
Abstract
Human osteosarcoma (OS) represents one of the most common primary sarcomas often originating in the metaphyses of long bones. However, its underlying molecular pathogenesis is still only vaguely understood. Several tight junction proteins were shown to be associated with and involved in tumorigenesis. This study is aimed to evaluate the role of Claudin 8 (CLDN8) in human OS. Lentivirus-based short hairpin RNA targeting CLDN8 specifically depleted its endogenous expression in U2OS and SW1353 OS cells, with a reduction by 97.7% and 89.3%, respectively, in contrast to control. Depletion of CLDN8 led to a significant diminution in cell viability and proliferation. To test the mechanism by which CLDN8 modulates cell proliferation, the flow cytometry assay and apoptosis assay were performed and confirmed that G1-S transition was blocked and a strong proapoptotic effect was induced in U2OS cells by CLDN8 knockdown. These data demonstrate that CLDN8 plays an essential role in OS proliferation in vitro, which will provide a new opportunity for discovering and identifying novel effective treatment strategies.
Introduction
Osteosarcoma (OS), also known as osteogenic sarcoma, is the most frequently occurring primary malignant bone tumor after plasmacytoma. 1 It has an incidence of four to five cases per million worldwide and is a leading cause of cancer-related death in adolescents and young adults. 2 With the combination of surgery and chemotherapy, 50%–60% of patients with a localized OS will achieve long-term disease-free survival, while the long-term survival for patients with metastasis at diagnosis is only 20%–30%. 3 Therefore, there is urgent need to identify the genetic risk factor for OS.
Tight junctions are located near the apical surface and consist of a cytoplasmic plaque of proteins as well as intracellular protein fibers. 4,5 As integral membrane proteins, claudins form tight junctions together with occludin. Claudins are tetraspan proteins consisting of a family of at least 26 members ranging in molecular mass from 20 to 28 kDa. 6 In mammals, the expression domains of Claudins 1–4, Claudins 6–11, and Claudins 14–16 are restricted to one or several nephron segments. 7 In the mouse nephron, Claudin 8 (CLDN8) is restricted to the proximal tubule (PST), distal tubule, and collecting duct, 8,9 while in the human nephron, CLDN8 has been colocalized thus far with distal markers. 10 Recently, McKee et al. 11 discovered that the zebrafish ortholog, CLDN8, is expressed in the PST and across the distal pronephros segments and duct. It has been reported that CLDN8 plays important roles in the paracellular cation barrier of the distal renal tubule and in the paracellular barrier to prevent sodium back-leakage in the distal colon. 12 Tight junctions are the major determinants of paracellular permeability, which, in the gut, protect the body from entry of harmful substances such as microbial components. Disrupted tight junctions have been observed in the small intestine of cystic fibrosis mice, in which CLDN1, CLDN7, CLDN8, and` PMP22 expressed at lower levels and CLDN2 expressed at a higher level. 13 Bioinformatic analysis revealed that CLDN8 was downregulated in inflammatory bowel disease. 14 Furthermore, CLDN8 also plays a role in the spermatogonial stem cell niche. 15
Several claudins were shown to be involved in various cancer types. CLDN1 and CLDN12 were frequently overexpressed in colorectal cancer, whereas CLDN8 showed downregulation in tumor tissue on the RNA level. 16 However, CLDN8, along with CLDN7, is identified as an immunohistochemical marker for the differential diagnosis of chromophobe renal cell carcinoma and renal oncocytoma. 17,18 To date, the role of CLDN8 in human OS remains unknown. In this study, to investigate whether CLDN8 is involved in OS growth, endogenous CLDN8 was depleted in U2OS and SW1353 OS cells by lentivirus-based RNA interference (RNAi), and the effects of CLDN8 silencing on cell viability, proliferation, and cell cycle were further investigated.
Materials and Methods
Materials
Dulbecco's modified Eagle's medium (DMEM) was obtained from Hyclone (cat no. SH30243.01B+). Fetal bovine serum (FBS) was obtained from Biological Industries (Cat No. 04-001-1A). Lipofectamine 2000 and TRIzol® Reagent were purchased from Invitrogen. Moloney Murine Leukemia Virus (M-MLV) Reverse Transcriptase was purchased from Promega. All other chemicals were obtained from Sigma-Aldrich. The lentiviral vector (pFH-L) and packaging vectors (pVSVG-I and pCMVΔR8.92) were purchased from Shanghai Hollybio. The antibodies used were as follows: rabbit anti-CLDN8 antibody (1:500 dilution, Cat No. AP9222c-ev; Abgent), rabbit anti-CDK2 antibody (1:1000 dilution, Cat No. #2546; Cell signaling), rabbit anti-GAPDH antibody (1:100,000 dilution, Cat No. 10494-1-AP; Proteintech Group, Inc.), and horseradish peroxidase-conjugated goat antirabbit (1:5000 dilution, Cat No. SC-2054; Santa Cruz) secondary antibodies.
Cell culture
Human OS cells U2OS, SW1353, and human embryonic kidney cells 293T were purchased from the Cell Bank of Chinese Academy of Science (Shanghai, China). All cell lines were maintained in DMEM supplemented with 10% FBS at 37°C in humidified atmosphere of 5% CO2.
Lentivirus-delivered short hairpin RNA transduction
The short hairpin RNA (shRNA) sequence targeting the human CLDN8 gene (NCBI Accession Number: NM_199328.2) is 5′- TGATTCCCTGCTGGCTCTTTCCTCGAGGAAAGAGCCAGCAGGGAATCA-3′, which has been subjected to BLAST analysis against the human genome database to eliminate cross-silence phenomena with nontarget genes. A scrambled fragment (5′- GCGGAGGGTTTGAAAGAATATCTCGAGATATTCTTTCAAACCCTCCGCTTTTTT-3′) that has no significant homology with mouse or human gene sequences was used as a negative control. The shRNAs were cloned into the pFH-L vector, which was then transfected into 293T cells with packaging vectors (pVSVG-I and pCMVΔR8.92) using Lipofectamine 2000, according to the manufacturer's instructions. The supernatant was collected 48 hours later, centrifuged (4000 g, 4°C, 10 min) to remove cell debris, filtered through 0.45-μm cellulose acetate filters, and then concentrated again (4000 g, 4°C, 15 min). U2OS and SW1353 cells were dispensed into six-well plates at a density of 40,000 cells per well and transduced with shRNA-expressing lentivirus (shCon or shCLDN8) at a multiplicity of infection of 20. The lentiviral vectors expressed green fluorescence protein (GFP), which allowed for measurement of infection efficiency in transduced cells.
Quantitative real-time PCR analysis
U2OS and SW1353 cells were harvested after lentivirus transduction for 5 days. Total cellular RNA was extracted using TRIzol reagent and reversely transcripted to cDNA by M-MLV reverse transcriptase, according to the manufacturer's instructions. Quantitative Real Time PCR (qPCR) products were detected with SYBR Green on the BioRad Connet Real-Time PCR platform. The qPCR procedure was as follows: denaturation at 95°C for 1 min, 40 cycles of denaturation at 95°C for 5 seconds, and extension at 60°C for 20 seconds. The β-actin gene was amplified as internal control. Relative quantitation was analyzed by taking the difference ΔC(T) between the C(T) of β-actin and C(T) of the target gene and computing 2−ΔΔC(T). The following primers were used:
CLDN8,
5′-CCGTGATGTCCTTCTTGGCTTTC-3′ (forward)
5′-CTCTGATGATGGCATTGGCAACC-3′ (reverse)
β-actin,
5′-GTGGACATCCGCAAAGAC-3′ (forward)
5′-AAAGGGTGTAACGCAACTA-3′ (reverse)
Western blotting analysis
U2OS and SW1353 cells were harvested after lentivirus transduction for 5 days. Total protein was extracted with 2 × sodium dodecyl sulfate (SDS) sample buffer (100 mM Tris-HCl [pH 6.8], 10 mM EDTA, 4% SDS, 10% Glycine). Equal amounts of lysate (30 μg) in each lane, as determined by the bicinchoninic acid assay, were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes. The membranes were blocked with 5% nonfat dry milk in Tris-buffered saline with Tween 20 (TBST) for 1 hour at room temperature, and incubated with TBST containing anti-CLDN8 or anti-GAPDH antibody overnight at 4°C, followed by incubation with secondary antibody for 1 hour at room temperature. The blots were detected using the enhanced chemiluminescence (ECL) kit (Amersham) and visualized by exposure to X-ray film. GAPDH was used as a control to verify equal protein loading.
MTT viability assay
To evaluate the effect of CLDN8 on the viability of OS cells, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) colorimetric assay was performed in U2OS and SW1353 cells after lentivirus transduction for 3 days. Briefly, the OS cells were dispensed into 96-well plates at a concentration of 2000 per well. The plates were incubated for 1–5 days at 37°C. On each day, 20 μL of MTT solution (5 mg/mL) was added and incubated for 4 hours. Afterward, the entire supernatant was discarded and acidic isopropanol (10% SDS, 5% isopropanol, and 0.01 M HCl) was added at a volume of 100 μL per well and incubated overnight at 37°C. The absorbance at 595 nm of each well was determined using an enzyme-linked immunosorbent assay (ELISA) reader.
Colony formation assay
To evaluate the effect of CLDN8 on the long-term proliferation of OS cells, colony formation assay was performed in U2OS cells after lentivirus transduction for 4 days. Briefly, U2OS cells were dispensed into six-well plates at a concentration of 500 per well. The culture medium was changed at 3-day intervals. U2OS cells were cultured for 9 days until the most single colony contains more than 50 cells. The colonies were stained with crystal violet for 3 min, and then washed with water and air-dried. Cell colonies were captured and counted under a microscope.
Fluorescence-activated cell sorting analysis
To evaluate the effect of CLDN8 on OS cell cycle progression, flow cytometry assay was performed in U2OS cells after lentivirus transduction for 7 days. Briefly, U2OS cells were dispensed into 6-cm dishes at a concentration of 80,000 per dish. After culture at 37°C for 40 hours, cells were harvested, fixed in 70% ethanol, and stored overnight at 4°C. The cells were then treated with NaCl/Pi staining solution (50 μg/mL PI and 100 μg/mL RNase A). Following incubation for 1 hour in the dark at room temperature, cells were analyzed by flow cytometry (FACS Calibur; Becton Dickinson). The fractions of the cells in G0/G1, S, and G2/M phases were analyzed with dedicated software (Becton Dickinson).
Cell apoptosis assay
U2OS cells were treated with two different ways (shCon and shCLDN8) before 96 hours of incubation. Subsequently, the cells were washed with PBS and reinoculated in 6-cm dishes at a density of 8 × 104 cells/dish. After 48 hours, the cells were collected, washed with PBS again thrice, and dealt with Annexin V/7-AAD double staining following the instructions of the Apoptosis Assay Kit (KeyGEN Biotech). Cells were analyzed by flow cytometry (FACS Calibur; Becton Dickinson).
Statistical analysis
Statistical analysis was performed using the SPSS 16.0 software package (SPSS, Inc.). All data are expressed as mean ± standard deviation. Differences between two groups were analyzed by Student's t-test, and a p-value of <0.05 was considered statistically significant.
Results
Knockdown of CLDN8 expression with lentivirus-delivered shRNA
U2OS and SW1353 cells were transduced with shRNA-expressing lentivirus (shCon or shCLDN8). GFP expression was observed by fluorescent microscopy 3 days post-transduction. As depicted in Figure 1A and Supplementary Figure S1A (Supplementary Data are available online at
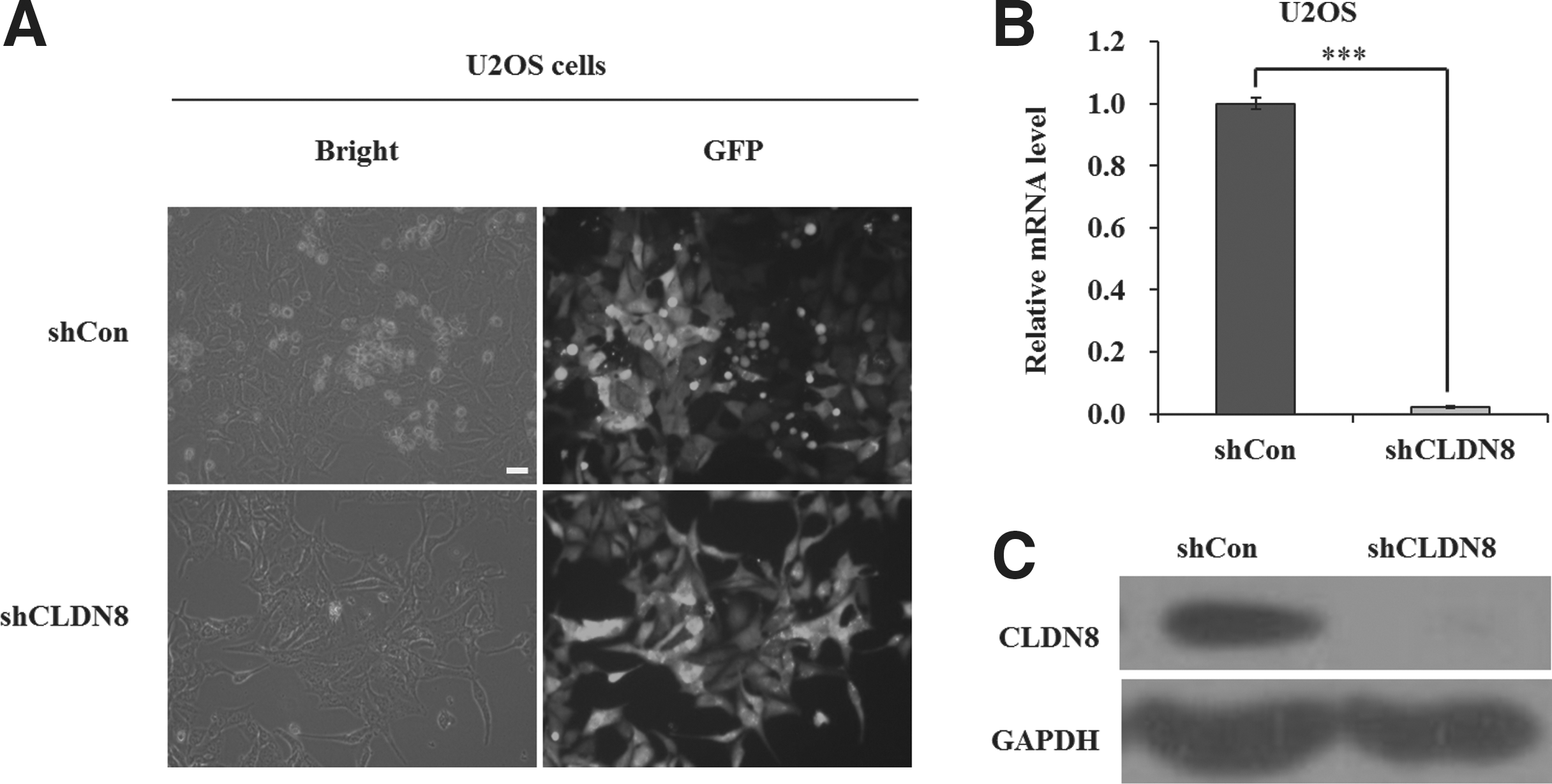
Lentivirus-delivered short hairpin RNA (shRNA) targeting CLDN8 depleted its endogenous expression in U2OS cells.
Effect of CLDN8 knockdown on cell proliferation and colony formation
The authors next examined the effects of CLDN8 knockdown on viability and proliferation of U2OS and SW1353 cells. After infection of CLDN8 shRNA, MTT assay was performed in U2OS and SW1353 cells for 5 consecutive days. As depicted in Figure 2A and Supplementary Figure S2, the number of viable cells infected with shCLDN8 was much lower than those infected with shCon (***p < 0.001). In addition, colony formation assay was also performed in U2OS cells and representative photographs are presented as Figure 2B. Crystal violet staining and fluorescence expression showed that both the size of monoclone and the number of total colonies were decreased in shCLDN8-infected cells. The colony formation ability was significantly impaired, with a reduction by 88.6% in U2OS cells after CLDN8 knockdown (p < 0.001, Figure 2C). These indicate that knockdown of CLDN8 could strongly reduce viability and proliferation of OS cells.
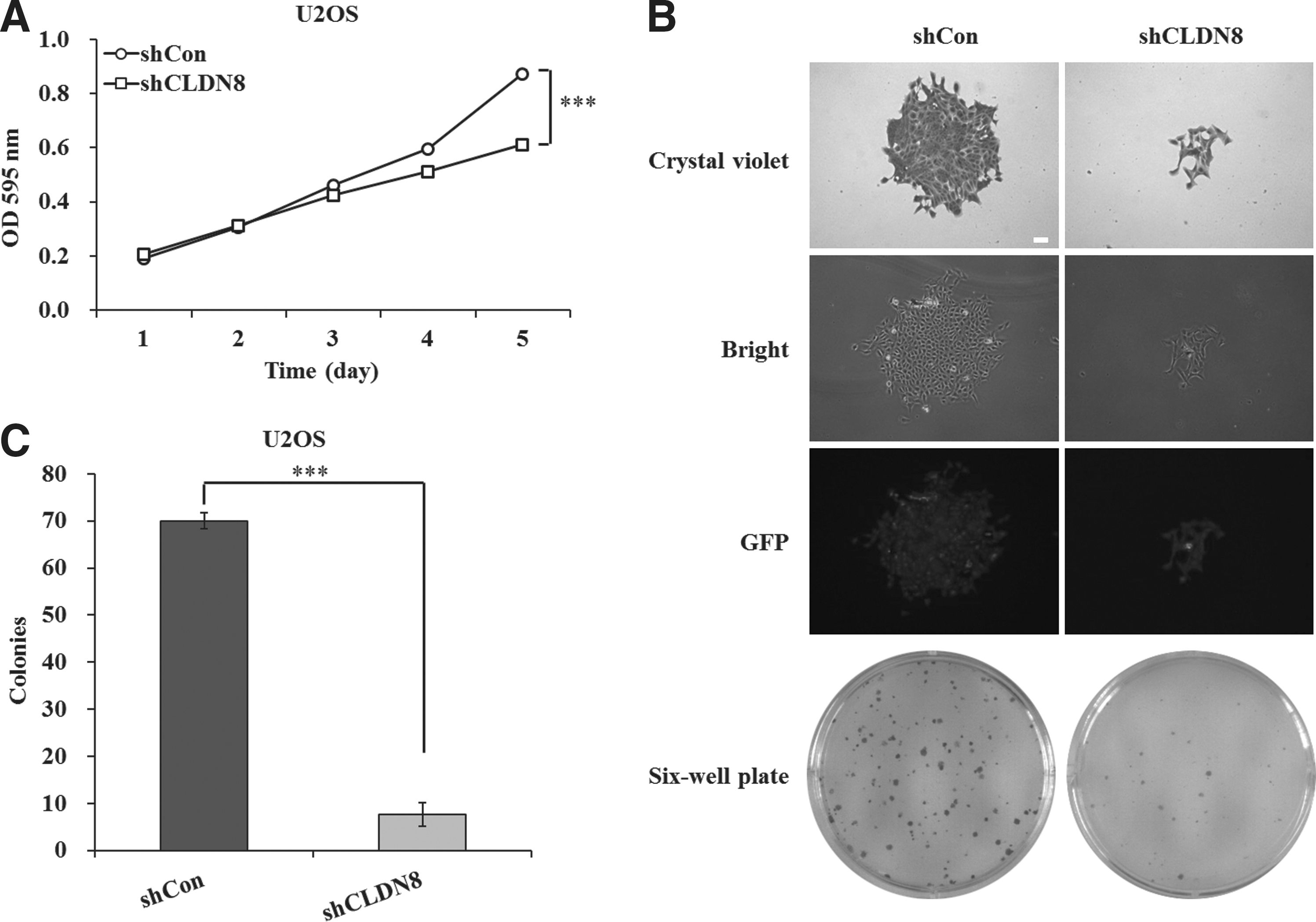
Knockdown of CLDN8 inhibits viability and proliferation of U2OS cells.
Effect of CLDN8 knockdown on cell cycle progression
To test the mechanism by which CLDN8 modulates cell proliferation, the flow cytometry assay was used to determine the cell cycle of U2OS cells. Representative images of cell cycle distribution are presented as Figure 3A. As depicted in Figure 3B, the cell percentage of the G0/G1 phase was increased from 59.82% ±0.51% in shCon-infected cells to 69.07% ±0.44% in shCLDN8-infected cells. By contrast, the cell percentage of the S phase was decreased from 26.88% ±0.65% in shCon-infected cells to 16.96% ±0.34% in shCLDN8-infected cells. This indicates that CLDN8 shRNA could strongly block (p < 0.001) the cell cycle progression of U2OS cells. Taken together, these results suggest that knockdown of CLDN8 could inhibit U2OS cell growth by inducing G0/G1 phase cell cycle arrest.
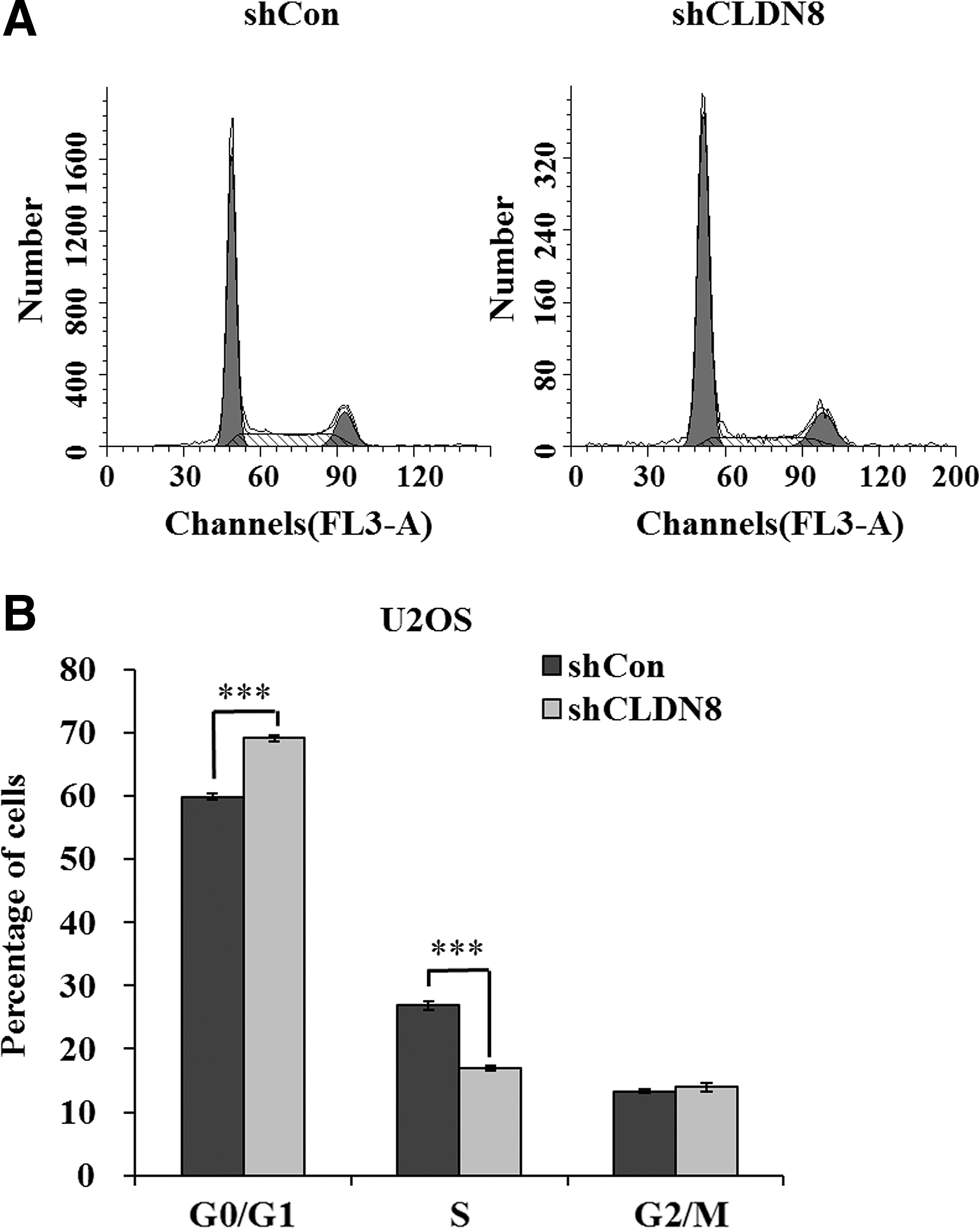
Knockdown of CLDN8 arrests cell cycle progression in U2OS cells.
Depletion of CLDN8 promoted U2OS cell apoptosis
To explore the effect of CLDN8 knockdown on cell apoptosis, the authors used Annexin V-APC/7-AAD double staining on infected U2OS cells. Annexin V versus 7-AAD plots showed the percentages of viable (Annexin V−/AAD−), necrotic (Annexin V–/7-AAD+), early apoptotic (Annexin V+/7-AAD–), and late apoptotic (Annexin V+/7-AAD+) cells, respectively. As is shown in Supplementary Figure S3, the percentage of early apoptotic cells was obviously increased (***p < 0.001) after infection, and the percentage of the whole apoptotic cells (early and late) was also markedly increased. Therefore, these results demonstrated that downregulation of the CLDN8 gene could cause a strong proapoptotic effect in U2OS cells.
Depletion of CLDN8 interfered with the expression of some cell cycle regulators of U2OS cells
To explore the mechanism linking CLDN8 expression and cell cycle, the authors detected the expression of some cell cycle regulators and found that the expression of CDK2 was decreased after the knockdown of CLDN8 (Supplementary Fig. S4). CDK2 is a protein kinase of the CDK family, which plays an important role in cell cycle progression. The activity of Cdk2/cyclin E kinase is important for the G1 to S transition. Thus, the reduction of CDK2 after inhibition of CLDN8 is corresponding to the result of the former cell cycle analysis.
Discussion
An abnormal distribution of tight junctions has been observed in primitive sarcoma, synovial sarcoma. 19 Many of the previously described cell–cell junctions in Ewing's sarcoma/primitive neuroectodermal tumor are poorly formed tight junctions given the high frequency of CLDN1 and ZO1 expression. 20 These studies indicated that tight junction dysregulation may be associated with primary sarcomas. However, whether tight junction proteins change and participate in development of OS has not yet been investigated. In this study, the authors aimed to determine the role of CLDN8, which has been identified as a candidate biomarker in renal oncocytoma, in human OS. They provide new evidence demonstrating that CLDN8 may be a potential tumor marker in human OS.
Knockdown of endogenous CLDN8 expression by shRNA-expressing lentivirus significantly decreased the viability and proliferation of U2OS and SW1353 OS cells. Moreover, fluorescence-activated cell sorting (FACS) analysis showed that knockdown of CLDN8 induced a significant arrest in the G0/G1 phase of the cell cycle and cell apoptosis assay demonstrated a strong proapoptotic effect caused by the depletion of CLDN8, which were helpful for the inhibition of cell proliferation. Therefore, it is likely that CLDN8 could modulate OS growth through the regulation of a subset of cancer-related genes involved in the G1-S checkpoint. In addition, the cell cycle regulator, CDK2, was detected as reduced in U2OS cells after infection, which corresponds to the results of FACS. Further investigations are needed to reveal the molecular basis for the oncogenic function of CLDN8 in OS cells.
In all, knockdown expression of CLDN8 by RNAi significantly inhibits the growth of U2OS cells by inducing cell cycle arrest. These findings demonstrate for the first time that CLDN8 plays an essential role in cell growth. Furthermore, gene therapy is a potential research direction in the treatment of cancer. 21 As a newfound related gene of OS, CLDN8 might be used as the target for the treatment of OS in the future. However, more investigations toward the clinical usage of CLDN8 depletion in human OS are still necessary.
Footnotes
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.