
Abstract
C3Hf/HZgr mice were transplanted with SCCVII carcinoma cells and treated with Newcastle disease virus (NDV). The treatment slows down the growth of transplanted tumor. Furthermore, by using specific monoclonal antibodies, the frequencies of CD4+, CD8+, and CD4+CD25+ T lymphocytes were determined in the spleen of tumorous mice at particular times following tumor transplantation and/or NDV application. The incidence of lymphocytes CD4+ and CD8+ decreased and of CD4+CD25+ increased in the spleen of mice during the time following tumor transplantation. However, the frequency of regulatory CD4+CD25+ T lymphocytes in the spleen is very low, while CD4+ and CD8+ increased to normal level following intraperitoneal (i.p.) NDV injection in tumor-bearing mice. Thus, besides directly destroying transplanted tumor, NDV seems to be involved against growing tumor by reducing the frequency of regulatory T lymphocytes maintaining the frequency of CD4+ and CD8+ T lymphocytes within the control values pointing to its role in immunomodulation.
Introduction
Virus activities against tumor are well recognized. 1,2 Besides direct selective destruction of tumor cells by particular viruses, their immunomodulating activities in tumor-bearing organism have also been considered. 2,3 The Newcastle disease virus (NDV), like other viruses, infects host cells resulting in prominent virus replication, which is more expressed in tumor than in normal cells, leading to pronounced oncolysis. 4 Actually, NDV replication is over 10,000 times more pronounced in human malignant than in normal cells. 5,6
Besides pronounced oncolytic activities, NDV modulates either an innate (oncolytically derived tumor cell fragments initiate activities of dendritic cells and macrophages, as well as induce chemokines and cytokines) or acquired (T-lymphocyte stimulation by pronounced antigen presentation) immune response. 7,8
There are numerous mechanisms involved in tumor progression, such as angiogenesis induction and particularly immune suppression. Tumor-induced immunosuppressive mechanisms are part of physiologic regulatory mechanisms responsible to prevent autoimmunity with T-regulatory (Treg) lymphocytes as pivotal mediators. 9 T-regulatory lymphocytes characterized as CD4+CD25+ cells play an essential role in controlling T-cell numbers neutralizing autoreactive T cells. 10 Treg constitute 5%–10% of peripheral CD4+ T lymphocytes in normal mice, but it has been shown that a tumor can differentiate, expand, recruit, and activate these cells. 11 –13
Furthermore, the incidence of Treg cells is commonly elevated in tumors, in peripheral blood, or in lymphoid organs of tumor-bearing hosts. 12 These Treg cells, known as tumor Treg, 12,13 may promote local tumor growth at the tumor site and may be relevant for the progression of systemic disease. 14 Treg suppresses CD4+ and CD8+ T cells, B cells, NK cells, dendritic cells, and macrophages, but cell targets and stages of the immune response critical for this suppression are still unclear. 12,13,15,16 In the spleen of tumor-bearing organisms decreased incidence of CD4+ and CD8+ T lymphocytes and increased level of CD4+CD25+ T lymphocytes in relation to tumor progression have been described. 17
It is known that the application of NDV suppresses tumor growth or induces even its elimination if combined with local irradiation. 18 However, the question remains as to what is the possible action of NDV on a particular T-cell incidence and how this is related to the host immune reaction against the tumor. In the present study, the tumor growth rate and the incidence of particular groups of T lymphocytes are determined in C3Hf/HZgr mice injected with SCCVII mammary carcinoma treated with NDV at particular times after tumor transplantation to shed new light on these phenomena.
Materials and Methods
Experimental animals
Male C3Hf/HZgr mice were obtained from the Animal Breeding Unit of Ruđer Bošković Institute (RBI, Zagreb, Croatia). At the beginning of the experiment, the animals were 3 months old with body weight 20–22 g. Admission to food (4 RF 21 GPL; Mucedola srl) and tap water was ad libitum. The temperature of the room with experimental animals was 22°C, humidity 55%, and light–dark intervals exchanged every 12 hours. All experiments were performed in accordance with the Croatian law on animal welfare.
Tumor and virus application
Spontaneously arising murine mammary squamous cell carcinoma (SCCVII), obtained in 1996 from the British Columbia Cancer Research Centre, Vancouver, Canada, was used in the experiments. Tumor samples are kept in the RBI tumor bank. These preliminary in vitro experiments indicated that the NDV LaSota strain at a dose of 200 EID50/cell inhibited almost completely the growth of cancer cells after 48 hours of exposure.
Tumor cell suspension was mechanically prepared from growing tumor as described previously 19 with 107 viable cells in 1 mL. The mice were injected subcutaneously with 0.1 mL of the suspension (106 viable cells). Eight, 13, and 18 days following tumor transplantation, the mice were injected with 5×108 EID50 of NDV either intraperitoneally or directly into the growing tumor. The dose of NDV used in these experiments was determined as the dose that, in the preliminary studies, showed a significant antitumor activity and, based on available data from the literature, 2,20 could show a certain immunomodulatory potential in tumor-bearing organisms. In both treatments, the required dose of the virus was administered in 100 μL of saline. There were seven animals in each group. By using the caliper, three orthogonal diameters (A, B, and C) of growing tumor were determined every 2 or 3 days. Tumor volume was determined by using the formula ABCπ/6. The experiment was repeated twice.
Virus
An apathogenic strain of NDV, LaSota strain (hereinafter LS), was used in the experiments. The virus was cultivated in an allantoic fluid of fertilized 10-day-old SPF (Specific Pathogen Free) chicken eggs. The allantoic fluid was collected and EID50 was determined. Virus titer was adjusted on 109 EID50/mL allantoic fluid, and subsequently, the allantoic fluid was lyophilized (Pestikal LaSota spf; Veterina d.o.o.).
Spleen lymphocyte suspension
Ten and 20 days after tumor transplantation, the mice were sacrificed, spleens removed, single-cell suspensions prepared mechanically, and lymphocytes were isolated on a density gradient by using Lymphoprep. 21 At each time point, seven mice per group were used.
Determination of lymphocyte populations
Spleen lymphocytes were washed once in phosphate-buffered saline (PBS) pH 7.3 with 5% fetal calf serum (FCS) adjusted to a final concentration of 1×106 cell per mL and incubated with particular monoclonal antibodies for 30 minutes on ice in the dark. Monoclonal antibodies anti-CD3, anti-CD4, anti-CD8, and anti-CD25, as phycoerythrin or FITC conjugate (BD Pharmingen Bioscience), and corresponding isotype controls were used. The cells were washed in PBS, centrifuged at 200×g for 5 minutes, and resuspended in PBS with 5% FCS. All samples were finally resuspended in a propidium iodide (1 μg/mL)-containing buffer.
Flow cytometric analysis
The fluorescence intensity at the single-cell level was evaluated by flow cytometry (FACScalibur; Becton Dickinson). For each sample, 10,000 cells were analyzed. Fluorescence signals were recorded on a frequency histogram or dot plot using logarithmic amplification. Cells were considered unstained if the associated fluorescence did not differ from the fluorescence of cells labeled with isotype control (negative control). Nonviable cells labeled with propidium iodide were excluded from analysis. The experiment was repeated twice.
Statistical analysis
Mean values and standard deviations were calculated for each experimental group. The independent sample t-test and one-way analysis of variance (ANOVA) followed by the Tukey post hoc test were used to determine significant differences between groups. Statistical significance was defined as p<0.05.
Results
The influence of NDV on the growth of SCCVII mammary carcinoma transplanted into C3Hf/HZgr mice
The eighth day following the injection of 106 carcinoma SCCVII cells subcutaneously into the right thigh of C3Hf/HZgr mice the tumor volume reached about 80 mm3. Eight, 13, and 18 days following tumor transplantation, the mice were further injected with 5×108 EID50 of NDV either intraperitoneally (NDV i.p.) or directly into the growing tumor (NDV i.t.). As presented in Figure 1, 2 days following the first virus injection, that is, on day 10 after tumor cell transplantation, there was no difference in tumor volume between the groups (one-way ANOVA, p>0.05). However, in the period of 13–21 days after injection of tumor cells, both NDV treatments caused a statistically significant inhibition (p<0.05) of tumor growth, ∼35%–45% compared to control. Starting from the 24th day, a substantial increase in tumor volume was observed in the NDV i.t. group compared to the NDV i.p. group, but the difference between these two virus treatments was not statistically significant. On day 24, a significant difference (p<0.05) was observed only in the NDV i.p. treatment compared to control.
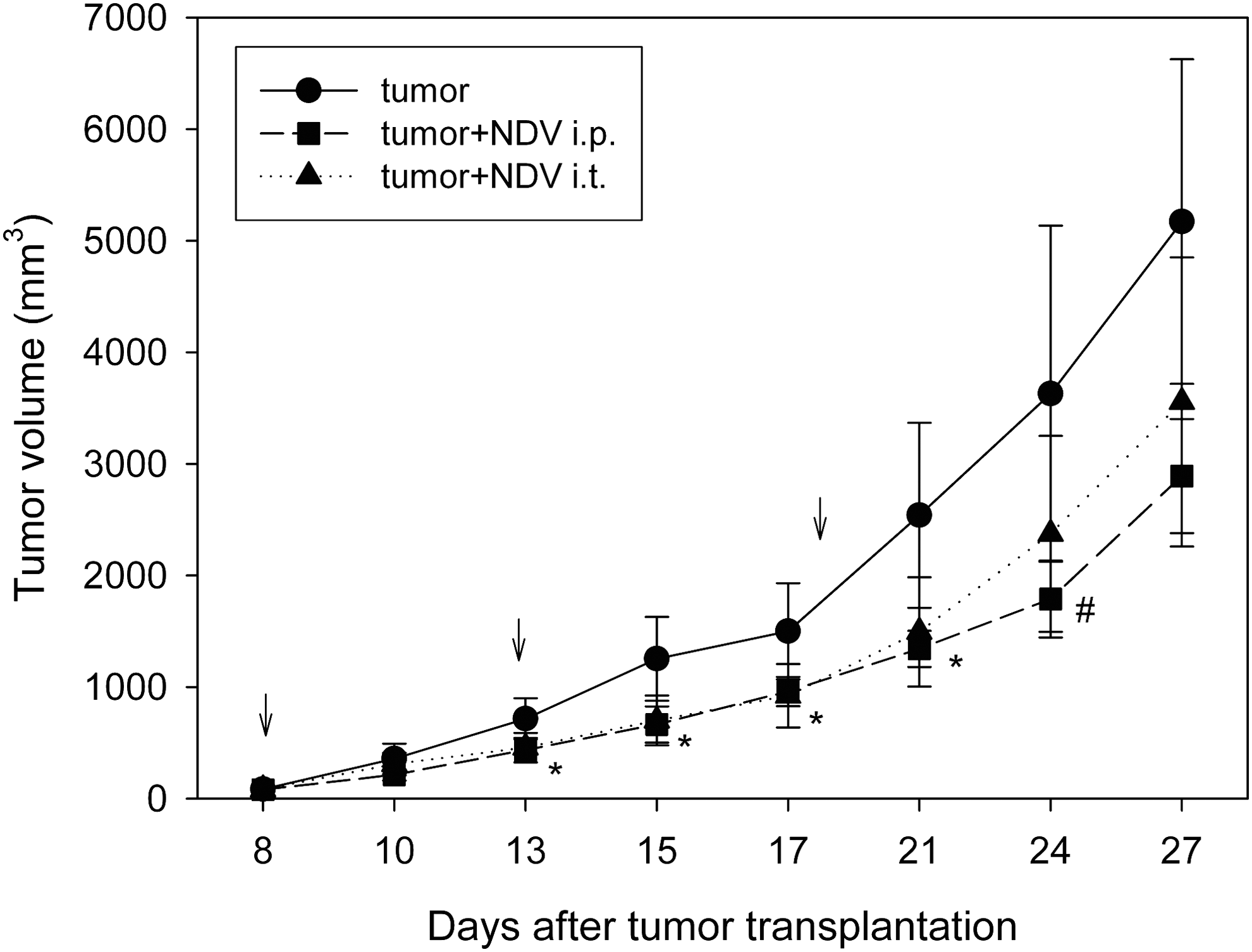
Tumor growth curves after SCCVII tumor cell subcutaneous injection to C3Hf/HZgr mice treated with 5×108 EID50 of Newcastle disease virus (NDV) LaSota virus either intraperitoneally or directly into the tumor 8, 13, and 20 days (arrows) after tumor transplantation. Mean values of tumor volume with SD (N=7) are shown. Asterisks indicate statistically significant differences (p<0.05, Tukey post hoc test) compared to control for both NDV-treated groups on days 13–21, and the sign (#) indicates significant difference (p<0.05) between NDV i.p. and control group on day 24 after tumor transplantation.
The frequencies of T-lymphocyte populations into the spleen of C3Hf/HZgr mice with SCCVII carcinoma following NDV treatment
By using specific monoclonal antibodies (anti-CD3, anti-CD4, anti-CD8, and anti-CD25—as phycoerythrin or FITC conjugates with particular isotype controls), the frequencies of CD4+, CD8+, and CD4+CD25+ T lymphocytes were determined in the spleen of C3Hf/HZgr mice with growing SCCVII carcinoma 10 and 20 days after tumor cell injection. Furthermore, mice bearing the tumor were injected intraperitoneally with NDV (LS) 10, 13, and 18 days after tumor cell transplantation and the frequencies of particular T-cell populations determined 10 and 20 days after tumor transplantation.
As presented on Figure 2, there is a pronounced decrease in the frequencies of CD4+ and CD8+ T lymphocytes with an increase of CD4+CD25+ T-lymphocyte frequency during tumor growth. However, if NDV is applied, the frequencies of CD4+ and CD8+ T lymphocytes are about normal 10 and 20 days after tumor transplantation. Furthermore, a pronounced decrease of CD4+CD25+ T lymphocyte incidence 20 days after tumor transplantation and 2 days after the third NDV injection is of particular interest pointing to the immunomodulating action of the virus in the tumor-bearing organism.
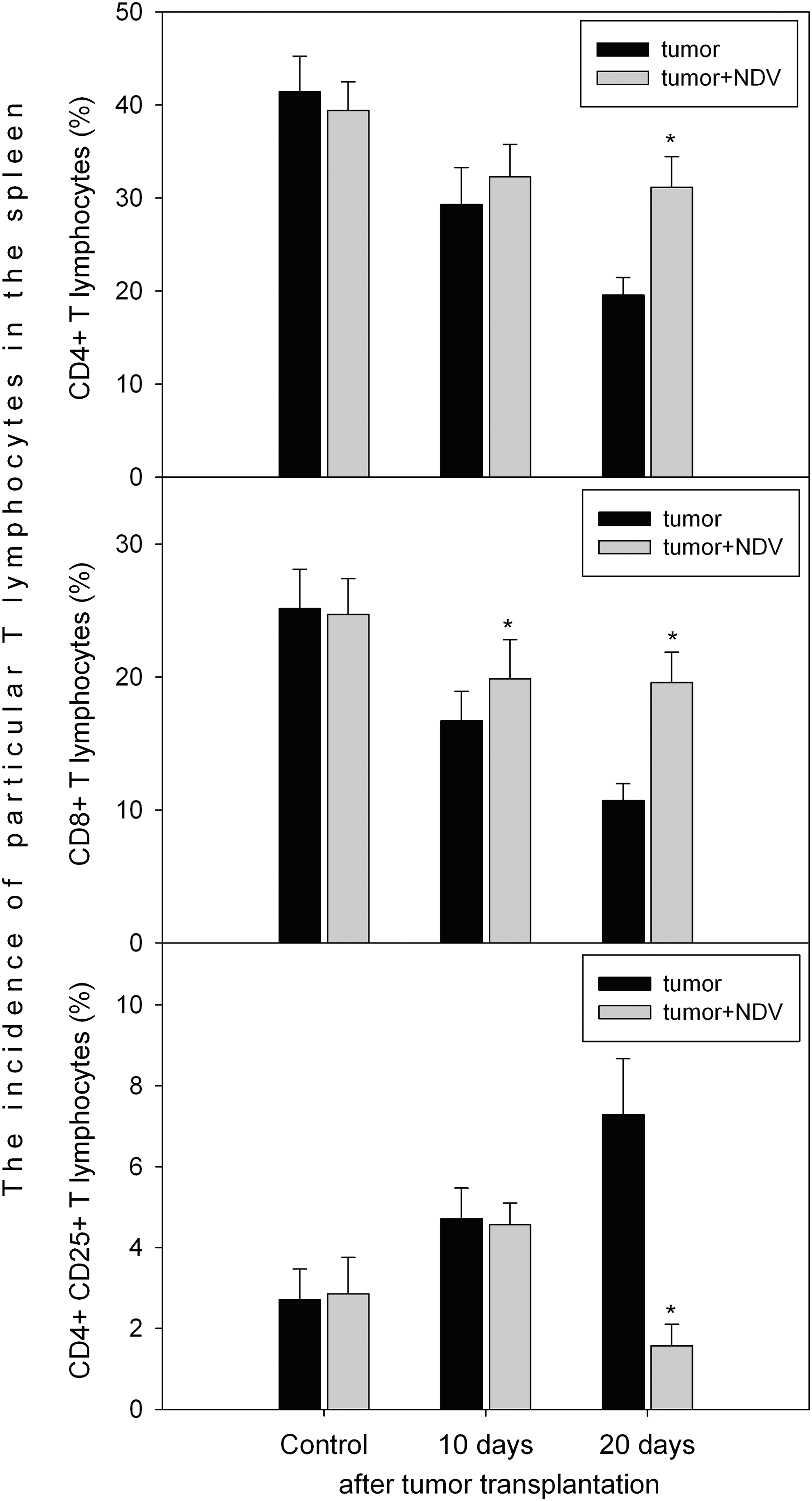
The incidence of CD4+, CD8+, and CD4+CD25+ T lymphocytes in the spleen of C3Hf/HZgr mice transplanted subcutaneously with SCCVII cells and treated intraperitoneally with 5×108 EID50 of NDV LaSota virus 8, 13, and 18 days after tumor transplantation. The analyses were performed 10 and 20 days after tumor cell transplantation. Asterisk above the bar indicates statistically significant difference to nontreated tumor-bearing mice (p<0.01, t-test for independent sample, N=7, two independent replicates per spleen sample were performed).
Discussion
The application of NDV to tumor-bearing mice either directly into the growing tumor or intraperitoneally resulted in a significant suppression of transplanted tumor growth, which is in accordance with previous and numerous other results. 8,17,18,22,23 Furthermore, the results presented here pointed to the important role of NDV on the incidence of particular T-lymphocyte populations in tumor-bearing mice.
It is well known that in mice with a growing tumor, the incidence of CD4+ and CD8+ T cells decreased and CD4+CD25+ increased. 10,15,24
It was shown that a tumor of different origins increase Treg-cell incidence over the homeostatic conditions, in tumor-draining lymph nodes and spleen. 24 Several data point to the tumor-creating environment similar to chronic inflammation and a source of both chemokines and TGF-β influencing migration and conversion of CD4+ T cells, respectively. 24 –27 TGF-β induces a proliferation of Treg cells, and tumors not secreting TGF-β can directly license immature myeloid dendritic cells to produce it through unknown mechanisms. 24,28 TGF-β is one of the most potent tumor-produced suppressive factor enabling conversion of naive T cells into suppressor cells while inhibiting proliferation of preexisting Tregs. 24,29 It was described that Tregs inhibit the immune response predominantly at the tumor site, whereas few remain inside the draining node, 30,31 but there are observations about increased Treg numbers in draining the lymph node and spleen of tumor-bearing mice. 14,24,32
NDV application in mammary carcinoma-bearing mice, as presented in this article, resulted in an increased incidence of CD4+ and CD8+ T lymphocytes in the spleen being significantly higher in comparison to nontreated tumor-bearing mice. This is in accordance with clonal expansion of CD4+ and CD8+ lymphocytes in mice with growing tumor if NDV was applied. 32,33 It is, further, important to note that the T-cell-dependent immune response is recovered in the patients with tumor treated with NDV. 34,35
Treg mediate their suppressive function through inhibitory mechanisms that depend on direct cell contact and soluble molecules, including inhibitory cytokines, cytolysis, metabolic disruption, and targeting of dendritic cells. 36 Thereby, preferentially inhibitory cytokines, including IL-10, TGF-β, and IL-35, cytolysis through secretion of granzymes, perforin, galections, and induction of death-receptor pathway-mediated apoptosis in effector T cells are used. 36,37
Furthermore, in addition to Treg analysis, myeloid-derived suppressor cells as well as changes in their key cytokines such as TGF-β, IL-10, arginase 1, and nitric oxide synthase should be analyzed to confirm these findings.
It is of particular interest to note that NDV application in mammary carcinoma-bearing mice resulted in a decreased incidence of CD4+CD25+ T lymphocytes if three doses of NDV were used. Namely, as presented, only one NDV injection did not influence CD4+CD25+ cell incidence. This is the first observation about NDV application on T reg cell incidence in tumorous mice.
In addition, the results can be discussed in the context of the NDV doses used in this study as well as their potential use in humans. It was shown that substantially higher doses than those used in the present study can be safely administered in humans. 2 For example, the maximal tolerated dose was around at least 3.3×109 infectious particles when the virus was administered by the intravenous route, and even higher doses of NDV (4.3×1012 infectious particles) can be safely administered in intratumoral route. 38 Therefore, taking into account these data, the dose of NDV applied in this study, and for which the authors have shown to have immunomodulatory effects in tumor-bearing organisms, can be considered as easily achieved and highly tolerable in humans.
To conclude, NDV is involved in the modification of tumor-bearing organisms' immunological response by keeping the frequencies of CD4+ and CD8+ close to normal values and by reducing CD4+CD25+ frequency. These events, in particular T-lymphocyte incidence and well-known direct NDV action against tumor cells are responsible for the successful therapeutical antitumor approach particularly if combined with proper tumor irradiation. 18
Footnotes
Acknowledgment
This study was supported by the Ministry of Science, Education and Sports of the Republic of Croatia (Grant No. 009899 and 1-08-198).
Disclosure Statement
The authors declare no potential conflicts of interest.