
Abstract
Galectin-1 is a member of carbohydrate-binding proteins and plays critical roles in tumor growth and progression. It has been reported that galectin-1 is upregulated in human hepatocellular carcinoma (HCC) and facilitates HCC cell migration and invasion. In this study, the authors aimed to explore the effects of the knockdown of galectin-1 on HCC cell survival and sensitivity to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). Human HCC cells were transfected with galectin-1-targeting small interfering RNA (siRNA) with or without 100 ng/mL TRAIL treatment and tested for apoptosis and gene expression changes. Cotransfection of Bcl-2- and survivin-expressing plasmids with galectin-1 siRNA was done, before TRAIL exposure, cell viability, and apoptosis were assessed. The authors found that siRNA-mediated downregulation of galectin-1 caused apoptosis in HCC cells, which was coupled with reduced Bcl-2 and survivin and increased Bax expression. Overexpression of Bcl-2 and survivin significantly blocked galectin-1 silencing-induced apoptosis of HCC cells. Knockdown of galectin-1 significantly enhanced TRAIL cytotoxicity against HCC cells, as determined by the MTT assay. Moreover, galectin-1 downregulation significantly induced apoptosis in TRAIL-treated HCC cells. Such effects were almost completely counteracted by the enforced expression of Bcl-2 and survivin. Taken together, these data first show that galectin-1 downregulation induces apoptosis in and augments TRAIL cytotoxicity to HCC cells largely through regulation of Bcl-2 and survivin expression. These findings provide a rationale for preclinical and clinical evaluation of targeting galectin-1 for improving TRAIL-based therapy against HCC.
Introduction
Hepatocellular carcinoma (HCC) is the third leading cause of cancer-related mortality worldwide, with a 5-year survival rate of 5%–6%. 1,2 The poor prognosis of HCC is mainly due to the lack of effective treatment options. Tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) is a member of the TNF family of cytokines. TRAIL has become an attractive anticancer agent, because it can selectively induce apoptosis in cancer cells with minimal toxicity to normal cells. 3,4 TRAIL exerts its proapoptotic activity through the binding to death receptors, DR4 and DR5, expressed on the surface of target cells. 5 When TRAIL binds to DR4 and/or DR5, the death receptors become trimerized and then recruit the adaptor protein, Fas-associated protein with death domain, and procaspase-8 or procaspase-10, leading to activation of caspase-8 or -10. Once activated, caspase-8 and -10 can activate the effector caspase-3, -6, and -7 to trigger apoptotic death. 6 HCC tissues have shown abundant expression of TRAIL, whereas adjacent nontumorous tissues have little expression of this cytokine. 7 Compelling evidence indicates strong resistance to TRAIL cytotoxicity in HCC cells. 5,6 Overexpression of antiapoptotic proteins and loss of proapoptotic proteins are important mechanisms leading to a poor apoptotic response to TRAIL. 8 The Bcl-2 family, consisting of antiapoptotic (e.g., Bcl-2 and Mcl-1) and proapoptotic (e.g., Bax and Bim) members, is widely involved in the regulation of apoptosis. 9 Deregulation of the Bcl-2 family proteins has been shown to modulate TRAIL-induced apoptosis in HCC cells. 10 Many attempts have been made to enhance TRAIL's therapeutic efficacy against cancer. 5
Galectin-1, a member of carbohydrate-binding proteins, is expressed by many types of cancers, such as nonsmall-cell lung cancer 11 and ovarian cancer. 12 Galectin-1 is involved in cancer progression and affects multiple aspects of tumor biology, including cell proliferation, adhesion, migration, and invasion. 12,13 Overexpression of galectin-1 has been detected in HCC and it contributes to tumor growth in vivo. 14 Another study reported that galectin-1 overexpression promotes HCC cell migration and invasion. 15 However, the role of galectin-1 in HCC cell survival and TRAIL resistance remains unclear.
In the present study, the authors employed a small interfering RNA (siRNA) to knock down galectin-1 in HCC cells and investigated its effects on cell apoptosis and sensitivity to TRAIL.
Materials and Methods
Cell culture and treatment
Human HCC cells (SK-Hep1 and Huh7) were purchased from the Shanghai Institute for Biological Science (Shanghai, China) and maintained in a high-glucose Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin (Invitrogen, Carlsbad, CA). For assessment of TRAIL cytotoxicity, cells were exposed to 100 ng/mL human recombinant TRAIL (Millipore, Billerica, MA) for 24 h, and cell viability and apoptosis were examined. This TRAIL concentration was chosen because it has been reported 16 that TRAIL at 100 ng/mL is cytotoxic to HepG2 HCC cells, but not normal hepatocytes.
Plasmids and siRNA
The human Bcl-2- and survivin-expressing plasmids in pcDNA3.1(+) were constructed as described previously. 17,18 In brief, full-length human Bcl-2 and survivin cDNA were amplified by polymerase chain reaction (PCR) using a human placenta cDNA library (Invitrogen) as a template. The PCR products were cloned into the pcDNA3.1(+) vector (Invitrogen) following the manufacturer's protocol. All constructs were sequenced to ensure accuracy. The green fluorescent protein (GFP)-expressing plasmid was purchased from Invitrogen. Specific siRNA-targeting human galectin-1 mRNA and negative control siRNA were purchased from Santa Cruz Biotechnology (Santa Cruz, CA).
Cell transfection
SK-Hep1 and Huh7 cells were seeded at a density of 8 × 105 cells per well into six-well plates and incubated overnight before transfection. For knockdown of galectin-1, galectin-1 siRNA and nonspecific siRNA, at a final concentration of 50 nM, were incubated with Lipofectamine 2000 (Invitrogen) at room temperature for 30 min and separately added to the cell culture. At 48 h after transfection, cells were collected and tested for gene expression and apoptosis. To study the role of Bcl-2 and survivin in galectin-1-mediated apoptosis, cells were cotransfected with galectin-1 siRNA (50 nM), pcDNA3.1-Bcl-2 (0.5 μg), and pcDNA3.1-survivin (0.5 μg) and incubated for 48 h before gene expression and apoptosis analysis. To explore the effect of galectin-1 on TRAIL sensitivity, cells were transfected with galectin-1 siRNA alone or together with Bcl-2- and survivin-expressing plasmids, 48 h before exposure to 100 ng/mL TRAIL. Transfection efficiency (>70%) was determined by cotransfection of a GFP-expressing plasmid and analysis of the percentage of GFP-positive cells by fluorescence microscopy.
Quantitative real-time PCR analysis
Total RNA was extracted from cells using a TRIzol reagent (Invitrogen). Reverse transcription was carried out using SuperScript II reverse transcriptase and random primers (Invitrogen). Real-time PCR was performed using FastStart DNA Master SYBR Green I reagents and with the LightCycler 480 Real-Time PCR System (Roche Diagnostic, Meylan, France). Primers sequences were as follows: galectin-1 forward 5′-CAAACCTGGAGAGTGCCTTC-3′, galectin-1 reverse 5′-GATGCACACCTCTGCAACAC-3′ 19 , β-actin forward 5′-CCCTGAGGCACTCTTCCAG-3′, β-actin reverse 5′-ACTTGCGCTCAGGAGGAGC-3′. Cycling conditions were as follows: an initial denaturation at 95°C for 10 min followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. Each reaction was done in triplicate. The relative amount of galectin-1 mRNA was calculated after normalization against β-actin mRNA levels using the comparative threshold cycle method. 20
MTT assay
Cells were plated at a density of 3 × 103 cells per well in 96-well plates and transfected with galectin-1 siRNA or control siRNA before TRAIL treatment. Afterward, cells were incubated with 3-(4, 5-dimethylthiazol-2yl)-2, 5-diphenyltetrazolium bromide (MTT) at a final concentration of 0.5 mg/mL at 37°C for 4 h. Dimethyl sulfoxide was added to dissolve the formazan product, and the absorbance was measured at 570 nm.
Apoptosis detection by flow cytometry
Cell apoptosis was detected using a commercially available AnnexinV-fluorescein isothiocyanate (FITC) Apoptosis Detection Kit (BioVision, Mountain View, CA), according to the manufacturer's instructions. In brief, cells were seeded at a density of 8 × 104 cells per well into 12-well plates and transfected with indicated plasmids or siRNAs. At 48 h after transfection, cells were trypsinized, washed with an ice-cold phosphate-buffered saline, and suspended in a 1× binding buffer. The cell suspension was added with 5 μL of AnnexinV-FITC and propidium iodide (PI) and incubated for 10 min at room temperature in the dark. Cells were analyzed using a FACSCalibur flow cytometer (BD Biosciences, San Jose, CA). At least 10,000 cells were analyzed per measurement.
Western blot analysis
Cells were transfected with indicated plasmids or siRNAs and 48 h later, lysed for 30 min in an ice-cold lysis buffer (50 mM Tris-HCl pH 8.0, 150 mM EDTA, 1% TritonX-100, and 0.5% sodium dodecyl sulfate [SDS]) containing a protease inhibitor mixture (Roche Diagnostics, Mannheim, Germany). The protein concentrations in the cell lysates were measured using the Protein Quantification Kit (Bio-Rad Laboratories, Hercules, CA). Equal amounts of protein were resolved on SDS–polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes. After blocking, the membranes were probed with the following primary antibodies: mouse or rabbit anti-galectin-1, anti-Bcl-2, anti-Bax, anti-survivin, anti-Mcl-1, anti-DR4, anti-DR5, and anti-β-actin, which were purchased from Santa Cruz Biotechnology. After washing, membranes were then incubated with appropriate secondary antibodies for 1 h. Immunoreactive bands were visualized with the enhanced chemiluminescent detection system (Amersham Biosciences, Piscataway, NJ). Densitometry was performed using the Quantity One software (Bio-Rad Laboratories).
Statistical analysis
Data are expressed as means ± standard deviations. The difference among the means of multiple groups was analyzed by the one-way analysis of variance followed by the Tukey test. p-Values < 0.05 were considered significant.
Results
siRNA-mediated downregulation of galectin-1 causes apoptosis in human HCC cells
The authors first checked the effect of galectin-1 downregulation on apoptosis of HCC cells. Real-time PCR and Western blot analysis revealed that the mRNA (Fig. 1A) and protein (Fig. 1B) levels of galectin-1 in SK-Hep1 and Huh7 cells were significantly (p < 0.05) reduced by transfection of galectin-1 siRNA, compared to nontransfected cells. Apoptosis analysis by Annexin-V/PI staining demonstrated that downregulation of galectin-1 significantly induced apoptosis in HCC cells. More specifically, apoptosis increased from 2.7 ± 0.8% to 18.5 ± 1.9% in SK-Hep1 cells and from 3.8 ± 0.6% to 20.2 ± 2.6% in Huh7 cells (Fig. 1C).
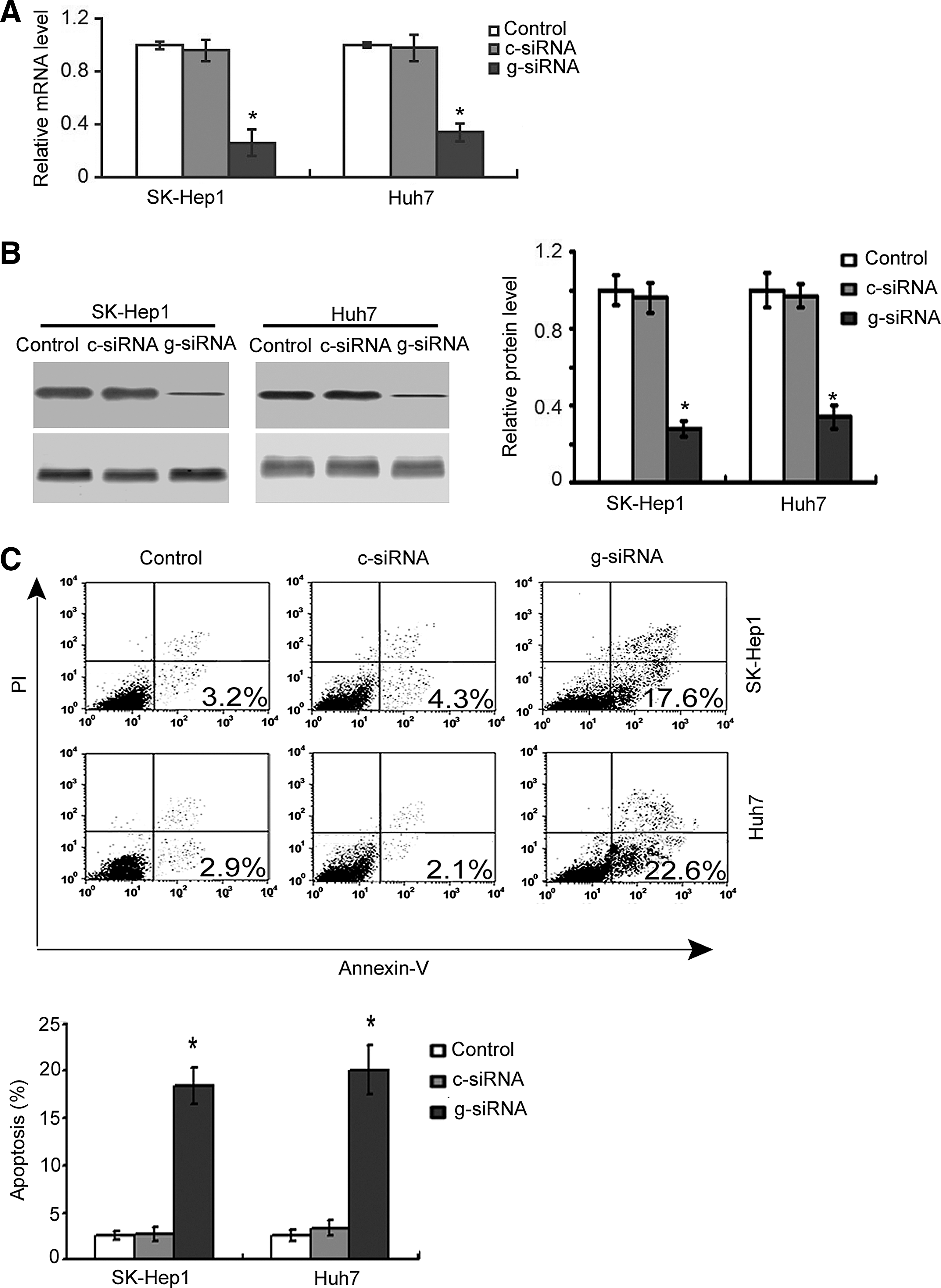
Knockdown of galectin-1 induces apoptosis in hepatocellular carcinoma (HCC) cells.
Effects of galectin-1 silencing on apoptosis regulators in human HCC cells
Having identified the requirement of galectin-1 for the survival of HCC cells, the authors next sought to determine the apoptosis regulators involved in galectin-1 downregulation-mediated cell death. Western blot analysis showed that SK-Hep1 and Huh7 cells with galectin-1 downregulation had a marked decline in the expression of Bcl-2 and survivin and elevation in the expression of Bax, compared to nontransfected cells (Fig. 2). However, galectin-1 silencing had no impact on the expression of Mcl-1 and TRAIL receptors (DR4 and DR5).
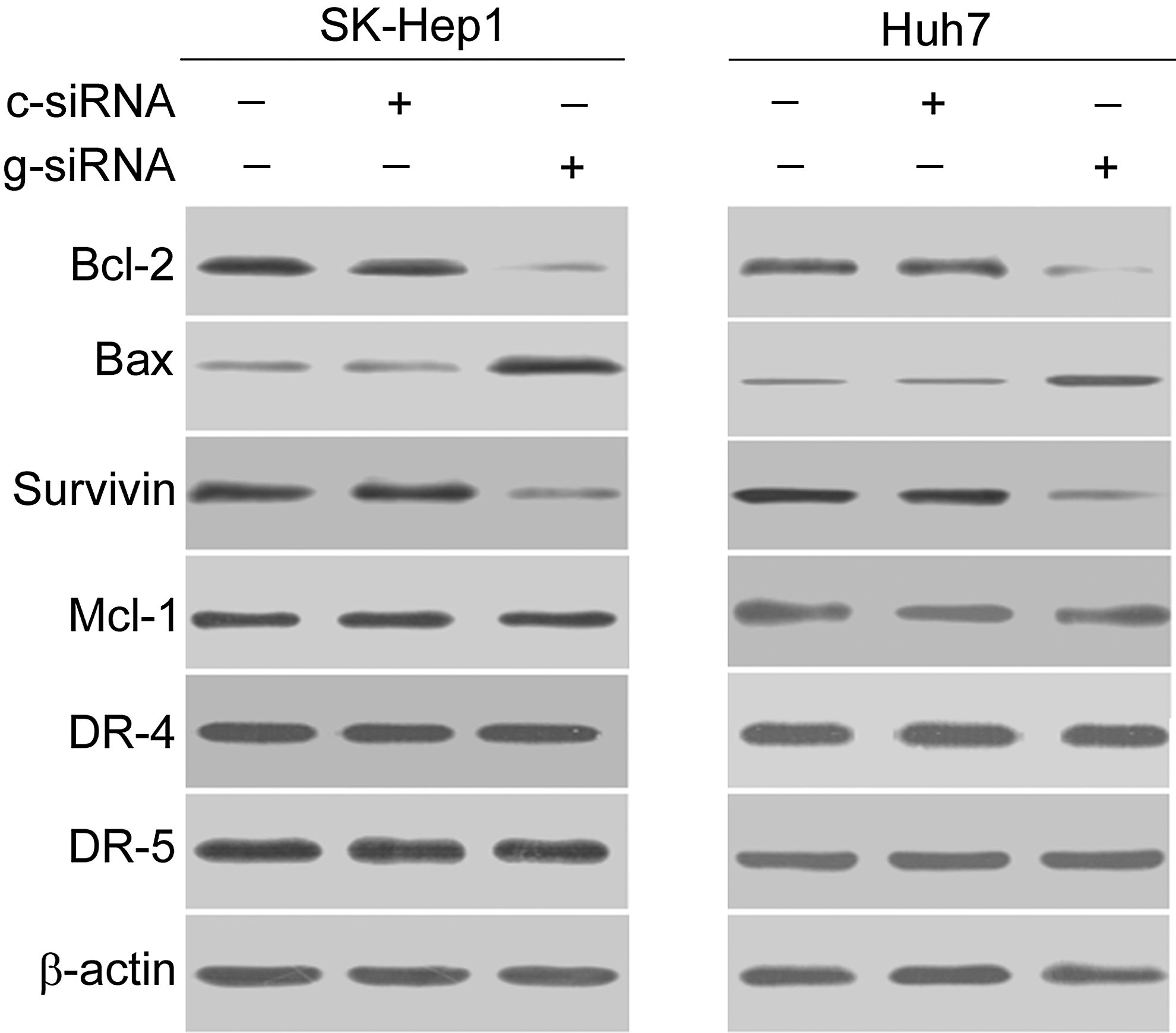
Western blot analysis of indicated proteins in SK-Hep1 and Huh7 cells transfected with control siRNA (c-siRNA) or galectin-1 siRNA (g-siRNA). Representative blots from three independent experiments are shown.
Overexpression of Bcl-2 and survivin counteracts galectin-1 downregulation-induced apoptosis
Next, the authors explored the possibility if enforced expression of Bcl-2 and survivin prevents HCC cells from galectin-1 depletion-induced apoptosis. As shown in Figure 3A, cotransfection with Bcl-2- and survivin-expressing plasmids restored the expression of Bcl-2 and survivin in galectin-1 siRNA-transfected cells. Restoration of Bcl-2 or survivin alone slightly reduced the apoptosis of HCC cells induced by galectin-1 silencing (Fig. 3B). Most interestingly, overexpression of both Bcl-2 and survivin almost completely blocked galectin-1 downregulation-induced apoptosis.
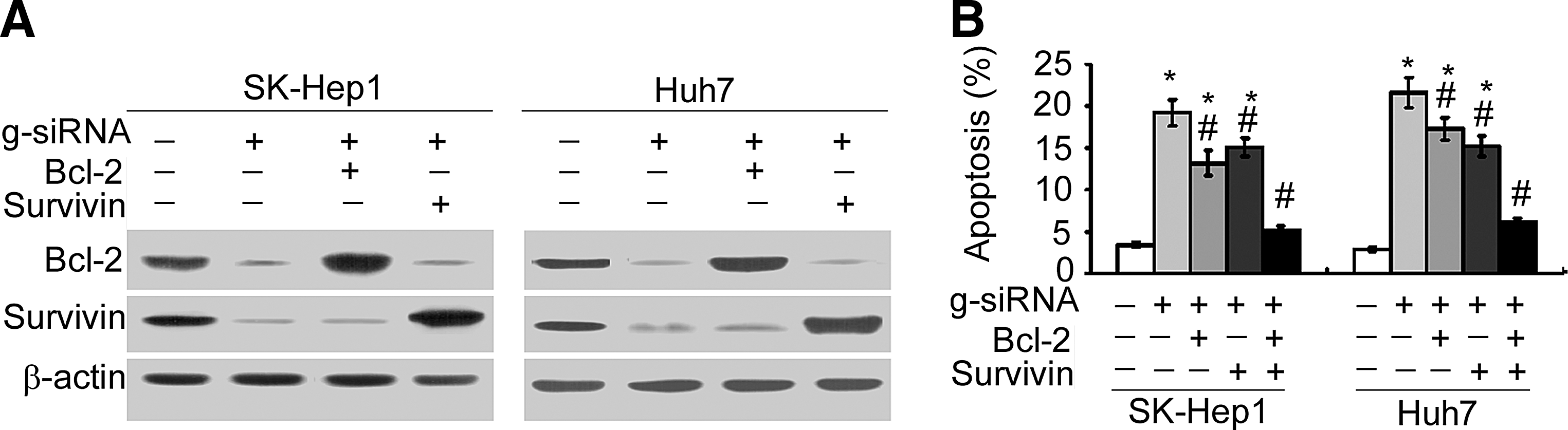
Overexpression of Bcl-2 and survivin counteracts galectin-1 downregulation-induced apoptosis.
Downregulation of galectin-1 enhances TRAIL cytotoxicity against HCC cells
Next, the authors examined the effect of galectin-1 downregulation on the susceptibility of HCC cells to TRAIL. The MTT assay demonstrated that exposure to 100 ng/mL TRAIL for 24 h caused 11%–14% reduction in the viability of SK-Hep1 and Huh7 cells, compared to untreated cells (Fig. 4A). Galectin-1 silencing exerted a marginal, but not significant, effect on the viability of HCC cells studied (p > 0.05 vs. nontransfected cells; Fig. 4A). Notably, the delivery of galectin-1 siRNA, but not control siRNA, significantly (p < 0.05) augmented TRAIL cytotoxicity against HCC cells. Moreover, apoptosis analysis indicated that silencing of galectin-1 potently enhanced the apoptosis of SK-Hep1 and Huh7 cells in the presence of TRAIL (Fig. 4B).
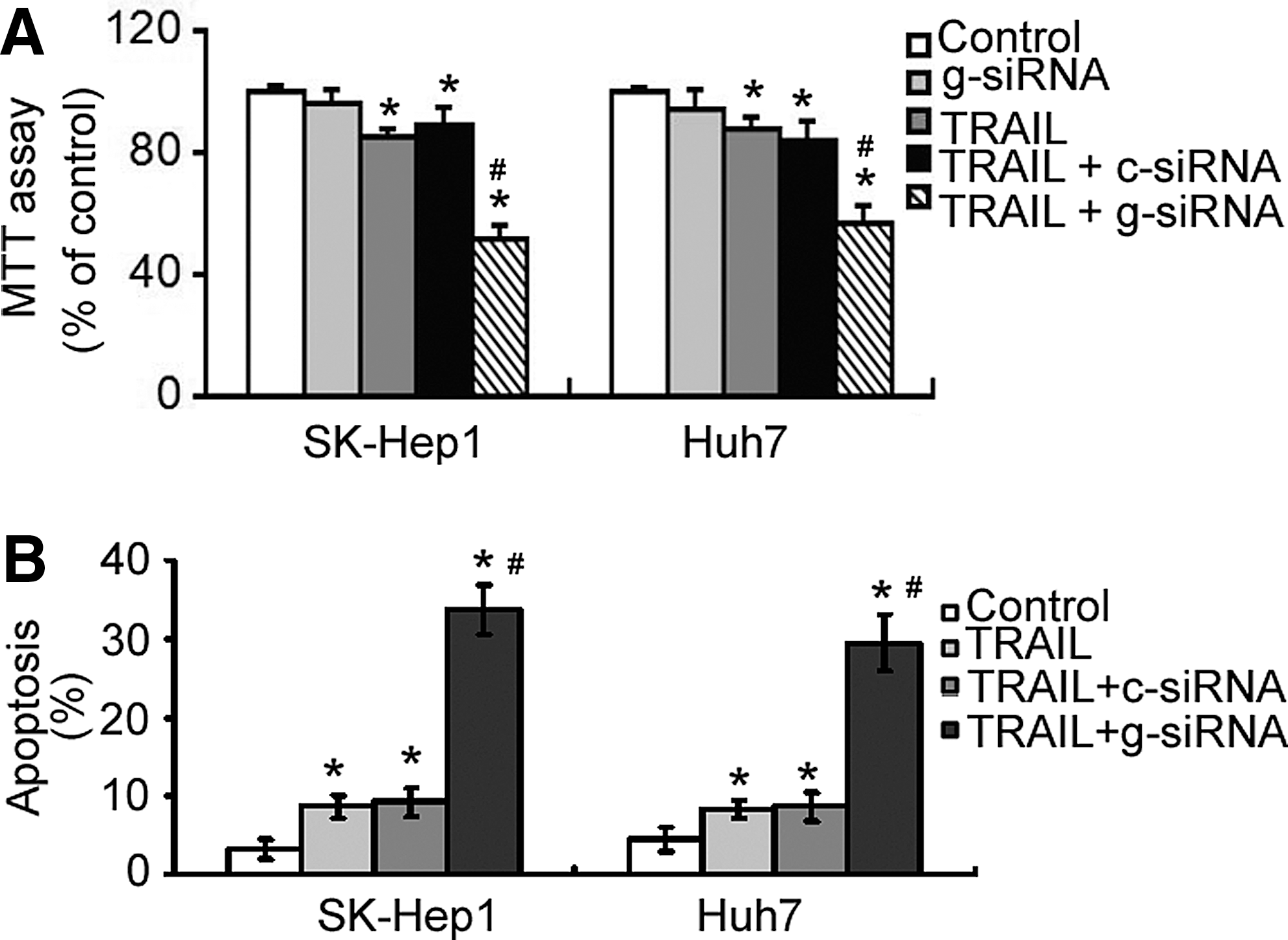
Downregulation of galectin-1 enhances tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) cytotoxicity against HCC cells. Cells were transfected with control siRNA (c-siRNA) or galectin-1 siRNA (g-siRNA) and incubated for 48 h before exposure to 100 ng/mL TRAIL for additional 24 h.
Galectin-1 silencing-induced TRAIL sensitization involves concomitant downregulation of Bcl-2 and survivin
Finally, the authors checked whether galectin-1 silencing-induced TRAIL sensitization was mediated through alteration of Bcl-2 and survivin proteins. Western blot analysis confirmed that galectin-1 knockdown resulted in a marked reduction in the protein levels of Bcl-2 and survivin in the presence of 100 ng/mL TRAIL (Fig. 5A). Overexpression of Bcl-2 or survivin partially prevented galectin-1-silenced HCC cells from apoptosis in the presence of TRAIL (Fig. 5B). Notably, enforced expression of both Bcl-2 and survivin almost completely abrogated the apoptosis of galectin-1-silenced, TRAIL-treated HCC cells.
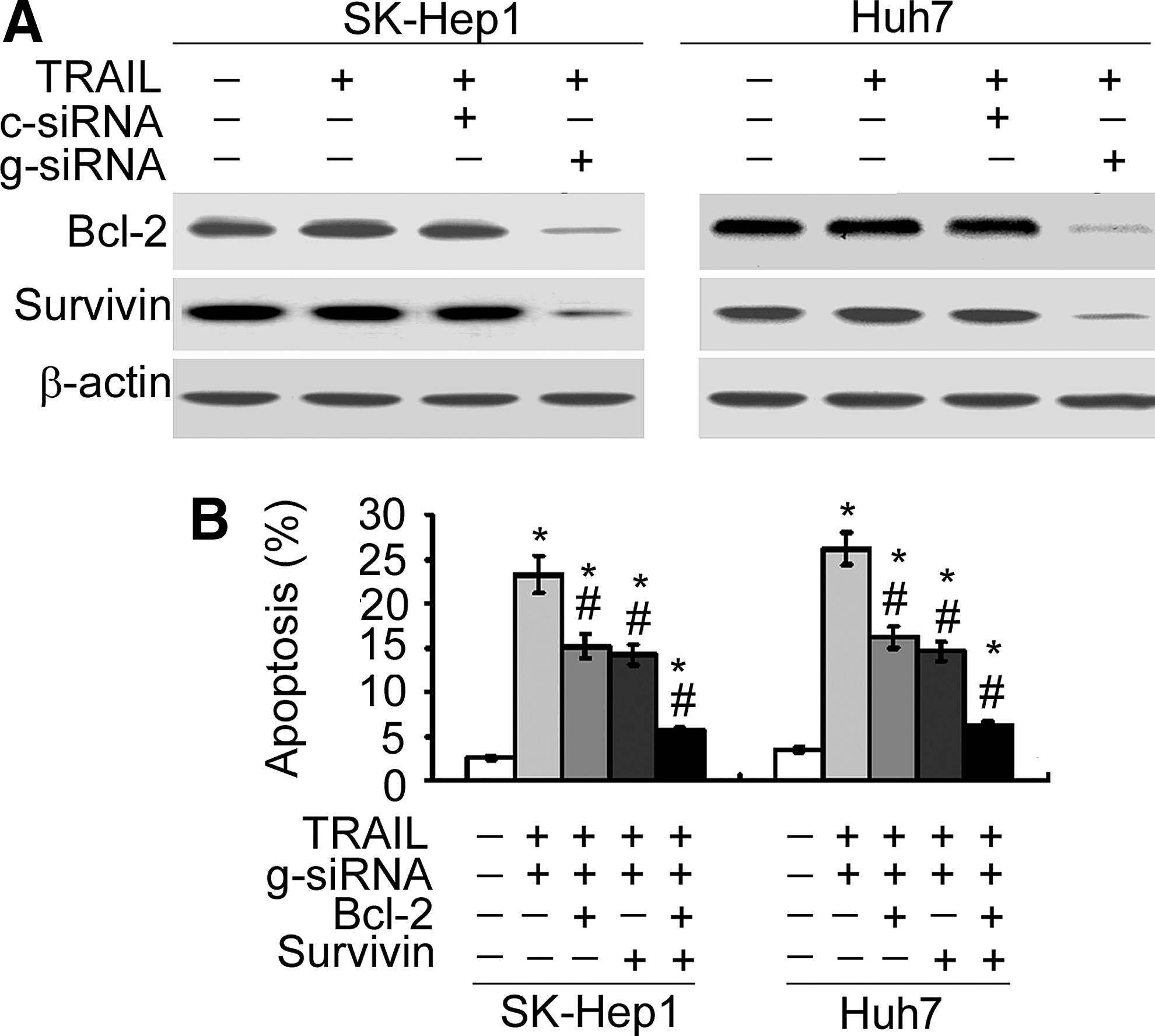
Overexpression of Bcl-2 and survivin blocks the apoptosis of galectin-1-silenced, TRAIL-treated HCC cells.
Discussion
Galectin-1 is implicated in multiple tumor progression steps, including cancer cell proliferation, adhesion, apoptosis, invasion, and tumor angiogenesis. 21 It has been documented that OTX008, a selective small-molecule inhibitor of galectin-1, can inhibit the proliferation of a panel of cancer cells. 21 Small-hairpin RNA-mediated knockdown of galectin-1 has been shown to suppress the growth and invasion of osteosarcoma cells. 22 The involvement of galectin-1 in HCC growth and progression has been described in several previous studies. 14,15,23 It has been reported that galectin-1 overexpression promotes HCC cell migration and invasion. 23 Consistently, these data confirmed the importance of galectin-1 in HCC cell survival. The authors found that targeted reduction of galectin-1 caused significant apoptosis in SK-Hep1 and Huh7 cells. Despite these favorable effects on tumor progression, galectin-1 may induce negative effects on some types of cancer cells. For instance, restoration of galectin-1 expression in human colorectal cancer cells induces apoptotic death. 24 Similarly, galectin-1 can trigger apoptosis in prostate cancer 25 and breast cancer 26 cells. These findings suggest that the biological roles of galectin-1 are cellular context dependent.
In HCC cells, galectin-1 overexpression has been found to modulate multiple molecular pathways involved in tumor progression, including the mitogen-activated protein kinase (MAPK), phosphoinositide 3-kinase (PI3-K)/Akt, and Wnt/β-catenin pathways. 14,15 These signaling pathways are usually involved in HCC cell survival through alteration of apoptosis regulators. 27,28 It has been documented that galectin-1 overexpression promotes tumorigenesis and cisplatin resistance in epithelial ovarian cancer through activation of ERK MAPK signaling and upregulation of Bcl-2. 12 These data further supported the regulation of the Bcl-2 family proteins by galectin-1. The authors found that galectin-1 downregulation resulted in a sharp decline in the Bcl-2 and survivin protein and elevation in the Bax protein. Bax translocalizes to the mitochondrial outer membrane, in response to proapoptotic stress signals, consequently inducing cytochrome c release from the mitochondria and activating the caspase signal pathway. 29 The antiapoptotic protein such as Bcl-2 can counteract BAX-mediated cell death. These data showed that the enforced expression of Bcl-2 and survivin almost completely blocked the induction of apoptosis by galectin-1 silencing. These data collectively indicate that suppression of Bcl-2 and survivin expression largely accounts for the proapoptotic effect of galectin-1 silencing in HCC cells. However, the signaling pathway(s) involved in galectin-1-mediated regulation of Bcl-2 and survivin remains to be further defined.
Alteration of apoptosis regulators, especially the Bcl-2 family, has been linked to affect the susceptibility of cancer cells to TRAIL. 30 Wang et al. 31 reported that ABT-263, a potent inhibitor of the Bcl-2 family, has shown the ability to sensitize HCC cells to TRAIL-induced apoptosis. Chen et al. 32 demonstrated that silencing of cyclooxygenase-2 can enhance the sensitivity of HCC cells to TRAIL through inhibition of the antiapoptotic proteins, Bcl-2 and Bcl-w. Given the regulation of apoptosis regulators by galectin-1, the authors next checked the effect of galectin-1 silencing on TRAIL sensitivity of HCC cells. The data revealed that depletion of galectin-1 significantly augmented the apoptosis of TRAIL-treated HCC cells. However, galectin-1 silencing had no impact on the expression of DR4 and DR5, suggesting that the action of galectin-1 may be linked to alteration of the apoptotic pathways downstream, DR4 and DR5. Most interestingly, overexpression of Bcl-2 and survivin significantly blocked the apoptosis of galectin-1-silenced, TRAIL-treated HCC cells.
Taken together, these data suggest that galectin-1 is implicated in the modulation of TRAIL sensitivity of HCC cells through alteration of Bcl-2 and survivin.
To the best of knowledge, this is the first report showing the importance of galectin-1 in the regulation of TRAIL sensitivity of HCC cells. The authors show that targeted reduction of galectin-1 induces apoptosis and enhances TRAIL cytotoxicity in HCC cells, which is largely mediated through downregulation of Bcl-2 and survivin. Therefore, galectin-1 may represent a promising target for improving the efficacy of TRAIL-based therapy against HCC.
Footnotes
Disclosure Statement
No competing financial interests exist.