer cells decreased its ability of colony formation. MTT and colony formation assay both suggested that TCTN1 was indispensable for PANC-1 cell viability.
Downregulation of TCTN1 induces PANC-1 cell cycle arrest in G2/M phase and cell apoptosis
A dysregulation of the cell cycle components may lead to tumor formation. As extensively accepted, cancer cells always exert a characteristic uncontrolled cell cycle.
Clinical treatments aiming at cell cycle inhibition were extensively taken as effective antitumor strategies. Thus, the authors tended to test whether TCTN1 knockdown could influence cell cycle distribution of PANC-1 cells through fluorescence-activated cell sorting assay. It was found that the population of the cells in the G2/M phase was obviously different among tested groups. The ratio of G2/M phase in the TCTN1 knockdown cells was dramatically higher compared with Lv-shCon group cells (31.24 ± 2.75 vs. 19.59 ± 0.27, p < 0.05), whereas the cell percentage in the S phase was lower (15.19 ± 3.04 vs. 23.77 ± 0.36, p < 0.05), suggesting that TCTN1-silenced cells were blocked in the G2/M phase (Fig. 3A, B). In accordance with it, compared with that in the Con and shCon groups, the expression of cdc2 decreased in Lv-shTCTN1-infected cells, while the expression of p21 and p27 increased and detected by western blotting (Fig. 3C). The results suggest that TCTN1 knockdown inhibited pancreatic cancer cell growth possibly by inducing cell cycle arrest.

Downregulation of TCTN1 induces cell cycle arrest.
Further analysis of sub-G1 phase representing apoptosis showed that TCTN1 knockdown could induce extensive apoptosis in PANC-1 cells (9.18 ± 0.24 vs. 0.28 ± 0.04, p < 0.001) (Fig. 4A). To further confirm the effect of TCTN1 on cell apoptosis, the authors then applied Annexin V-APC/7-AAD double staining on infected PANC-1 cells. Annexin V-APC versus 7-AAD plots from the gated cells showed four types of cells, viable (Annexin V−/AAD−), necrotic (Annexin V−/7-AAD+), early apoptotic (Annexin V+/7-AAD−), and late apoptotic (Annexin V+/7-AAD+). The knockdown of TCTN1 induced a much higher proportion of apoptotic cells (early apoptosis and late apoptosis) compared with controls (p < 0.001) (Fig. 4B, C). Moreover, the western blotting analysis demonstrated that the expression of Bax and cleavage of PARP-1 increased in Lv-shTCTN1-infected cells, while the expression of Bcl-2 decreased compared with that in the Con and shCon groups. So, all the results proved that knockdown of TCTN1 led to a strong proapoptotic effect in human pancreatic cancer PANC-1 cells.
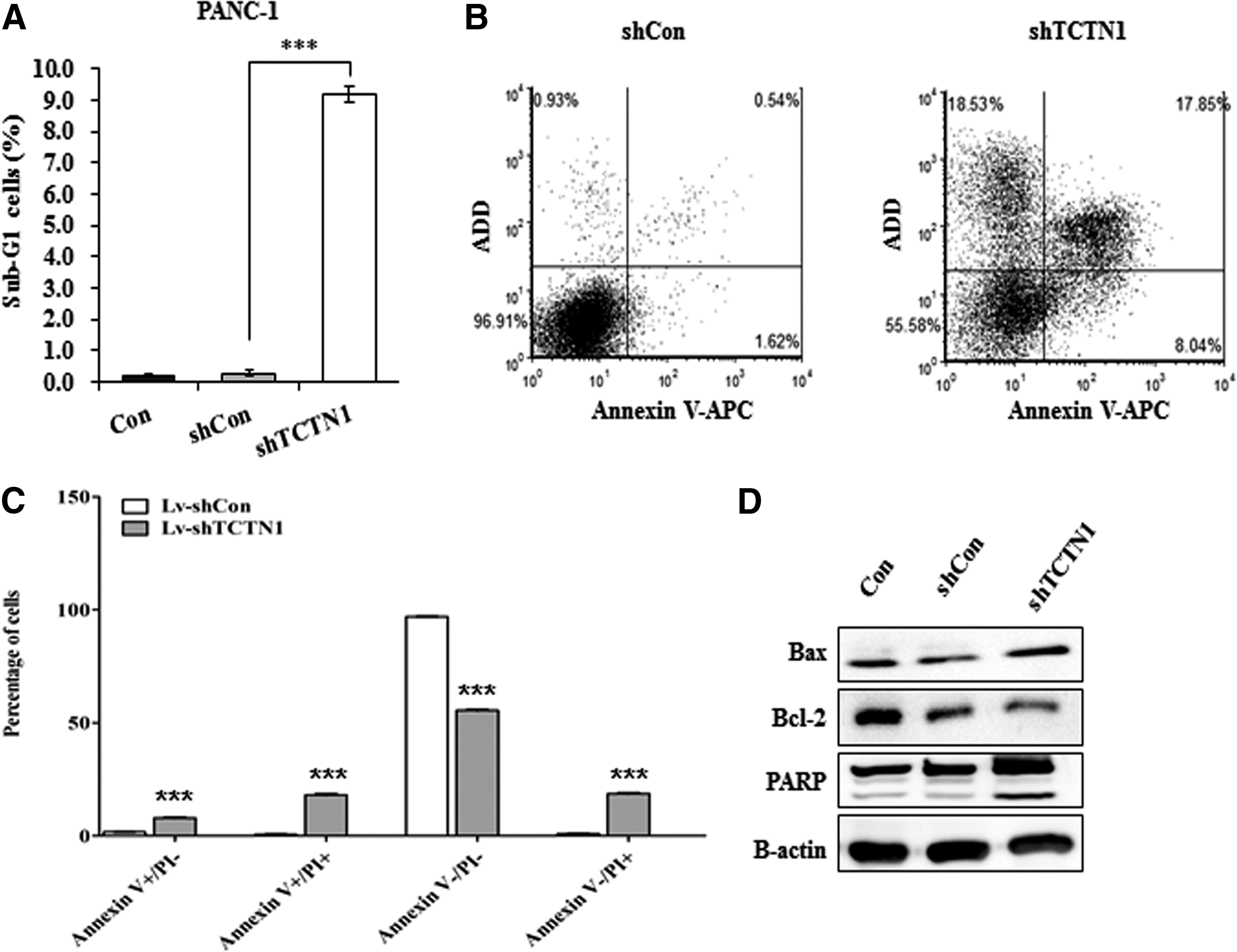
Downregulation of TCTN1 induces PANC-1 cell apoptosis.
Discussion
Early pancreatic cancer usually does not cause symptoms, so the disease is typically not diagnosed until it has spread beyond the pancreas itself. 13 Trails in searching specific biochemical markers in diagnosis have never stopped. In recent years, antisense therapy has arisen as a promising therapy for many diseases, including cancer. When the genetic sequence of a particular gene is known to be causative of a particular disease, it is possible to synthesize a strand of nucleic acid that will bind to the mRNA produced by that gene and inactivate it, effectively turning that gene off. 14
TCTN1 is a member of the tectonic family of secreted and transmembrane proteins. Current studies about this gene have already revealed its role in ciliogenesis and patterning of the neural tube as described above. Ciliary dysfunction underlies a group of diseases, including Joubert syndrome (JBTS), considered to be caused by abnormal mutation of the TCTN1 gene. 15,16 A mutation in TCTN1 that causes JBTS was identified. 8 What is more, multigene panel next-generation sequencing in patients with JBTS revealed the relationship between phenotypes and sequence variants in TCTN1 genes. 17 TCTN1 was expressed at different levels in most human tissues, including pancreas; accurate modulation of this gene is particularly essential. Evidences have already suggested that dysfunction of this gene is related with human diseases such as cancer, although the molecular basis for its role in physiological processes is entirely unknown.
For this purpose, the authors focused on its role in pancreatic cancer development. They constructed shRNA-loaded lentivirus to knock down TCTN1 expression in pancreatic cancer cell line PANC-1 and found that downregulation of TCTN1 in PANC-1 cells induced significant inhibition of cell viability as well as cell cycle arrest in the G2/M phase. Cell proliferation proceeds as an orderly progression through the cell cycle, which is governed by protein complexes comprising cyclins and cyclin-dependent kinases (CDKs). 18 Besides, it has been shown that some CDK inhibitors also play an important role in the cell cycle. Among them, p21 and p27 are members of the CIP/KIP protein family, which could suppress a relatively broad range of kinases, including cdk4 and cdk2. 19 In this study, the authors found that compared with that in the Con and shCon groups, the expression of cdc2 decreased in Lv-shTCTN1-infected cells, while the expression of p21 and p27 increased, which suggests knockdown of TCTN1-induced cell arrest through the inhibition of cyclins and CDKs, as well as the induction of CDK inhibitors.
Sub-G1 analysis further evidenced that decrease of TCTN1 led to extensive cell death of pancreatic cancer cell, PANC-1. The Annexin V test was also carried out to prove the stronger cell apoptosis caused by the knockdown of TCTCN1 in PANC-1 cells. The Bcl2 family protein includes proapoptotic (Bax, Bid, Bik, Bak) and antiapoptotic (Bcl2, BclX) proteins. The activation of the proapoptotic members leads to their translocation to the mitochondria 20 where they interact with antiapoptotic members and induce cytochrome c release and the loss of mitochondrial potential, thus inducing cell apoptosis. 21 However, the cell apoptosis process can be blocked by the overexpression of Bcl-2. 22,23 After cytochrome c release, the activity of caspase is activated, leading to the cleavage of poly (ADP-ribosylation) enzyme, PARP-1. In this study, the authors detected that the expression of Bax and cleavage of PARP-1 increased in Lv-shTCTN1-infected cells, while the expression of Bcl-2 decreased, which suggests that knockdown of TCTN1 induces cell apoptosis in the mitochondria-dependent pathway.
These results indicated that TCTN1 was probably a bona fide oncogene in pancreatic cancer. In the following study, if TCTN1 is proved to be the pivotal factor in pancreatic cancer development and progression, antisense therapy targeting TCTN1 seems to be an exciting story. Although comprehensive and detailed function is still to be obtained through further research, the authors have identified a novel potential oncogene, TCTN1, in pancreatic cancer as a promising target for diagnosis and clinical treatment.
Footnotes
Acknowledgment
The authors are thankful for the support of Department of Science Research Project in Heilongjiang Province (No. GC12C304-1).
Disclosure Statement
No competing financial interests exist.