
Abstract
Taxol is a cytotoxic antiepithelioma chemotherapy drug widely used clinically, which results in appearing a broad range of taxol-resistant tumors. Oncoprotein 18 (Op18)/stathmin is a genetically highly conserved small-molecule cytosolic phosphoprotein and highly expressed in tumors. Extracellular signal-regulated kinase (ERK) is a main member of mitogen-activated protein kinases (MAPKs). The study demonstrated that combination of blockage of ERK signal by ERK inhibitor PD98059 and Taxol greatly promoted taxol-induced cellular apoptosis and growth inhibition, decreased the expression of Op18/stathmin and total levels of phosphor-Op18/stathmin, while weakened the cyclin-dependent kinase 2 (cdc2) activity and antiapoptotic protein Bcl-2 expression and inhibited IL-10 autocrine in taxol-resistant NCI-H1299 cells; Taxol-resistant NCI-H1299 cells expressed high levels of ERK and phosphor-ERK in contrast to taxol-sensitive CNE1 cells, and ERK mainly phosphorylated Op18/stathmin at Ser 25 site. These findings suggest that ERK-mediated Op18/stathmin is involved in taxol resistance of tumors; blockage of ERK signal improves the sensitivity of tumor cells to taxol, which provides new clues for treating taxol-resistant carcinomas.
Introduction
Oncoprotein 18 (Op18)/stathmin, a 18–19 kDa relative molecular mass and genetically highly conserved small-molecule cytosolic phosphoprotein, is expressed at high levels in carcinoma cells. 1 It directly regulates microtubule dynamics equilibrium through phosphorylated inactivation and dephosphorylated activation, which mediate multiple cellular biological behaviors and even cellular death. 2 –4 Op18/stathmin has four phosphorylation sites, Ser16, Ser25, Ser38, and Ser63, and is a downstream molecular target of multiple signaling pathways. It is regulated by multiple kinases and phosphatases, such as mitogen-activated protein kinases (MAPKs), cyclin-dependent kinase 25 (cdc25), and protein phosphatase 2A (PP2A). They are vital to maintaining a malignant phenotype of tumor cells. 1,5 –8
The MAPK family is a series of typical Ser/Thr kinases, which has three subfamilies, and the extracellular signal-regulated kinase (ERK) is a main member of it. The outcome of ERK activation ranges from cellular survival to proliferation and differentiation, cellular senescence, and apoptosis as well. 9 –13 The earlier study discovered that Epstein–Barr virus-encoded LMP1 triggered regulation of the ERK-mediated Op18/stathmin signaling in nasopharyngeal carcinoma cells; Op18/stathmin was a downstream target of ERK, which was in association with the acceleration of cell cycle progression. 14
In the latest study, five different epithelial-derived carcinoma cell lines, including human nasopharyngeal carcinoma cell line CNE1, human gastric cancer cell line MGC, human breast cancer cell line MCF-7, human hepatocellular carcinoma cell line Hep3B-2, and human nonsmall cell lung cancer cell line NCI-H1299, were randomly used to screen taxol-resistant cells, which demonstrated that NCI-H1299 cells of these five cell lines were evidently taxol resistant; inhibition of Op18/stathmin expression promoted the sensitivity of NCI-H1299 cells to taxol. 1
This study mainly focuses on whether ERK-mediated Op18/stathmin signaling is involved in the resistance of NCI-H1299 cells to taxol and explored the possible molecular mechanisms of taxol resistance development.
Materials and Methods
Cell lines and culture
Both Taxol-sensitive human nasopharyngeal carcinoma CNE1 cells and taxol-resistant human nonsmall cell lung cancer NCI-H1299 cells were used in this study. 1 They were grown in RPMI 1640 (Gibco; BRL) supplemented with 10% fetal bovine serum (HyClone), 100 IU/mL penicillin, and 100 ug/mL streptomycin at 37°C in a humidified atmosphere of 5% CO2.
Antibodies and chemical reagents
Primary antibodies were the rabbit polyclonal antistathmin (Calbiochem), mouse monoclonal anti-β-actin (Sigma), rabbit anti-phosphoserine-IgG (Zymed), rabbit polyclonal anti-ERK1 (C-16) and mouse monoclonal antiphospho-erk1 IgG (sc-7383; Santa Cruz), mouse monoclonal anticaspase-3 (Santa Cruz), mouse monoclonal anticaspase-8 (Cell Signaling), rabbit monoclonal anticaspase-9 (Cell Signaling), rabbit polyclonal antistathmin (phospho S25, S38; Abcam), rabbit monoclonal antistathmin (phospho S63; Abcam), rabbit polyclonal anti-cdc2 (Santa Cruz), rabbit polyclonal antiphospho-Thr161-cdc2 (Cell Signaling), and rabbit polyclonal anti-Bcl-2 (Santa Cruz). Secondary antibodies were horseradish peroxidase (HRP)-conjugated goat-antirabbit IgG and HRP-conjugated rabbit-antimouse IgG (Santa Cruz). Taxol was purchased from Santa Cruz (sc-201439), and PD98059 (Cat. No. 9900), a specific ERK inhibitor, was obtained from Cell Signaling Technology; either was dissolved in dimethyl sulfoxide (DMSO) at appropriate concentrations and stored at −20°C. Human interleukin10 ELISA kit was from R&D.
Cellular apoptosis assays
Cells were seeded at 5 × 105/mL and allowed to grow overnight. PD98059 (0, 5, 10 μM) was added for 2 hours when cells reached 80% confluency, DMSO was used as the control, cells were treated by taxol (100 nM) for 12 hours, washed with cold phosphate buffer solution (PBS), digested with 0.25% trypsin for 3–5 minutes, and then centrifuged at 800 rpm for 5 minutes, resuspended with PBS, and centrifuged and washed three times repeatedly, the supernatant was removed and cells were fixed with 70% ethanol, and cellular apoptosis assays were performed in triplicate by the fluorescence-activated cell sorter (FACS) through specialized company (Beijing Dingguo).
Preparation of cell lysates and western blot analysis
Cells were harvested and washed with PBS and lysed with the cell lysis buffer (50 mM Tris-HCl pH 8.0, 1 mM EDTA, 2% sodium dodecyl sulfate [SDS], 5 mM DTT, 10 mM PMSF) on ice for 30 minutes and then centrifuged at 15,000 g for 10 minutes after sonication for 30 seconds. The supernatant was collected as whole-cell lysates for western blot analysis. Protein concentration was determined by the BCA Protein Assay Reagent (Pierce).
Protein extracts (50 μg/sample) were separated by 10% (or 12%) SDS-polyacrylamide gel electrophoresis and then electrotransferred onto a nitrocellulose membrane. The membranes were blocked with a buffer containing 5% nonfat milk in PBS with 0.05% Tween-20 for 2 hours and incubated with primary antibodies overnight at 4°C. After the second wash, the membranes were incubated with HRP-conjugated secondary antibody for 1 hour at room temperature and developed with an enhanced chemiluminescence detection kit (Pierce).
Immunoprecipitation-western blot
Because no specific antiphosphor-stathmin antibody was available to assay the phosphorylation of Op18/stathmin, cell extracts were prepared with harvested cells lysed in an immunoprecipitation (IP) lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 10% NP-40, 1 mM EDTA, 10% glycerol, 10 mM NaF, 1 mM Na3VO4, 1 mM DTT, 1 mM PMSF, and protease inhibitor cocktail tablet). The supernatant obtained was treated as previously described. 1 Immunoprecipitates were collected and washed five times, then 500 μg was subjected to western blot analysis.
Assays of autocrine IL-10 by tumor cells
Cells were cultured in freshly complete medium without a pH indicator; when reached 80% confluence, cells were treated by PD98059 (0, 5, 10 μM) for 2 hours, then supplemented 100 nM taxol; 12 hours later, the media were collected and centrifugated at 3,000 rpm for 30 minutes; the supernatants were also separated and stored at −20°C for autocrine IL-10 assays referring to the instruction of Human Interleukin10 ELISA Kit (R&D). The values of optical density (OD) were measured at 450 nm wavelength. All experiments were carried out thrice repeatedly.
Statistics
All statistical calculations were performed using the statistical software SPSS17.0. Differences with p-values <0.05 were considered statistically significant. Data are expressed as mean ± SD.
Results
Blockage of ERK signal evidently promotes taxol-induced cellular apoptosis and growth inhibition in NCI-H1299 cells
NCI-H1299 cells were treated with gradient concentrations of ERK inhibitor PD98059 (0, 5, 10 μM) together with stable 100 nM taxol; the morphological photographs of cell growth showed that the degrees of cellular confluence were decreased, cells became sparse and round, appeared as a large number of translucent suspension cells with the increase of PD98059. FACS analysis showed that cellular apoptosis rates were 8.98%, 19.53%, and 21.18% for gradient concentrations of 0, 5, and 10 μM PD98059 and stable taxol, respectively; PD98059 greatly promoted taxol-induced cellular apoptosis in NCI-H1299 cells (Fig. 1A). Figures also demonstrated that cells presented no obvious changes in shape and density for single PD98059 treatment at gradient concentrations, similar to the control group treated with the solvent DMSO. While cellular apoptosis rates were only slightly leveled up for single PD98059 treatment of 0, 5, and 10 μM, separately were 0.12%, 3.99%, and 4.65%. ERK signal inhibition alone was insufficient to induce significant cellular apoptosis in NCI-H1299 cells (Fig. 1B).
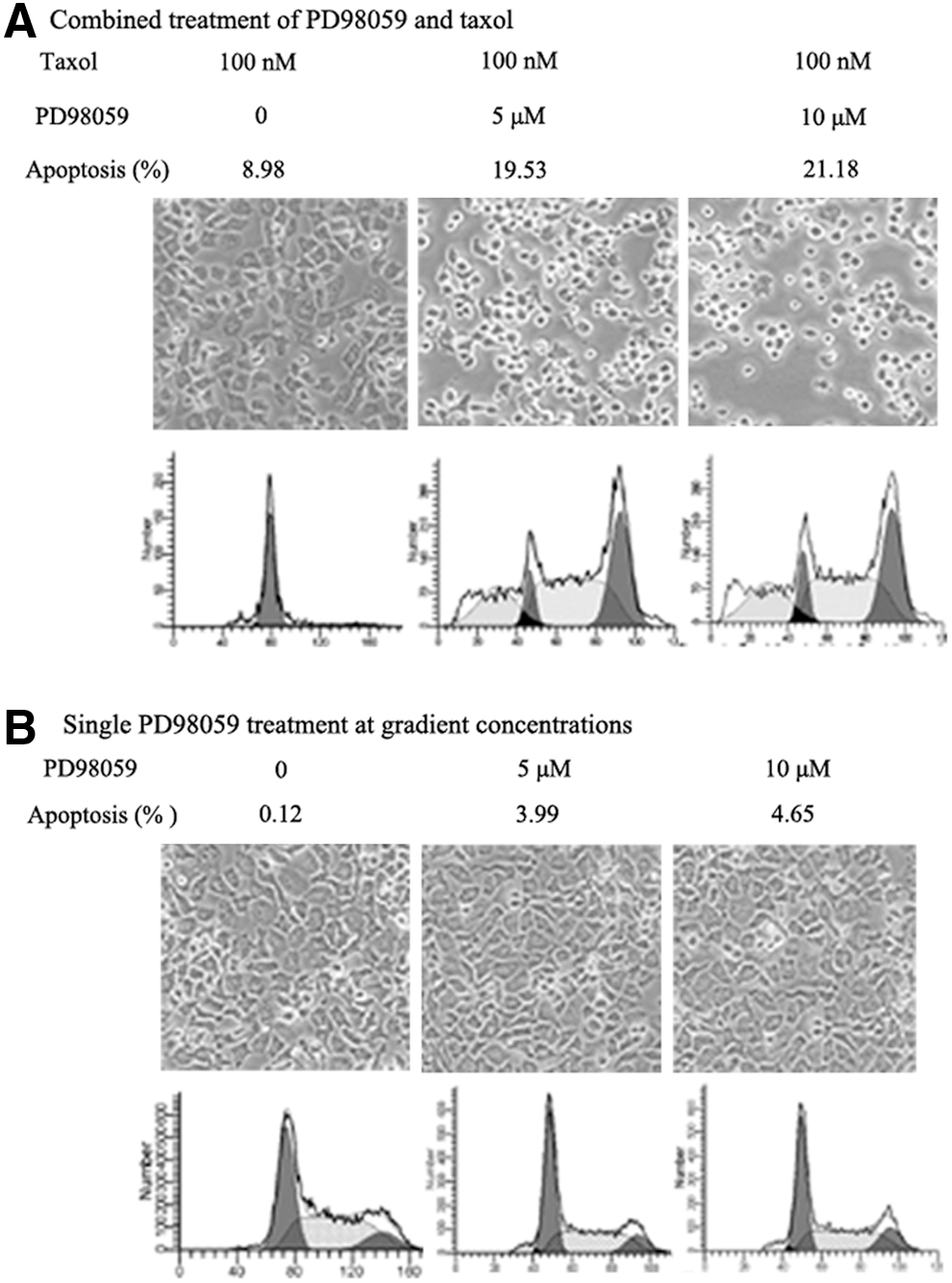
Blockage of ERK signal by PD98059 evidently promotes taxol-induced cellular apoptosis and growth inhibition in NCI-H1299 cells.
Combined treatment of PD98059 and taxol effectively increases the expressions of caspase-3 and caspase-9
Caspases were analyzed by western blot in NCI-H1299 cells treated by a combination of PD98059 and taxol; the cleavages of caspase-3 and caspase-9 were clearly increased with the increase of PD98059 in NCI-H1299 cells, but no obvious changes were observed in caspase-8 among three groups for combined treatment of PD98059 and taxol; caspase-3, caspase-8, and caspase-9 were not markedly affected in single taxol treatment group related to controls of the blank and the DMSO; similarly, no obvious changes were presented for solo PD98059 treatment at gradient concentrations (Fig. 2).
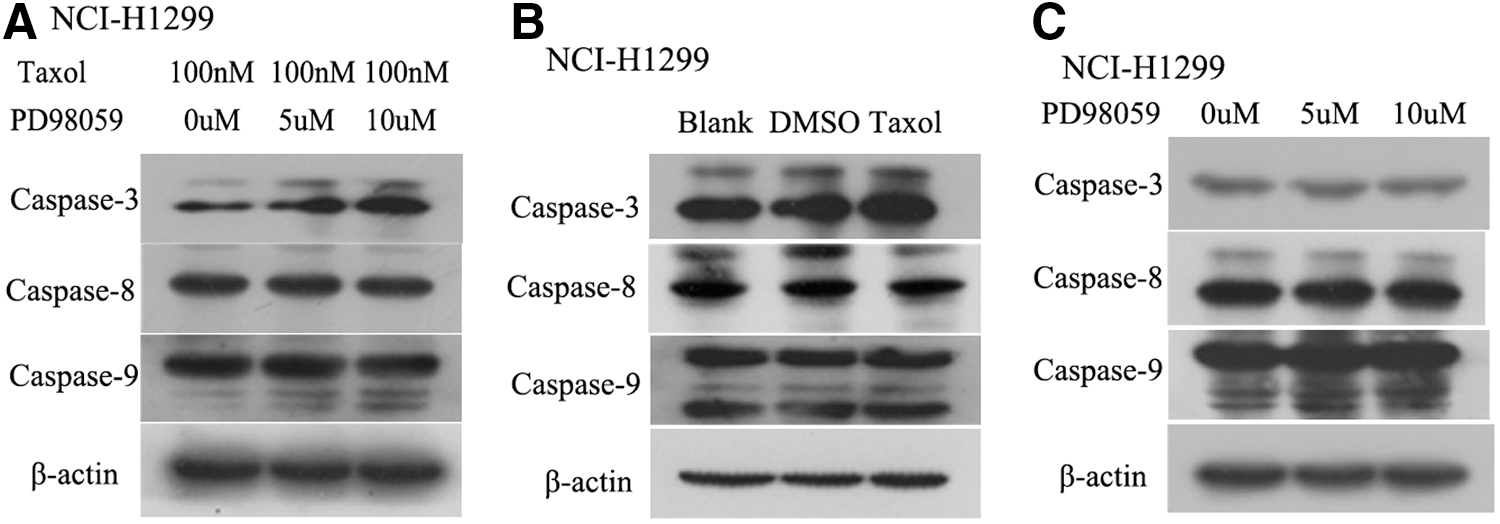
The expressions of a series of caspases were analyzed by western blot.
PD98059 evidently promotes taxol-induced cellular apoptosis through upregulating the expressions of caspase-3 and caspase-9 in NCI-H1299 cells.
Combination of PD98059 and taxol downregulates the expression of Op18/stathmin in NCI-H1299 cells
Former studies indicated that Op18/stathmin was closely associated with taxol resistance of cells and is also a downstream target of ERK. To explore whether the Op18/stathmin signaling regulation mediated by ERK is involved in taxol-resistant development in NCI-H1299 cells, the change of Op18/stathmin expression was analyzed for the combined treatment of blocking ERK signal and taxol.
Western blot analysis showed that Op18/stathmin expression was decreased with dose increases of PD98059 in taxol-treated NCI-H1299 cells; PD98059 augmented inhibition of Op18/stathmin expression induced by taxol (Fig. 3A). Similarly, the combination of fixed PD98059 and gradient taxol (0, 10, and 100 nM) markedly inhibited Op18/stathmin expression in the taxol dose-dependent pattern (Fig. 3B). The authors still find that single treatment of either taxol or PD98059 at gradient concentrations caused downregulation of Op18/stathmin expression to a certain extent (Fig. 3C, D). PD98059 augments taxol-induced inhibition of Op18/stathmin expression in synergy in NCI-H1299 cells.
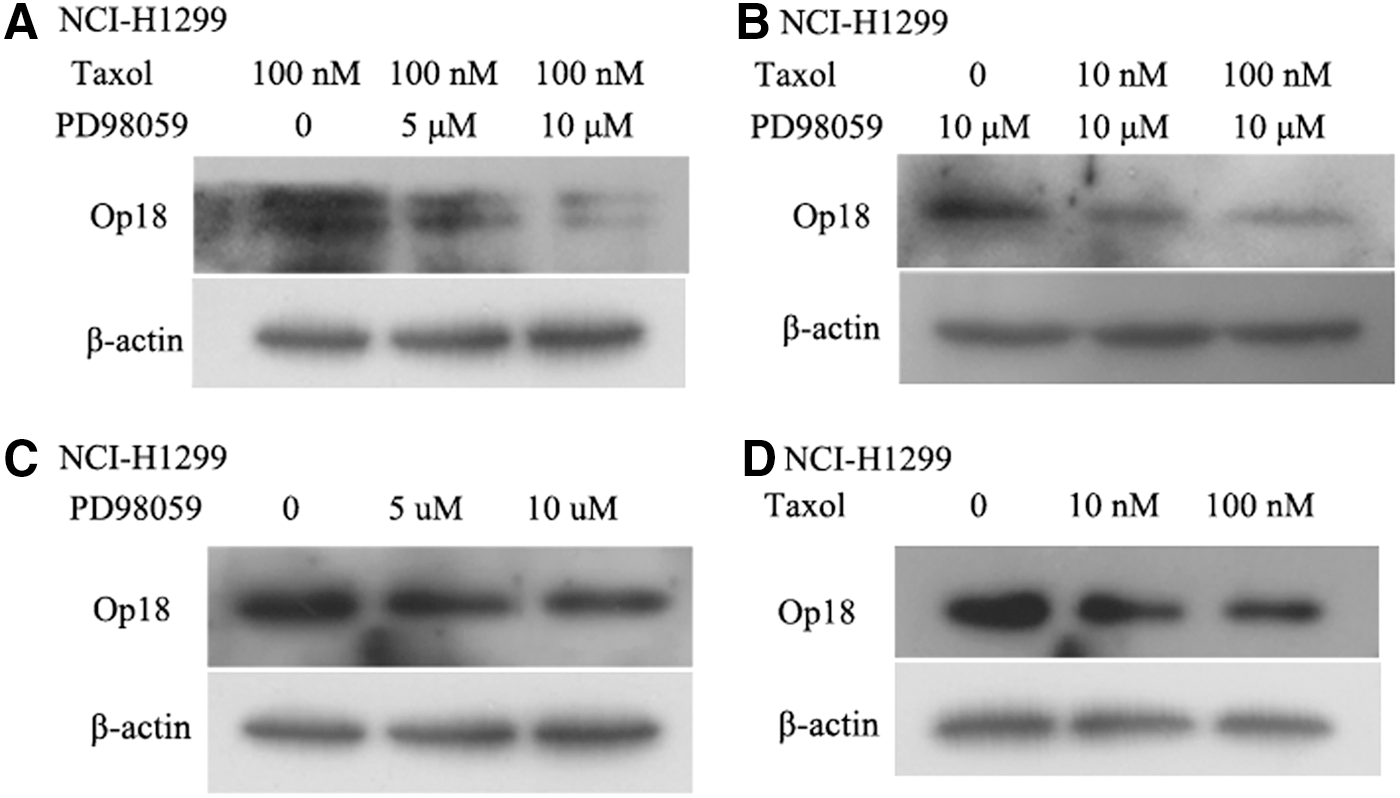
Combination of PD98059 and taxol cooperatively downregulates the expression of Op18/stathmin in NCI-H1299 cells.
PD98059 and taxol cooperatively inhibit the phosphorylation of Op18/stathmin; the main change of phosphorylated sites is ser25 for blocking ERK signal
The total levels of phosphorylated Op18/stathmin were analyzed by IP-western blot; briefly, Op18/stathmin was pulled down with the antistathmin antibody, then the antiphosphor-serine antibody was applied to detect the total levels of phosphor-Op18/stathmin.
The study found the combined treatment of PD98059 and taxol reduced the total levels of phosphor-Op18/stathmin in PD98059 dose-dependent manner (Fig. 4A). Furthermore, the main phosphorylated sites of Op18/stathmin mediated by ERK were analyzed by western blot; there were no obvious changes at phosphorylated sites of ser16, ser38, and ser63 of Op18/stathmin for blocking ERK signal, but PD98059 markedly inhibited the phosphorylation of Op18/stathmin at ser25 site with a dose increase, which indicated that blockage of ERK signal would promote ser25 dephosphorylation of Op18/stathmin, in other words, ERK mainly phosphorylated Op18/stathmin at ser25 site (Fig. 4B).
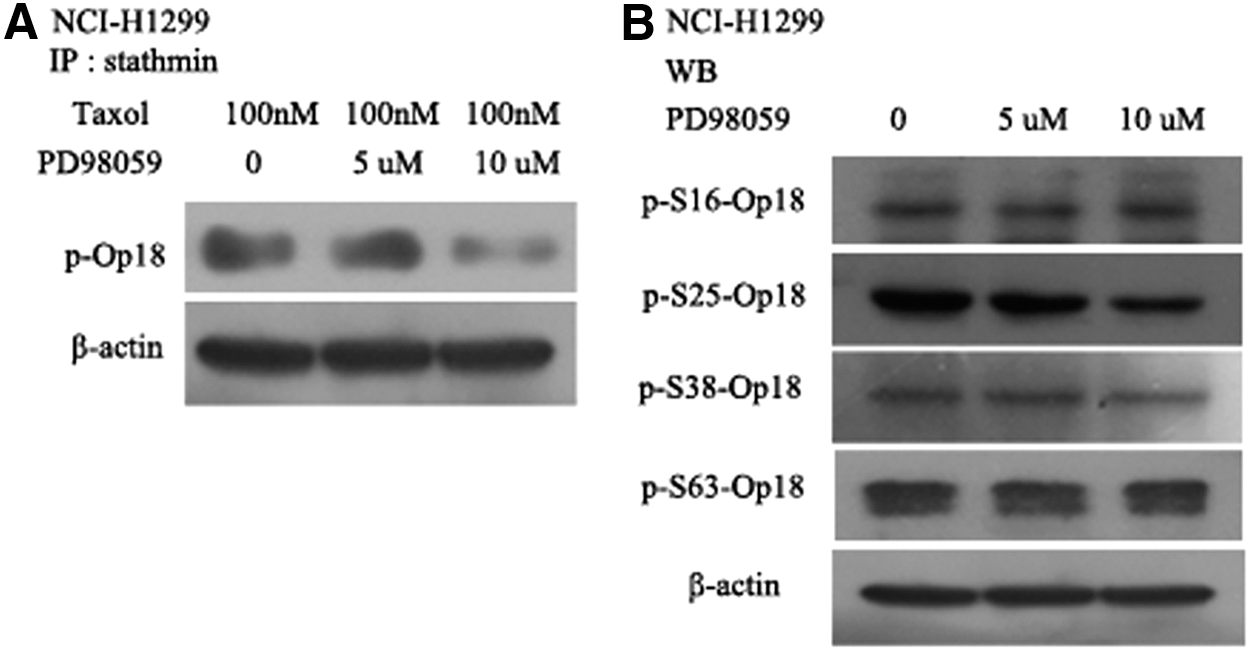
PD98059 and taxol cooperatively inhibited phosphorylation of Op18/stathmin.
PD98059 promotes downregulation of the expressions of p-thr161-cdc2 and Bcl-2 in NCI-H1299 cells
Previous studies certified that cdc2 was also an upstream kinase of Op18/stathmin signaling, involving the acceleration of cell cycle progression; cdc2 activation depended on the phosphorylation of its thr161 site. 15 Bcl-2 is a typical antiapoptosis protein in tumor cells. The study showed that PD98059 did not alter the expression of cdc2, but downregulated the phosphorylation of its thr161 site in a dose-dependent manner in taxol-treated NCI-H1299 cells. Meanwhile, PD98059 promoted inhibition of Bcl-2 expression; the inhibitive effects of 10 μM PD98059 is the most remarkable among three groups treated by constant taxol (Fig. 5A). In contrast to combined treatment, single taxol did not affect the expressions of cdc2 and Bcl-2, but downregulated the phosphorylation of cdc2 thr161 site (Fig. 5B); gradient PD98059 treatment merely declined Bcl-2 expression, but did not change the expressions of cdc2 and p-thr161-cdc2 (Fig. 5C).

PD98059 promotes inhibition of the expressions of p-thr161-cdc2 and bcl-2 in taxol-treated NCI-H1299 cells, but does not alter cdc2 expression.
PD98059 enhances inhibition of IL-10 autocrine by taxol-treated NCI-H1299 cells
Collected the supernatants of cell culture for IL-10 detection by the enzyme-linked immunosorbent assay (ELISA); the mean of OD values represented the level of autocrine IL-10 from tumor cells treated. The study showed that the average OD value was 0.331 in the single taxol group, while the value of the group of 5 μM PD98059 together with taxol was 0.271, the one of 10 μM PD98059 and taxol was 0.163.
The histograms demonstrated that the levels of autocrine IL-10 in supernatants by tumor cells were in turn decreased with PD98059 augment; the difference was significant between the combined group of 10 μM PD98059 and taxol and the other two groups (p < 0.05) (Fig. 6).
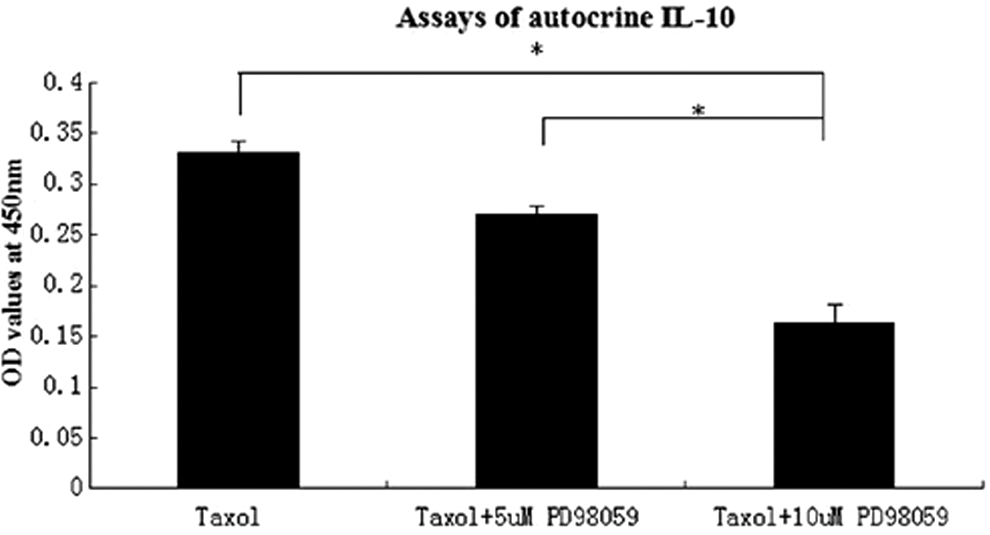
PD98059 enhances inhibition of IL-10 autocrined by NCI-H1299 cells treated by taxol in a dose-dependent manner. *p < 0.05.
Taxol itself also inhibits the expressions of Op18/stathmin and ERK and phosphor-ERK to a certain extent
Western blot analysis certified that taxol itself also decreased Op18/stathmin expression and similarly inhibited ERK expression and its phosphorylation to some extent in NCI-H1299 cells in contrast to two controls of the Blank and the DMSO (Fig. 7A).

Taxol itself inhibits the expressions of Op18/stathmin, ERK, and phosphor-ERK to a certain extent in NCI-H1299 cells.
In NCI-H1299 cells treated with gradient taxol at 0, 10, and 100 nM, the expressions of Op18/stathmin and ERK and phosphor-ERK were markedly decreased with taxol increase; treatment of 100 nM taxol was the most notable among the three groups (Fig. 7B).
Both ERK and phosphor-ERK are highly expressed in NCI-H1299 cells in contrast to taxol-sensitive CNE1 cells
The earlier study certified that human nasopharyngeal carcinoma CNE1 cells were taxol sensitive, but human nonsmall cell lung cancer NCI-H1299 cells displayed evident resistance to taxol, both were screened from five different epithelial-derived carcinoma cell lines randomly used; CNE1 cells were completely different from NCI-H1299 cells in taxol tolerance. 1 Both cell lines were used to investigate the possible relationship of ERK signal and taxol resistance.
Western blot analysis showed Op18/stathmin expression was at a high level in taxol-resistant NCI-H1299 cells in contrast to taxol-sensitive CNE1 cells, which were in agreement with the previous study 1 ; similarly, the expressions of ERK and phosphor-ERK were significantly higher in NCI-H1299 cells than in CNE1 cells, all these might imply that ERK activity is positive relevant to taxol resistance of tumor cells (Fig. 8).
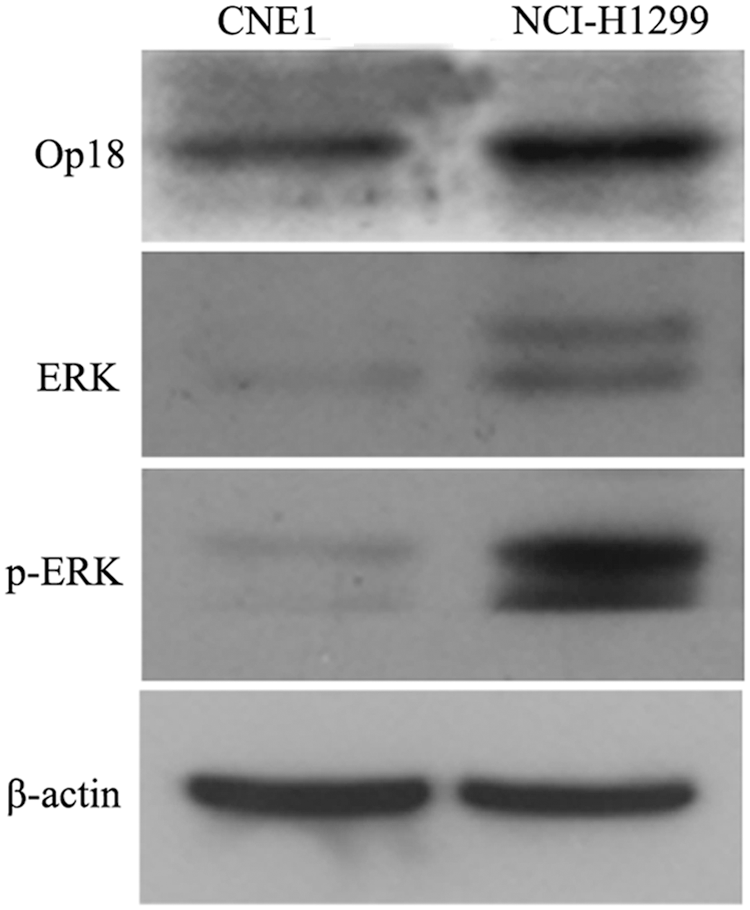
Western blot analysis displays bands of Op18/stathmin, ERK, and phosphor-ERK in both CNE1 cells and NCI-H1299 cells. High level of Op18/stathmin was presented in taxol-resistant NCI-H1299 cells in contrast to taxol-sensitive CNE1 cells; similarly, the expressions of ERK and phosphor-ERK were significantly higher in NCI-H1299 cells than in CNE1 cells.
Discussion
Taxol (namely paclitaxel) is the only antitumor's drug in clinical use identified by the plant screening program, and a microtubule-stabilizing drug approved by the Food and Drug Administration of America for the treatment of ovarian, breast, and lung cancer, as well as Kaposi's sarcoma and so on; taxol has been recognized through inducing microtubule polymerization to cause cell death. 16
Some serious side-effects, such as hypersensitivity reactions, myelosuppression, and neurotoxicity, are closely associated with taxol, which limits the dose increment of taxol for treating taxol-resistant tumors. 17 –19 To increase taxol sensitivity and decrease its side-effects, combination chemotherapies with gemcitabine–paclitaxel, paclitaxel–cyclophosphamide, paclitaxel–cisplatin, and paclitaxel–epirubicin were applied in urothelial carcinoma cells, advanced nonsmall cell lung cancer, and metastatic pancreatic adenocarcinoma, which independently and significantly improved overall survival, but risk a multidrug-resistant development of tumors. 20 –22
Op18/stathmin is closely relevant to maintaining a tumor malignant phenotype, and an anticancer therapeutic target; the latest study confirmed that high expression of Op18/Stathmin was closely associated with the development of taxol resistance. 1 People have begun to explore the possibility of treating drug-resistant tumors through targeting Op18/Stathmin signaling bound traditional chemotherapeutics.
This study showed that the combined treatment of blockage of ERK signal by PD98059 and taxol suppressed cellular proliferation and promoted cellular apoptosis. Combination of PD98059 and taxol started the expressions of caspase-3 and caspase-9. It is well known that caspase-3 is the executor of cellular apoptosis, caspase-8 is an initiator of extrinsic cellular apoptosis, and caspase-9 is a promoter of endogenous cellular apopotosis, 23 which implied an endogenous cellular apoptosis pathway activation in NCI-H1299 cells for combined treatment. The combination obviously inhibited Op18/stathmin expression related to single treatment of taxol or PD98059. High level of Op18/stathmin is thought to be closely associated with a poor response to containing taxol chemotherapy in endometrial carcinoma and other epithelial-derived carcinomas. 1,17
The authors also find that combined treatment would decrease the total level of phosphor-Op18/stathmin; blocking ERK signal mainly changed the phosphorylation of Op18/stathmin at Ser 25 site. The other study confirmed that ERK has a 20-fold preference for Ser25 as opposed to Ser38 of Op18/stathmin. 24 Besides these, PD98059 cooperatively decreased the levels of p-thr161-cdc2 and antiapoptosis factor Bcl-2 in NCI-H1299 cells treated by taxol, which implied that the regulation of taxol resistance was complex and might concern some other factors, such as cdc2 kinase activity and Bcl-2. The previous study indicated that cdc2 was also an upstream kinase of Op18/stathmin involving in cellular proliferation in nasopharyngeal carcinoma cells, whose activity was positive relevant to phosphor-thr161. 25,26 The other study also showed that inhibition of Bcl-2 expression sensitized lung cancer cells to taxol. 27
IL-10 is a pleiotropic cytokine; more and more studies showed that IL-10 autocrined by tumors is helpful for evading immunosurveillance and promoting cell proliferation, invasion, and survival, and relevant to malignant degree, high levels of autocrine IL-10 are a poor prognostic factor in patients with laryngeal carcinoma and melanoma and cervical cancer. 28 –30 The study uncovered that IL-10 autocrine was markedly inhibited for blockage of ERK signal in taxol-treated NCI-H1299 cells. The expressions of ERK and phosphor-ERK were significantly higher in taxol-resistant NCI-H1299 cells than in taxol-sensitive CNE1 cells; these confirmed again that ERK is associated with taxol resistance of tumor cells as Op18/stathmin did. 1 The study also showed that taxol itself induced cellular apoptosis slightly and decreased the expressions Op18/stathmin, ERK, and phosphor-ERK to a certain extent in NCI-H1299 cells.
These findings suggest that ERK-mediated Op18/stathmin signaling is involved in the taxol resistance development in NCI-H1299 cells; blocking ERK signal can dramatically promote the sensitivity of NCI-H1299 cells to taxol, minimizes the taxol dose applied clinically and alleviates the taxol toxicity, which provides a new indicator for treating drug-resistant carcinomas.
Footnotes
Acknowledgments
This work is supported by the National Natural Science Foundation of China (Grant No. 81272274) and, in part, by the Key Project of Hunan Province Scientific Research of Colleges and Universities (No. 12A018) and Natural Science Foundation of Hunan province (No. 12JJ3104). The authors thank Dr. Tao Yongguang from Cancer Research Institute of Central South University for valuable suggestions.
Disclosure Statement
No competing financial interests exist.