
Abstract
Lung cancer as an aggressive type tumor is rapidly growing and has become the leading cause of cancer-related death worldwide. γ-Glutamylcyclotransferase (GGCT) has been shown as a diagnostic marker in various cancers. To reveal whether there is a correlation between GGCT and lung cancer, GGCT expression in human lung cancer cell lines was first determined by real-time quantitative PCR and western blot. GGCT is expressed in all tested lung cancer cell lines, A549, H1299, and H460. Then, a lentivirus-based system was applied to knock down GGCT in A549 cells, which were thus divided into Lv-shGGCT, Lv-shCon, and Con (noninfected) groups. Methylthiazol tetrazolium assay showed that the cell proliferation was decreased by over 50% in the Lv-shGGCT group compared with controls. The size and number of colonies were dramatically reduced in the GGCT knockdown group, as measured by colony formation assay. Moreover, A549 cells infected with Lv-shGGCT were arrested in the G0/G1 phase as assayed by flow cytometry. Furthermore, the expression levels of CDK4, CDK6, and cyclin D1 were decreased and the cleaved level of PARP was increased in GGCT knockdown cells. In conclusion, GGCT plays a critical role in lung cancer cell proliferation and may be a potential cancer therapeutic target.
Introduction
It has been reported that lung cancer is an aggressive and rapidly growing neoplasm disease. 1,2 The major etiology is primarily genetic and epigenetic damage caused by tobacco. 3 Lung cancer can easily spread to the brain, bone, hepatic, kidneys, and any part of the body. 4 –7 Nearly 85% of lung cancer cases are of nonsmall-cell lung cancer. 8 The traditional treatment for these patients includes surgical, radiotherapeutic, chemotherapeutic, or their combination. 1,9 In some defects, high recurrence rates caused by metastatic spread of tumor cells, low specificity, and side-effects are still present. Despite that notable technical advances have been made in these traditional approaches to the treatment of lung cancer, it remains the leading cause of cancer death worldwide. 1,9 Therefore, there is urgent need to develop a new therapy for the treatment of lung cancer.
In recent years, gene therapy has been an innovative approach to achieve this goal based on genetic alteration, one of the major causes of tumor growth and metastasis. 10 It has been shown that replacement of p53, which played a key role in cell cycle regulation and apoptosis, is a logical approach to therapy for nonsmall-cell lung cancer. 11 The DDR2 kinase gene, which is responsible for 3.8% of lung squamous cell carcinoma (SCC) and cell lines, has mutations that identify a novel therapeutic target for SCC. 12 Target disruption of Skp2 with virally mediated RNA interference has been reported as gene therapy for human small-cell lung carcinoma. 13
γ-Glutamylcyclotransferase (GGCT) is involved in the γ-glutamyl cycle and specifically cleaves γ-glutamyl-amino acid into free amino acids and 5-oxoproline. 14 It contributes to glutathione synthesis and degradation and potentially participates in transport of amino acid across cellular membranes. 15 Recently, overexpression of GGCT has been reported in various cancer tissues, such as esophageal SCC, mammary carcinomas, and osteosarcoma, suggesting that it has significant potential as a diagnostic marker. 15 –17 Knockdown of GGCT by siRNA was observed to inhibit cancer cell proliferation and invasion. 17,18 Nevertheless, the involvement of GGCT in human lung cancer has not yet been demonstrated.
In this study, a series of experiments were carried out to reveal the biological role of GGCT in lung cancer cell growth. The mRNA and protein expression patterns of GGCT were first detected in three lung cancer cell lines, A549, H1299, and H460, by quantitative real-time PCR (qRT-PCR) and western blot. Then, GGCT was knocked down in lung cancer A549 cells by a lentivirus-based system, which is used to functionally silence genes in cultured mammalian cells and stem cells. 19 The cell proliferation, colony formation, and cell cycle progression were analyzed by methylthiazol tetrazolium (MTT), colony formation, and flow cytometry, respectively.
Materials and Methods
Cell lines and culture medium
Human lung cancer cell lines, A549, H1299, H460, and human embryonic kidney cell line, HEK293T, were all purchased from the Cell Bank at the Chinese Academy of Science. A549 cells were cultured in F12K medium (Sigma) supplemented with 10% fetal bovine serum (FBS; Hyclone). H1299 and H460 cells were grown in RPMI-1640 medium (GIBCO) plus 10% FBS (Hyclone). HEK293T cells were maintained in Dulbecco's modified Eagle's medium (DMEM; GIBCO) plus 10% FBS (Hyclone). All cell lines were incubated in a humidified atmosphere (95% air, 5% CO2) at 37°C.
GGCT short hairpin RNA-expressing lentivirus construction
First, the short hairpin RNA (shRNA) lentiviral vector was constructed to induce lentivirus-mediated RNA interference of GGCT expression. The GGCT cDNA sequence was derived from GenBank with accession number NM_024051. The RNAi was designed based on a sequence specific to GGCT (5′- GATTATTTGCATGGGTGCAAACTCGAGTTTGCACCCATGCAAATAATCTTTTTT-3′) and negative control sequence (5′-GCGGAGGGTTTGAAAGAATATCTCGAGATATTCTTTCAAACCCTCCGCTTTTTT-3′). Both shRNAs were ligated into lentiviral pFH-L plasmid (Shanghai Hollybio). To generate GGCT-shRNA and Con-shRNA lentivirus, the plasmid mixture containing either the target sequence or the control was transfected into 80% confluent HEK293T cells with pVSVG-I and pCMVΔR8.92 packaging vectors (Shanghai Hollybio) using Lipofectamine 2000 (Invitrogen). A549 cells were grown in six-well plates at a density of 50,000 cells per well, and then infected with Lv-shGGCT or Lv-shCon lentivirus at a multiplicity of infection of 30. The efficiency of infection was determined by counting fluorescent particle emissions from green florescence protein (GFP) at 72 hours after infection.
Quantitative real-time polymerase chain reaction
Quantitative real-time PCR (qRT-PCR) was used to estimate GGCT expression and knockdown efficiency in lung cancer cells. Cells were harvested and total RNA was extracted using the TRIzol reagent (Invitrogen). cDNA was then reverse transcribed with oligodT using M-MLV reverse transcriptase (Promega) and adjusted to a final concentration of 30 ng/μL. qRT-PCR reactions were performed on the BioRad connect real-time PCR platform using the following system: SYBR premix ex taq 10 μL, forward primers 0.8 μL (2.5 μM), reverse primers 0.8 μL (2.5 μM), cDNA 5 μL, and ddH2O 4.2 μL. All PCR cycles were run according to the following protocol: initial denaturation for 1 minute at 95°C, denaturation for 5 seconds at 95°C, and primer annealing for 20 seconds at 60°C (a total of 40 cycles). The primer sequences for PCR amplification of the GGCT were 5′-CAGCAACCTGCTGACAGAGA-3′ and 5′-CCCTTCTTGCTCATCCAGAG-3′. The primer sequences for actin were 5′-GTGGACATCCGCAAAGAC-3′ and 5′-AAAGGGTGTAACGCAACTA-3′. Fold change of the gene was calculated by the equation 2−ΔΔCt.
Western blot assay
Western blot analysis was carried out to calculate the GGCT protein expression and knockdown efficiency in lung cancer cells. Cells were trypsinized, washed with ice-cold phosphate-buffered saline (PBS), scraped off, centrifuged, and lysed in 2×sodium dodecyl sulfate (SDS) sample buffer (100 mM Tris-HCl [pH 6.8], 10 mM EDTA, 4% SDS, and 10% glycine). Protein (30 μg) was loaded onto a 10% SDS-PAGE gel and transferred to a polyvinylidene fluoride (PVDF) membrane by electroblotting. Membranes were then blocked with 5% (w/v) skim milk and incubated with rabbit anti-GGCT antibody (1:500 dilution, HPA020735; Sigma), mouse anti-CDK4 antibody (1:500 dilution, 2906; Cell Signaling Technology), rabbit anti-CDK6 antibody (1:500 dilution, 19117-1-AP; Proteintech), mouse anti-cyclin D1 antibody (1:1000 dilution, MD-17-3; Medical & Biological Laboratories CO., LTD), rabbit anti-PARP antibody (1:1000 dilution, 9542; Cell Signaling Technology), and rabbit anti-GAPDH antibody (1:60,000 dilution, 10494-1-AP; Proteintech) overnight at 4°C. Then, they were probed with goat antirabbit or goat antimouse HRP-linked secondary antibodies (1:5000, SC-2054 or SC-2005; Santa Cruz) and visualized by the Luminescent Image Analyzer LAS-3000 (Fujifilm).
MTT test
Briefly, all infected and uninfected A549 cells were seeded in 96-well plates at a density of 2500 cells per well. MTT (5 mg/mL) was added after 1, 2, 3, 4, and 5 days of infection, and then incubated at 37°C for 4 hours. The reaction was stopped by addition of 100 μL acidic isopropanol (10% SDS, 5% isopropanol, and 0.10 M HCl). Rapid colorimetric assay for cellular growth and survival was recorded using a microplate reader (Bio-Rad) in absorbance at 595 nm.
Colony formation assay
Infected and uninfected A549 cells were plated onto six-well plates at a density of 500 cells per well for 8 days with the culture medium changed regularly. Adherent cells were washed with PBS and fixed in 4% paraformaldehyde at room temperature for 10 minutes. The cells were stained with 0.5% crystal violet for 10 minutes, washed, and air-dried. The colonies were counted directly under a Zeiss fluorescence microscope.
Cell cycle analysis by flow cytometry
A549 cells following infection with Lv-shGGCT or Lv-shCon were grown in 6-cm dishes at a density of 100,000 cells per dish and incubated for 40 hours. After a wash with ice-cold PBS, the cells were fixed with 75% cold ethanol overnight at 4°C. The cells were then treated with propidium iodide (PI; 50 mg/mL; Sigma-Aldrich) supplemented with 100 mg/mL DNase-free RNase A and incubated for 30 minutes. The suspension was filtered through a 50-mm nylon mesh, and all stained cells were analyzed using a Cytomics FC 500 flow cytometer (Beckman Coulter).
Statistics and data analysis
The data are presented as mean±SD of three independent experiments. Student's t-tests were performed for statistical analysis of comparisons between groups. A p-value of <0.05 was considered significant.
Results
GGCT gene and protein expression profiling in human lung cancer cell lines
To investigate the expression patterns of GGCT in A549, H1299, and H460 lung cancer cells, the qRT-PCR was carried out. As shown in Figure 1A, H1299 cells showed the highest transcription level of GGCT, and A549 cells exerted moderate GGCT expression, while H460 cells exerted the lowest. Thereafter, the GGCT protein expression in these cell lines was detected by western blot. As shown in Figure 1B, A549 and H460 cells performed higher levels of GGCT protein, H1299 cells with the highest transcription level, but presented the lowest protein pattern. New insights into GGCT biological function in lung cancer were evaluated based on A549 cells with higher gene and protein expression.
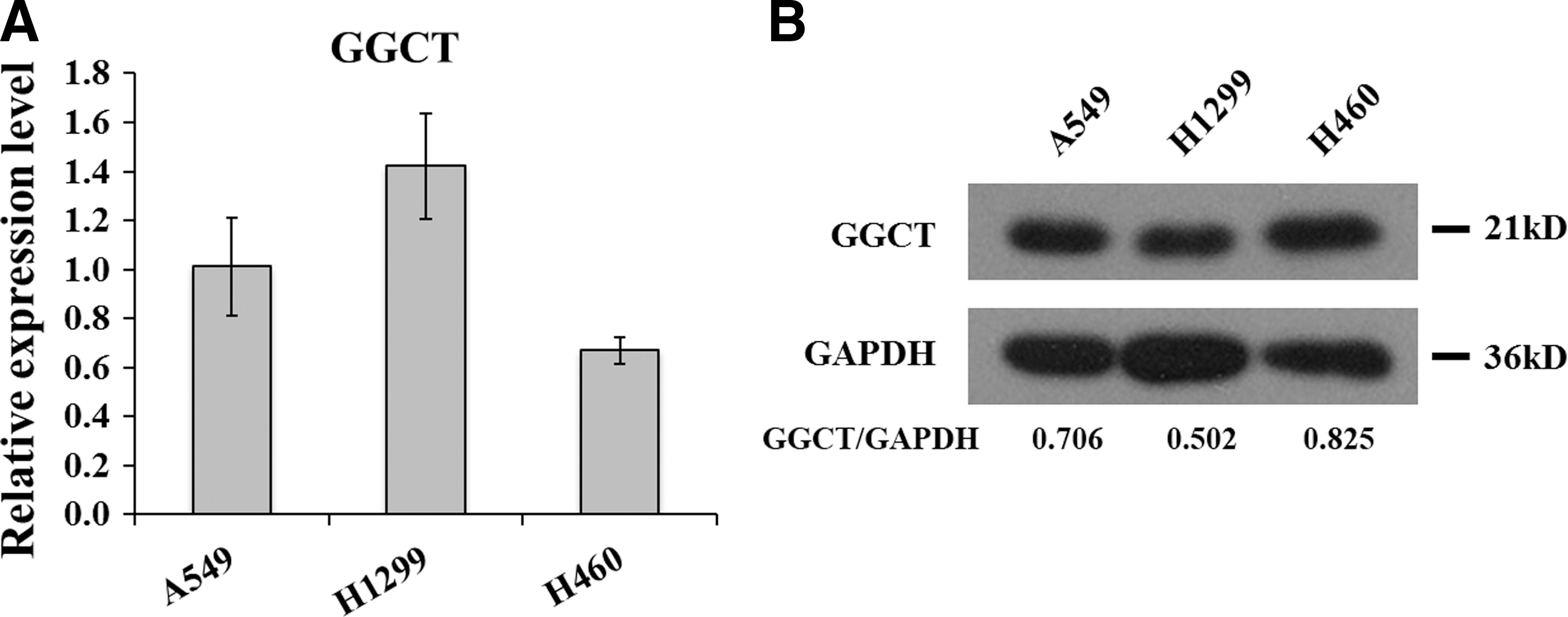
GGCT gene and protein expression profiling in A549, H1299, and H460 lung cancer cell lines.
Knockdown endogenous GGCT with shRNA lentiviruses in A549 cells
First, shRNA-expressing lentivirus directed against GGCT (Lv-shGGCT) was designed to knock down its expression in A549 cells. Additionally, A549 cells infected with lentiviral vectors expressing shRNA control were used as a negative control. The infection rate of A549 cells by Lv-shGGCT or Lv-shCon was more than 80%, as assessed by GFP fluorescence (Fig. 2A). To determine the knockdown efficiency of GGCT in A549 cells, the transcription and translation changes were identified by qRT-PCR and western blot. From Figure 2B, the authors could see that the GGCT mRNA level was downregulated significantly in the Lv-shGGCT group with a reduction of 60% compared with the Lv-shCon group. The GGCT protein level was also dramatically reduced in the Lv-shGGCT-infected cells in comparison with the control groups (Fig. 2C). In addition, the expression levels of GGCT were obviously decreased in both H1299 and H460 cell lines after Lv-shGGCT infection (Supplementary Fig. S1A; Supplementary Data are available online at
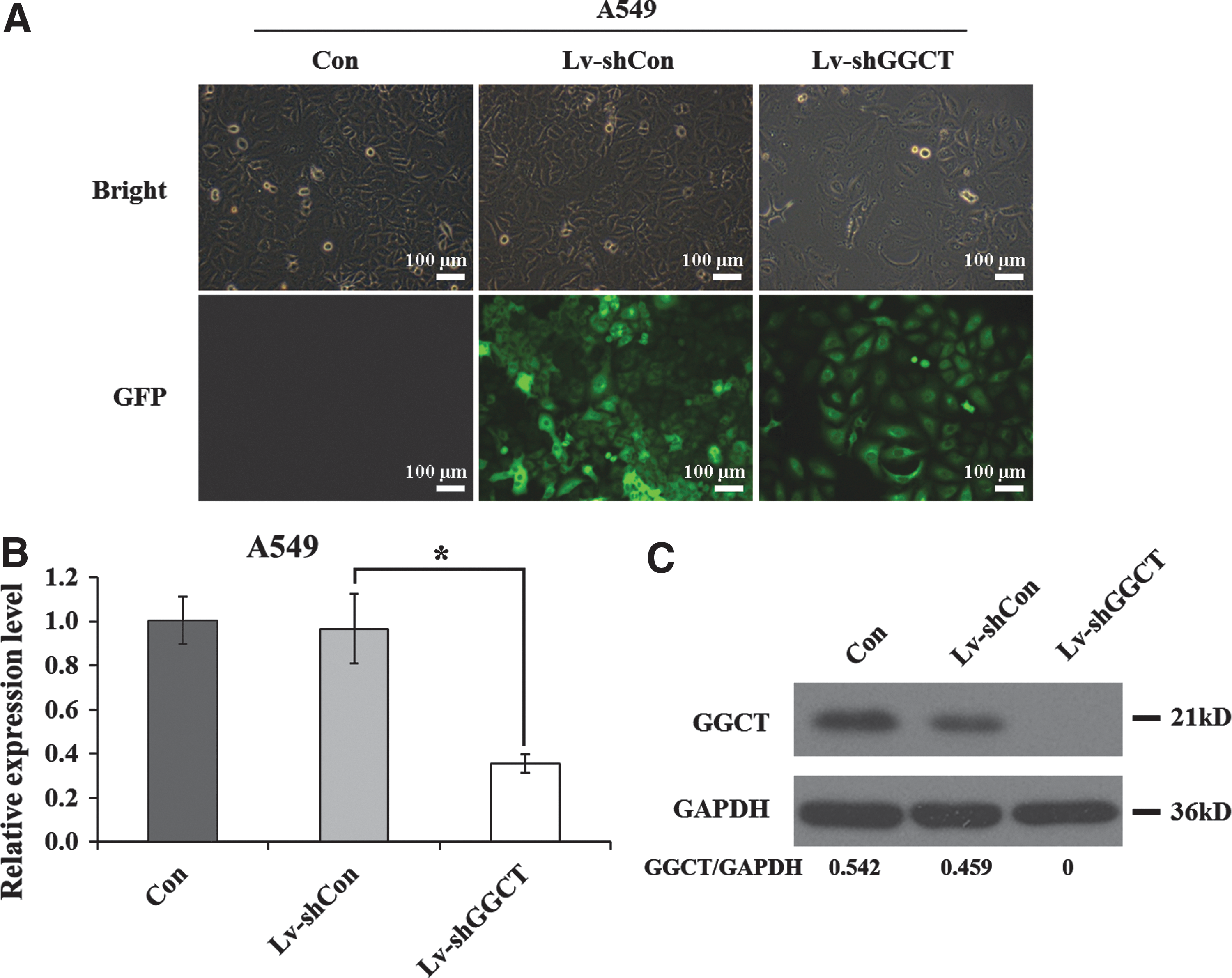
Knockdown efficiency of GGCT in A549 lung cancer cells.
GGCT knockdown is correlated with reduced proliferation and colony formation
Further investigations were conducted to explore the biological function of GGCT in cell proliferation and colony formation. As shown in Figure 3A, Lv-shGGCT infection in A549 cells could significantly inhibit the proliferation ability, as measured by MTT assay. The proliferation rate in the Lv-shGGCT group was reduced by 46.1% compared with the Lv-shCon group on the fourth day. On the fifth day, the proliferation rate was decreased by 50.8%. However, there was no difference between Lv-shCon-infected and noninfected A549 cells. Similarly, the growth curves of Lv-shGGCT groups were markedly lower than those of Lv-shCon groups in both H1299 and H460 cell lines (Supplementary Fig. S1B, C). These results indicated that GGCT knockdown could obviously suppress the proliferation of lung cancer cells.
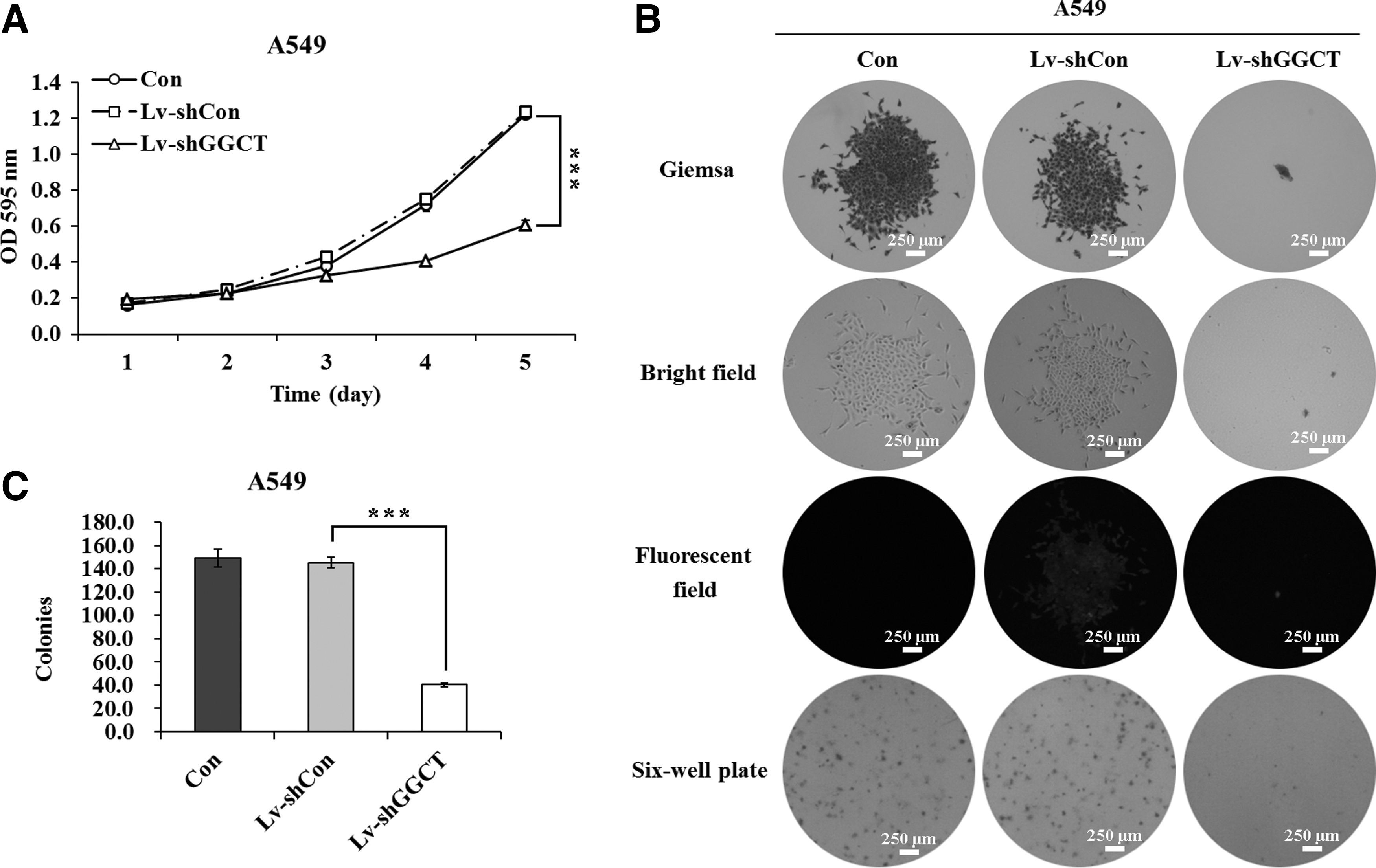
The cell proliferation and colony formation were inhibited by GGCT knockdown in A549 cells.
To determine whether GGCT knockdown affects the colony-forming ability of A549 cells, the colony formation assay was carried out. The authors can see from Figure 3B that both the size and number of colonies were dramatically reduced in the experimental group with Lv-shGGCT infection in contrast with controls. The number of colonies was significantly reduced by 73.2% and 72.4% after Lv-shGGCT infection compared with Con and Lv-shCon groups, respectively (Fig. 3C). These results indicated that GGCT knockdown could significantly inhibit the colony-forming ability of A549 cells.
GGCT knockdown blocks cell cycle progression of A549 cells
To measure the effect of Lv-shGGCT infection on cell cycle progression in A549 cells, flow cytometry was performed (Fig. 4A). As a result, GGCT knockdown arrested A549 cells in the G0/G1 phase. The cell percentage of the G0/G1 phase in Lv-shGGCT-infected cells increased by 31.2% in contrast with Lv-shCon-infected cells, while the cell percentage of the S phase in Lv-shGGCT-infected cells was decreased by 52.3%, and the G2/M phase was reduced by 21.6% in contrast with Lv-shCon-infected cells (Fig. 4B). However, no significant difference in percentage of cells was observed between the groups of Lv-shCon and control during G0/G1, S, and G2/M phases. These results indicated that GGCT knockdown in A549 cells led to G0/G1 cell cycle arrest. Furthermore, the expression alterations of G0/G1-associated markers were detected in A549 cells by western blot. As shown in Figure 4C, knockdown of GGCT resulted in decreases in CDK4, CDK6, and cyclin D1 expression, particularly absolute downregulation of CDK6. Besides, the cleaved level of PARP was obviously increased in the Lv-shGGCT group, which suggested that knockdown of GGCT might also induce cell apoptosis through cleavage of PARP.
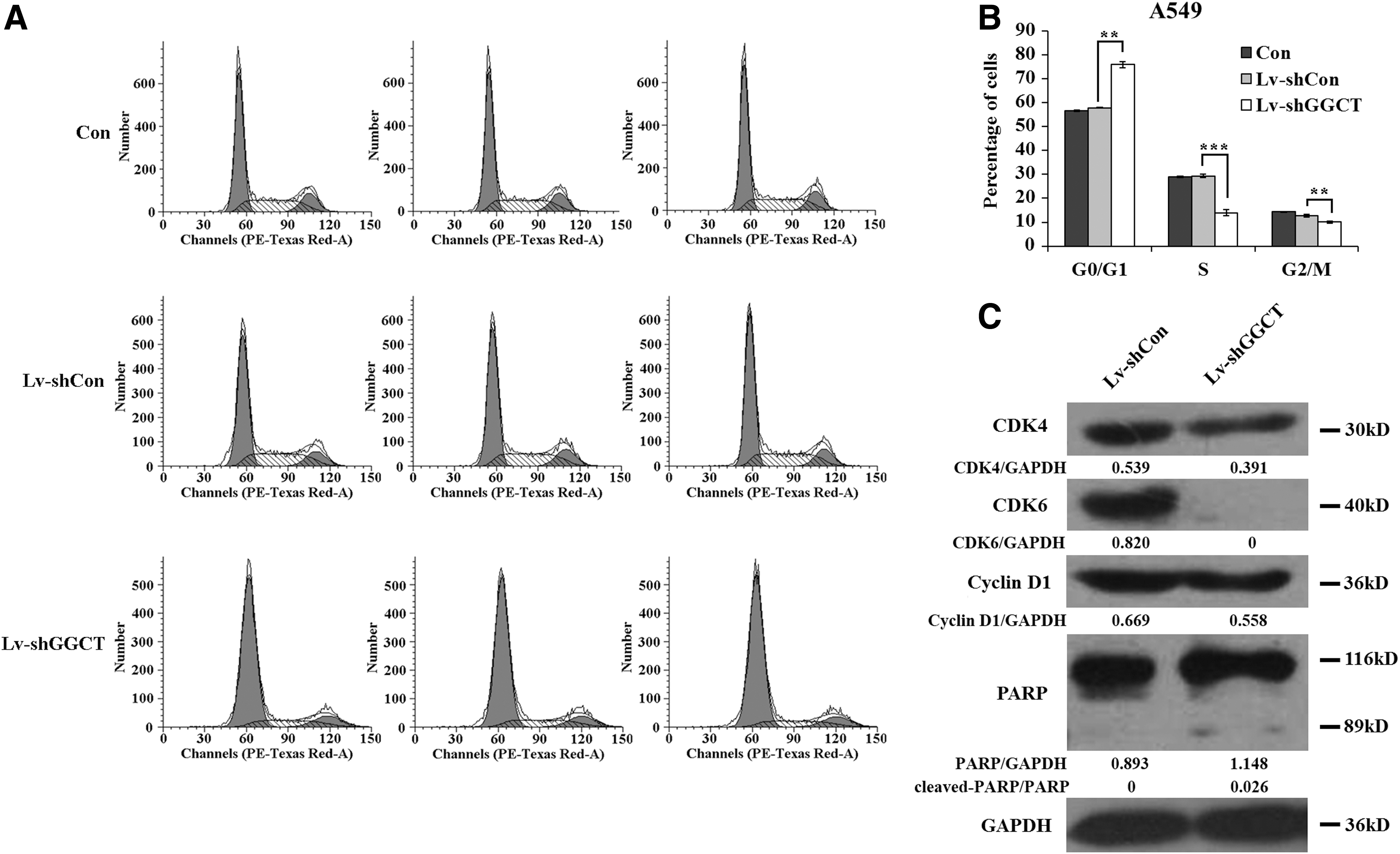
GGCT knockdown induces cell cycle arrest at the G0/G1 phase in A549 cells.
Discussion
Despite progress in traditional therapy of lung cancer, it remains the most common cancer around the world. 20 An emerging field of cancer gene therapy, which uses genetic material to modify cells, can provide a large number of innovative treatments. 21 Target gene knockdown as a cancer gene therapy strategy can pave the way toward the development and therapeutic approaches for the treatment of cancer. GGCT is an enzyme involved in the γ-glutamyl cycle, which results in glutathione synthesis and degradation and can transport amino acids across cellular membranes. 15 Furthermore, it is a potential diagnostic marker for various types of cancer; however, its implication for lung cancer is unclear. In this study, the authors identified that GGCT knockdown inhibited lung cancer cell proliferation and colony formation. These data were coincident with a recent report that downregulation of GGCT expression suppressed the growth of osteosarcoma cells. 17
Altered cell cycle distribution contributes to inhibition of cancer cell growth. For instance, AEG-1 expression knockdown suppressed neuroblastoma cell proliferation through blockade of cell cycle progression from the G0/G1 to S phase. 22 RNA interference-mediated silencing of iASPP functions to constrain cell proliferation and colony formation by G0/G1 block in U251 human glioblastoma cells. 23 Ohno et al. 24 showed that GGCT is a novel cell cycle-associated gene. This study showed that knockdown of GGCT arrested A549 lung cancer cells in the G0/G1 phase, which suggests that GGCT knockdown inhibited G0/G1-to-S phase transition to suppress cell proliferation. Cyclins and cdks are two types of crucial regulatory molecules that determine cell cycle progression. 25 Cyclin D1 binding to CDK4/6 forms the active complex of cyclin D1-CDK4/6, which phosphorylates retinoblastoma protein (pRb) and subsequently releases E2F transcription factors, resulting in the activation of specific gene expression required for G0/G1-to-S phase progression. 26 Furthermore, the authors found that knockdown of GGCT decreased the expression of CDK4, CDK6, and cyclin D1, which suggested that GGCT knockdown induced G0/G1 phase arrest potentially through disruption of cyclin D1-CDK4/6 complex formation. In addition, an increased cleavage of PARP was observed in GGCT knockdown cells, suggesting an induction of apoptosis.
GGCT is an enzyme in the γ-glutamyl cycle and converts γ-glutamyl amino acids into 5-oxoproline and free amino acids. 14 The authors thus suggest that GGCT knockdown may disturb the γ-glutamyl cycle; however, the potential correlation between the disordered cycle of γ-glutamyl and cell division needs further investigation. On the other hand, downregulation of GGCT may interfere with the transportation of amino acids, which are necessary for cell division processes and more research will be conducted.
In conclusion, GGCT knockdown could restrain the proliferation and colony formation of lung cancer cells. This investigation reveals the implication between GGCT expression and lung cancer cell proliferation and may provide an appropriate therapeutic target against lung cancer.
Footnotes
Acknowledgments
This study was supported by the Shanghai Natural Science Foundation (No. 13ZR1433000) and The Research project of Shanghai Bureau of Health (No. 20124350).
Disclosure Statement
No conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.