
Abstract
Dickkopf-related protein 3 (DKK3) gene, as a tumor suppressor gene, has been discovered in various cancers, but its relationship with tumor chemoresistance is still unclear. In this study, this laboratory detected that DNA methylation contributes to the downregulation of DKK3 in docetaxel resistance of human lung cancer cells and its possible biochemical mechanism. DKK3 has been proved to be downregulated by hypermethylation in docetaxel-resistant lung cancer cells. Upregulation of DKK3 can reverse the chemoresistance of docetaxel-resistant cell lines in vitro by growth inhibition and enhancement of apoptosis. Conversely, downregulation of DKK3 could induce parental human lung cancer cells insensitivity to docetaxel by promoting proliferative capacity and inhibiting apoptosis of cancer cells. In addition, the authors observed that overexpression of DKK3 might decrease the expression of P-glycoprotein. All results suggested that epigenetic downregulation of DKK3 leads to docetaxel resistance in human nonsmall cell lung cancer (NSCLC) cells by increased expression of P-glycoprotein. DKK3 may reveal a novel molecular target for docetaxel resistance for NSCLC patients in the future.
Introduction
Lung cancer is one of the highest incidence of malignancies and leading cause of cancer-related death in the world. It is reported that 80% of all lung cancer patients are nonsmall cell lung cancer (NSCLC). Most of the lung cancer patients are diagnosed in the advanced stages (Stages III and IV). 1 Despite development of cancer treatment and introduction of new technology, advanced lung cancer remains associated with poor prognosis. Docetaxel, a semisynthetic analogue of paclitaxel, is widely used as an anticancer drug in the treatment of lung cancer. 2 Docetaxel confers its antineoplastic activity by inhibiting microtubule depolymerization, leading the metaphase to anaphase transition arrest, activating the spindle assembly checkpoint, and subsequently causing apoptosis. 3,4
However, chemotherapy agents become increasingly ineffective when the disease has spread beyond the lung at the advanced stage. Chemoresistance almost occurs in recurrent patients and remains the most important dilemma in restricting the clinical application of anticancer drugs. 5 Thus, a further study of the molecular mechanism underlying docetaxel resistance of NSCLC should be helpful to improve the efficacy of its clinical application.
Accumulated evidences have proved that cancer cells resistant to docetaxel result from both genetic and epigenetic modification of various critical genes. 6,7 Aberrant DNA methylation, leading to downregulation of key genes, which played a crucial role during the initiation and development of cancer. 8 However, the correlation between DNA methylation and docetaxel resistance of NSCLC is rarely reported.
In the laboratory, the authors previously established a docetaxel-resistant H1299 cell line (H1299/DTX) by continuous exposure to increasing concentrations of docetaxel. It is important for us to understand the molecular mechanisms underlying docetaxel resistance and explore novel therapeutic markers for reversing chemoresistance in NSCLC.
Dickkopf-related protein 3 (DKK3), a putative Wnt antagonist, has been proved to represent as a tumor suppressor for downregulation in various human cancers. In many types of cancers, the failure of its normal expression is closely associated to CpG island methylation on the DKK3 promoter; thus, the published articles of DKK3 have mainly focused on the methylation-related field. 9 The silencing expression of DKK3 is always correlated with poor prognosis of human cancer, including lung cancer, 10 ovarian cancer, 11 renal clear cell carcinoma, 12 and prostate cancer. 13 However, the association of DKK3 with chemotherapy resistance in NSCLC cells was seldom seen in the previous studies. It is vitally necessary that the molecular mechanism underlying docetaxel resistance should be elucidated and searched for novel therapeutic markers for reversing chemoresistance.
In this study, the authors investigated that epigenetic downregulation of DKK3 mediated by aberrant promoter methylation contributes to docetaxel resistance in NSCLC cell lines. At the same time, DKK3 re-expression could reverse docetaxel resistance of lung cancer cell. The results of this study may provide a novel strategy for reversing chemoresistance of NSCLC.
Materials and Methods
Cell culture
Human NSCLC H1299 cells were bought from the Cell Bank of Chinese Academy of Medical Science. Cells were conditioned in the RPMI 1640 medium containing 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/L streptomycin in a humidified atmosphere at 37°C, 5% CO2. The docetaxel-resistant H1299 cell line (H1299/DTX) was established by continuous exposure to increasing concentrations of docetaxel, and cells were grown in the presence of 5 μg/L final concentration of docetaxel.
RNA isolation and semiquantitative reverse transcription–polymerase chain reaction
Total RNA of tumor cells was isolated using the Trizol reagent (Invitrogen), and the spectrophotometer (BioPhotometer) was used to measure the concentration of total RNA. Reverse transcription (RT) was accomplished by the PrimeScript™ RT Reagent Kit (Takara) following the instruction manual of the manufacturer. Two micrograms of total RNA was applied for cDNA synthesis, and 20 μL reaction volume was prepared for further polymerase chain reaction (PCR). PCR conditions were in accordance with the manufacturer's instructions. The primer sequences for each gene were designed as shown in Table 1. The transcription of glyceraldehyde 3-phophate dehydrogenase (GAPDH) was performed for normalization. The PCR products were electrophoresed on 2% agarose gels and detected by ethidium bromide (EB) staining.
PCR, polymerase chain reaction; DKK3, Dickkopf-related protein 3; MDR-1, multidrug resistance-1; GAPDH, glyceraldehyde 3-phophate dehydrogenase.
Methylation-specific PCR
Methylation-specific PCR (MSP) was performed to detect the methylation status of DKK3 gene promoter by using bisulfate-modified genomic DNA as the template. Total DNA was isolated from cultured cells using the QIAamp DNA Mini Kit (Qiagen). One microgram of genomic DNA was bisulfate treated with the EZ-DNA methylation Gold Kit (Zymo Research) and finally resuspended in 10 μL TE buffer. Two sets of primers were used to display the differences between methylated and unmethylated status. The primers targeting the unmethylated and methylated DKK3 promoter regions are described in Table 1. The following steps were performed with an instruction manual. The PCR products were visualized in 2% agarose gel by EB staining. Lymphocyte DNA was used as unmethylation and methylation-positive control in the experiment. A water blank was performed as a negative control.
Transient transfection plasmid or siRNA interference
The DKK3 expressing plasmid (pCS2-hDKK3) and negative control were available from Addgene (Plasmid 15496). Small interfering RNAs (siRNAs) targeting DKK3 (siRNA-DKK3) were obtained from Santa Cruz Biotechnology, Inc. Nonspecific control siRNA (siRNA/NC) was used as control. Plasmid DNA was extracted by the Plasmid Mini Kit (Qiagen). Cells were cultured in six-well plates (2×105cells/well) and transfected with the SiRNA Mate (GenePharma) or Turbofect Transfection reagent (Thermo Scientific) following the manufacturer's protocol.
Western blot
The cells were isolated directly or 72 hours after transfection; the cells were washed twice with phosphate buffered saline (PBS) before the proteins were extracted. Proteins were separated electrophoretically by 10% sodium dodecyl sulfate–polyacrylamide gel and transferred to a polyvinylidene fluoride (PVDF) membrane. The membranes were blocked for 2 hours with 5% nonfat milk at room temperature. The membranes were incubated first with the REIC/DKK3 antibody (1:2000; Abcam) or P-glycoprotein antibody (1:500; Bioss) overnight at 4°C and then incubated with a second antibody for 2 hours at 37°C. Proteins were then displayed with the ECL substrate (Cell Signaling Technology) in accordance with the manufacturer's instructions. Protein levels were normalized to GAPDH.
Cell viability assay
Single-cell suspensions were planted into 96-well plates (2×103 cells/well) directly or 24 hours after transfection. Various concentrations of docetaxel (0, 10, 20, 40, 80, 160, 320, 640, 1280 μg/L) were then added. Forty-eight hours later, cell viability was shown by the MTT assay.
Colony formation assay
Three hundred cells were planted into each well in six-well plates directly or 48 hours after transfection. After 14 days, cells were fixed with methanol and stained with 0.1% crystal violet. Then, the number of visible colonies was counted. The cloning efficiency was calculated as follows: (the clone number/the number of plant cells) ×100%.
Flow cytometric analysis of apoptosis
Cells were harvested 48 hours after transfection for cell apoptosis measurement, then resuspended in 0.5 mL binding buffer with 5 mL Annexin V and 5 mL propidium iodide (PI), and reacted for 15 minutes at 37°C in the dark. Then, the early apoptotic rate was measured by flow cytometric analysis following the manufacturer's protocol.
Statistical analysis
The results are presented as mean±standard error and they were analyzed by SPSS16.0 software (SPSS). The differences between two groups were carried out by Student's t-test. A p value of <0.05 was considered statistically significant.
Results
DNA methylation induces silent expression of DKK3 in H1299/DTX cell compared with parental cell lines
The H1299/DTX cell line has been established by exposure to increasing concentrations of docetaxel in the laboratory. The IC50 value of H1299/DTX cells against docetaxel was increased significantly compared with the parental H1299 cell line (357.63±31.89 vs. 28.62±10.07 μg/L, p<0.01; Fig. 1A-c). DKK3 was shown to be downregulated in H1299/DTX compared to H1299 cells. The results were displayed by semiquantitative PCR and western blot in the two cell lines (Fig. 1A-a, b). To detect whether DNA methylation induced the downregulation of DKK3, MSP analysis was used to investigate the methylation status of the promoter in the DKK3 gene. The results showed less densely methylated H1299 cells compared with H1299/DTX (Fig. 1B). Furthermore, cells treated with 5-Aza-dC, a specific DNA methyltransferase inhibitor, for 72 hours led to a significantly increased expression of DKK3 mRNA and protein in the H1299/DTX cell line in a dose-dependent manner (Fig. 1C-a, b). All the results illustrated that expression of DKK3 might be associated with DNA methylation in the cells. Moreover, the expression levels of mRNA and protein of DKK3 were decreased in a dose-dependent status after exposure to docetaxel in the H1299 cell line (Fig. 1D-a, b), suggesting that the expression of DKK3 might be associated with docetaxel chemosensitivity in NSCLC cells.
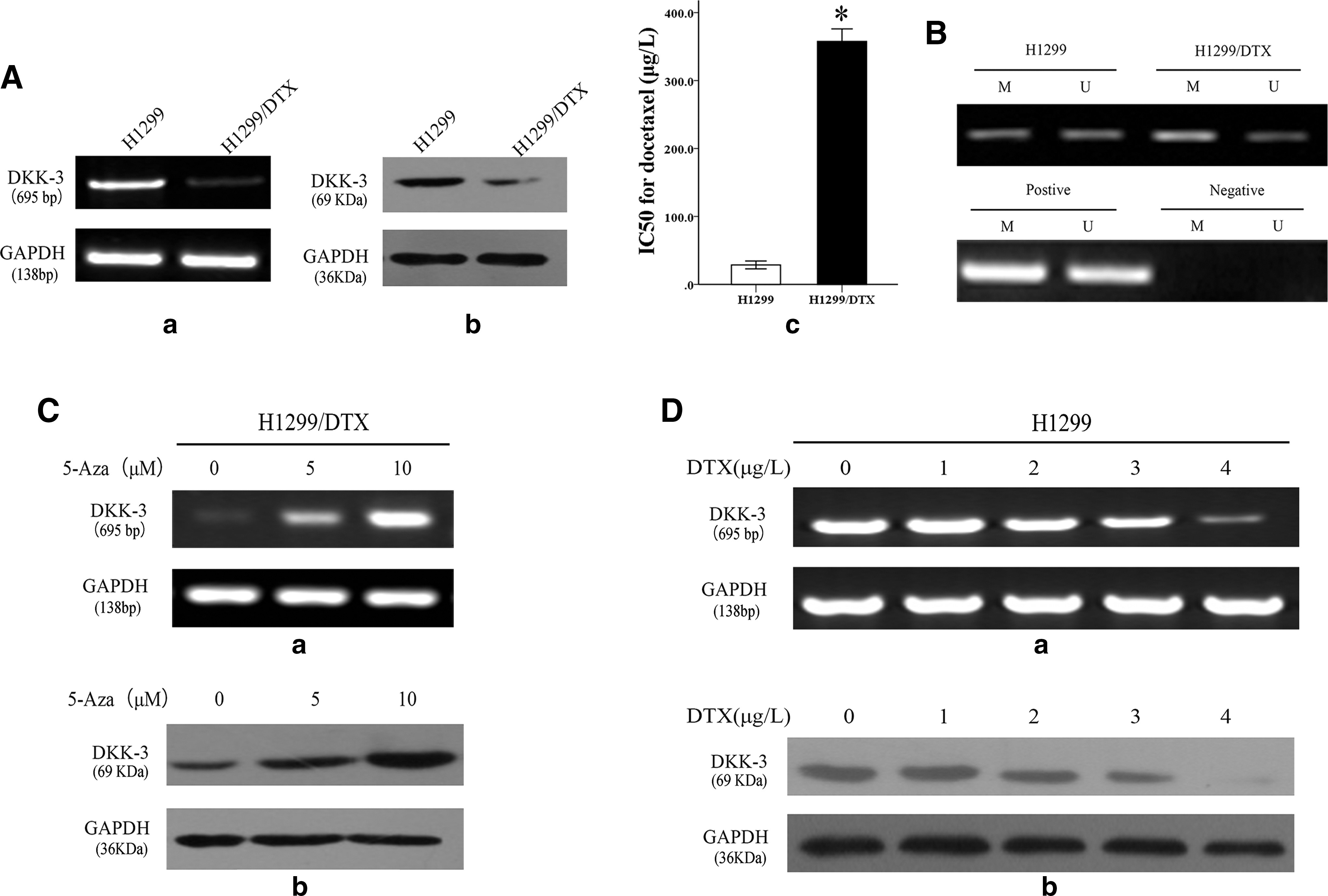
Dickkopf-related protein 3 (DKK3) was downregulated by DNA methylation in docetaxel-resistant lung cancer H1299/DTX cell line compared with the parental H1299 cell line.
DKK3 affects cell apoptosis, proliferation, and chemosensitivity to docetaxel in NSCLC cells
H1299/DTX cells and parental cell lines were transfected with pCS2-hDKK3 or SiRNA-DKK3, respectively, for the following experiments. The DKK3 gene and protein were evidently increased in the H1299/DTX cell lines and decreased in parental cells by PCR and western blot (Figs. 2A-a, b and 3A-a, b).
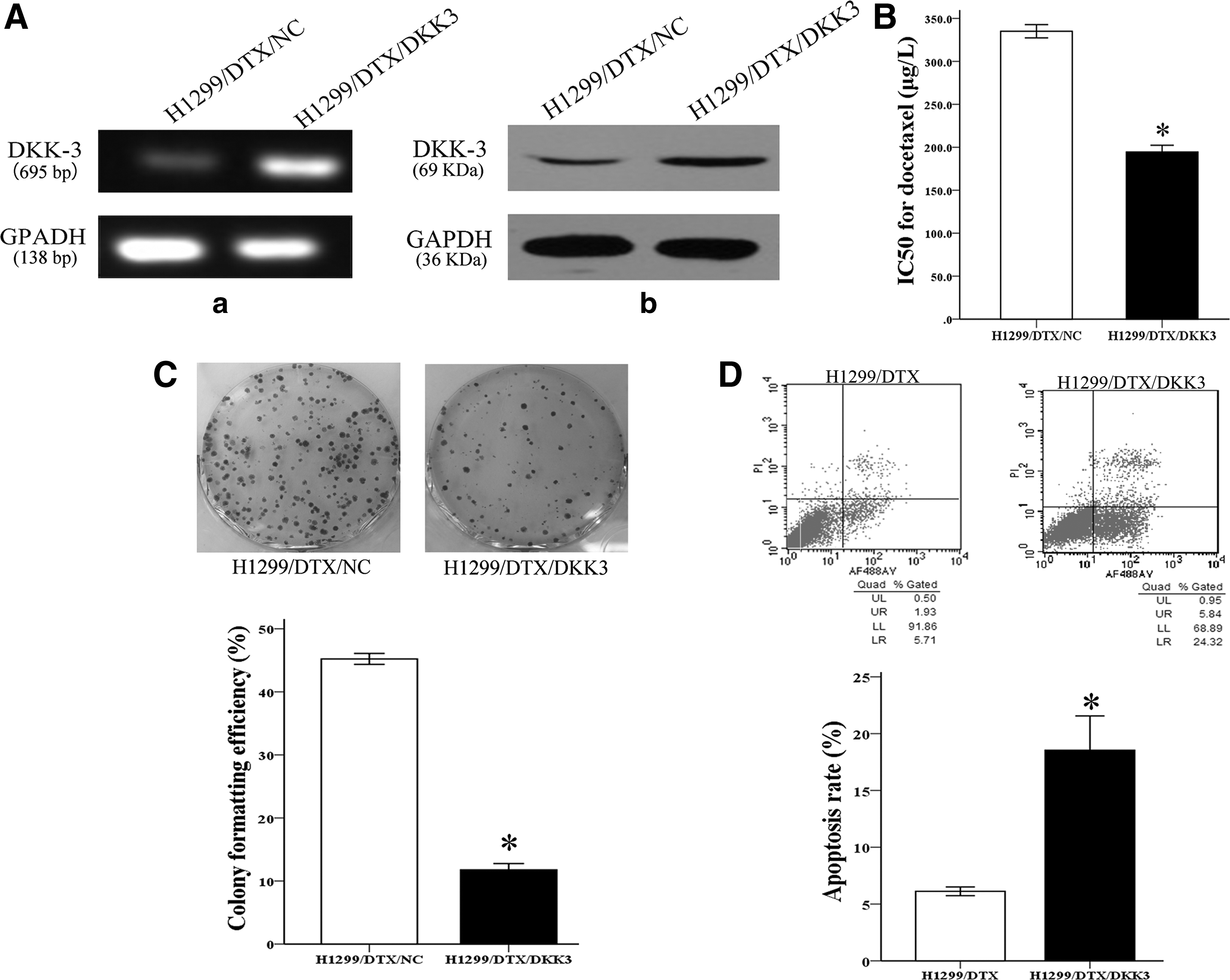
Overexpression of DKK3 reversed chemoresistance of the H1299/DTX cell line to docetaxel in vitro.
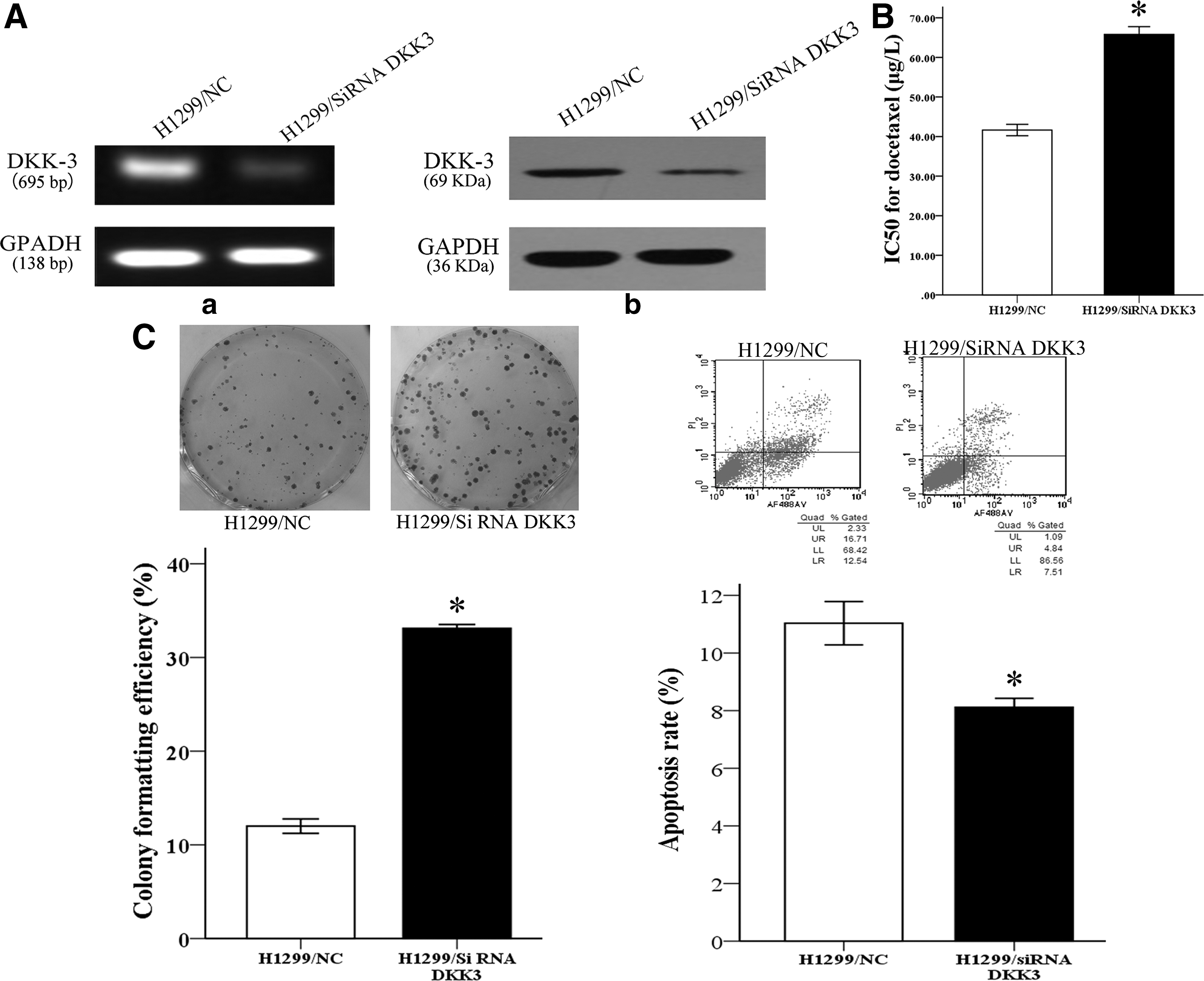
Knockdown of DKK3 expression decreased chemosensitivity of the H1299/DTX cell line to docetaxel in vitro.
The chemosensitivity of NSCLC cells to docetaxel was performed by the MTT assay. The IC50 value for docetaxel in siRNA/DKK3-transfected H1299 cells was increased by 1.58-fold (65.79±3.40 vs. 41.64±2.48 μg/L) compared with siRNA/NC-transfected parental cell lines (p<0.05; Fig. 3B). Conversely, the IC50 value of docetaxel in DKK3-transfected H1299/DTX cells was significantly reduced by 58.04% (194.47±13.65 vs. 335.09±13.34 μg/L) compared with NC-transfected resistant cells (p<0.05; Fig. 2B). These results indicated that the expression level of DKK3 had a close relationship with the chemosensitivity of NSCLC cells to docetaxel.
Furthermore, the authors observed the proliferating ability of cell-transfected siRNA or plasmid by colony formation assay. Overexpression of DKK3 significantly inhibited the colony formation capacity in docetaxel-resistant cells (p<0.05; Fig. 2C). Conversely, siRNA-mediated DKK3 knockdown promoted colony formation of parental cells (p<0.05; Fig. 3C). It indicated the proliferation inhibitory ability of DKK3 in vitro. They also detected the apoptosis rate by flow cytometric analysis. It was shown that enforced expression of DKK3 led to an increase of the apoptosis rate in H1299/DTX from 18.53%±5.23% to 6.12%±0.66% compared with the NC group (p<0.05; Fig. 2D). In addition, downregulation of DKK3 in H1299 cells results in a decreased apoptosis rate (11.03±1.30 vs. 8.11±0.53, p<0.05; Fig. 3D). Taken together, these results demonstrated that upregulation of DKK3 increases the sensitivity of NSCLC cells in vitro to docetaxel by inhibiting cell growth and enhancing cell apoptosis. Similar results have been shown in A549 cells (Supplementary Fig. S1).
DKK3 resensitizes H1299/DTX cells to docetaxel by downregulated expression of P-glycoprotein
P-glycoprotein (P-gp), an efflux pump, plays an important role for multidrug resistance in tumor cells, which is encoded by the multidrug resistance-1 gene (MDR-1 gene). Combined with the previous results, the authors speculated that P-gp may be an important protein associated with docetaxel resistance in the H1299/DTX cell line (Fig. 4A). To better understand the association of DKK3 expression and docetaxel chemosensitivity, they upregulated the expression of the DKK3 gene in H1299/DTX cell lines. Results from PCR and western blot revealed a significantly decreased expression level of the MDR-1 gene and P-gp in DKK3-transfected docetaxel-resistant H1299 cells, implying that resensitization of NSCLC cells to docetaxel by DKK3 may occur through MDR-1 downregulation (Fig. 4B-a). P-gp, associated with chemoresistance, was also downregulated in DKK3-transfected docetaxel-resistant NSCLC cells (Fig. 4B-b).
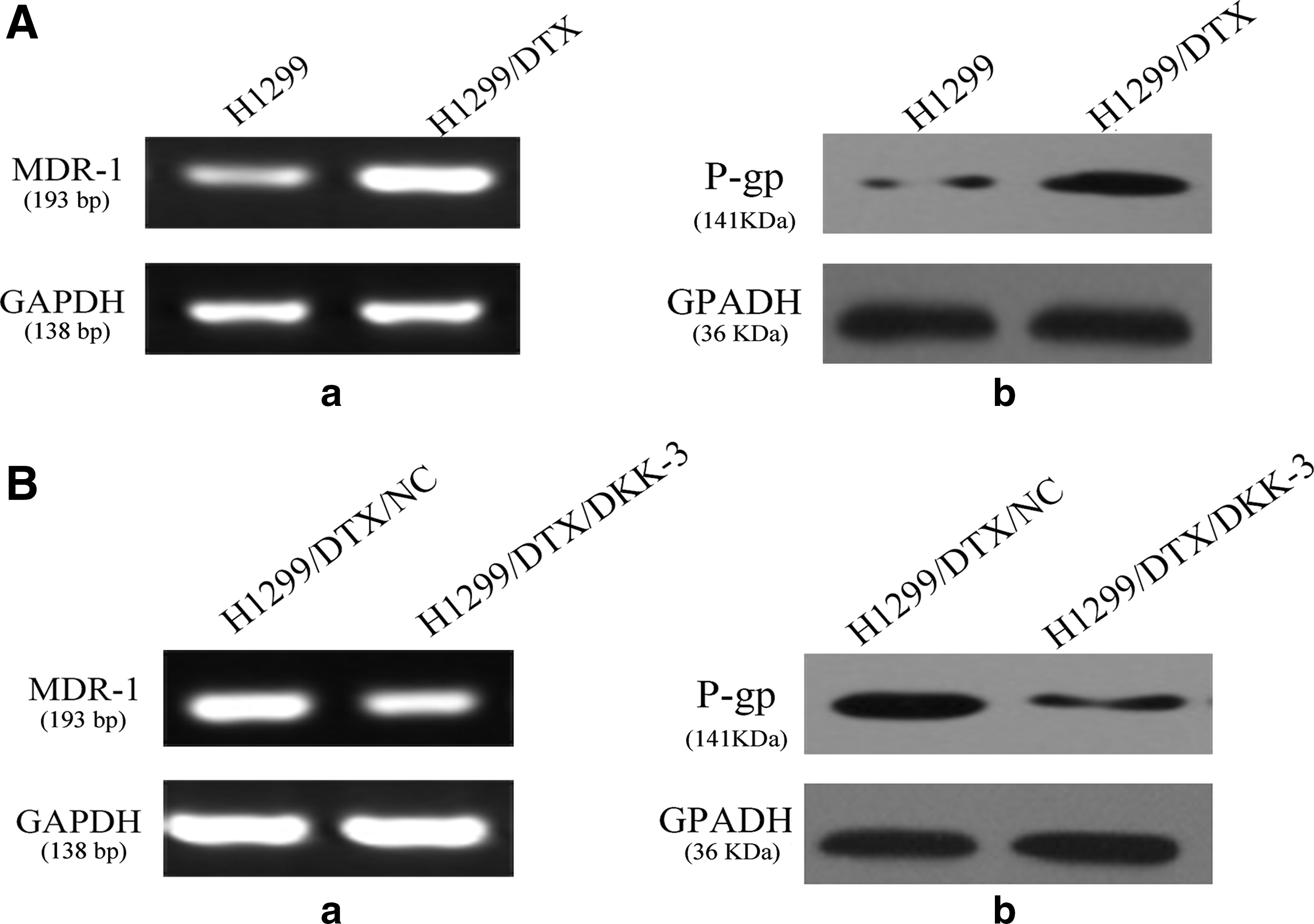
Expression of P-glycoprotein was associated with regulation of DKK3.
Discussion
Docetaxel is always used as a chemotherapy agent in the treatment of NSCLC. Chemoresistance has been the most important obstacle of docetaxel in clinical application for patients. 14 To gain a better insight of the molecular mechanism of chemoresistance of NSCLC and searching for the reversion treatment, the docetaxel-resistant NSCLC cell line H1299 has been established in the laboratory after selection by drug pressure and named as H1299/DTX. The docetaxel-resistant cells displayed morphological and physiological differences compared with parental NSCLC cells. 15 Previous reports have shown that multiple factors were associated with the sensitivity of tumor cells to docetaxel, including alteration in tubulin dynamics, 16 differences in the expression of β-tubulin isotype, 17 and overexpression of the MDR gene. 18 Both genetic and epigenetic dysregulation of crucial genes were involved in the resistant mechanism of docetaxel in NSCLC cells.
DKK3, one of Wnt antagonists in the DKK family, is usually inactivated in several human malignancies with hypermethylated CpG islands in the promoter regions and acts as a suppressor gene in cancer cells. 19 Epigentic silence, especially aberrant DNA hypermethylation, is an important mechanism for tumor suppressor gene inactivation in cancer, which has been proved in many reports. 20 Aberrant expression of DKK3 is a critical determinant factor of cancer outcome. Functional inactivation of DKK3 is closely correlated with gene mutation, epigenetic inactivation, or cytoplasmic mislocalization in many cancers, in which hypermethylation of CpG islands in the promoter regions is often observed. 21 Moreover, re-expression of DKK3 significantly inhibited the growth of tumor cells and xenografts in mice, as well as induced cell apoptosis. 22 Although these results demonstrated that DKK3 may play a critical role in tumor progression, to date there are almost no reports focused on the relationship of DKK3 expression with chemosensitivity to tumor cells. However, the role of epigenetic silence of DKK3 in human docetaxel-resistant lung cancer cells remains unclear and needs to be further elucidated.
In the study, DKK3 was downregulated significantly in H1299/DTX cells compared with H1299 cells. MSP analysis demonstrated that the methylation status of DKK3 in H1299/DTX cells was higher than parental cell lines. 5-Aza-dC, a specific DNMT inhibitor, restored DKK3 expression in a dose-dependent manner. Moreover, DNA and protein levels of DKK3 were decreased in a dose-dependent status response to docetaxel in H1299 cells. The results indicated that DKK3 expression may be associated with chemosensitivity to docetaxel.
Next, the authors detected that overexpression of the DKK3 gene in docetaxel-resistant cells led to an increased response to docetaxel in vitro and was associated with the inhibition of cell proliferation and enhancement of apoptosis. Conversely, downregulation of DKK3 expression in parental H1299 cells induced relative insensitivity to docetaxel by enhancing the ability of cell proliferation and decreasing cell apoptosis. This study showed that the levels of DKK3 expression might contribute to the sensitivity to docetaxel in human lung cancer cells.
Furthermore, the authors discussed the possible molecular mechanisms of DKK3-induced chemosensitivity enhancement of cancer cells. P-glycoprotein, as an efflux transporter encoded by the MDR-1 gene, influences absorption and excretion of a variety of drugs. 23 It plays an important role in the anticancer drug efflux to enhance cell survival against chemotherapy drugs and its overexpression is one of the most crucial mechanisms by which cancer cells are resistant to the cytotoxic effects of anticancer drugs. 24 They hypothesize that inactivation of DKK3 may relieve the expression of P-glycoprotein, thus contributing to docetaxel resistance.
In this experiment, the authors detected that P-gp is upregulated in docetaxel-resistant cell lines. Overexpression of DKK3 may downregulate the expression level of the MDR-1 gene and then affect P-gp expression. The results have partly explained the altered expression of DKK3 in human NSCLC cell lines with different sensitivities to docetaxel. DKK3 acted as a restorer of docetaxel chemosensitivity in NSCLC cells associated with P-gp expression. However, whether DKK3 directly targets MDR-1 or not is still unclear. For a more comprehensive understanding of chemoresistant mechanisms in NSCLC cells, a lot of work is going to be validated in more NSCLC cell lines and tissue samples of cancer patients in the future.
Taken together, this study demonstrates that downregulation of DKK3 was induced by DNA methylation in docetaxel-resistant human lung cancer cells, which contributes to drug resistance by the upregulation of MDR-1 and P-gp expression. Re-expression of DKK3 increases docetaxel chemosensitivity in human NSCLC cells. These results suggest that DKK3 may act as a potential target for reversing docetaxel resistance in NSCLC patients in the future.
Conclusions
In conclusion, the authors have observed that downregulation of DKK3 in docetaxel-resistant human NSCLC cell line H1299/DTX compared with the parental cell line H1299 has proved that DKK3 could act as a chemosensitivity restorer to docetaxel in NSCLC cells by downregulating the P-gp expression and resulting in the inhibition of cell proliferation and enhancement of cell apoptosis. Therefore, the experimental results presented that DKK3 represents a potential therapeutic target for the treatment of docetaxel-based regimen-resistant NSCLC patients.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.