
Abstract
Cancer Biotherapy and Radiopharmaceuticals (CBR)
officially retracts the article entitled, “PHOX2B Is Associated with Neuroblastoma Cell Differentiation,” by Liqun Yang, Xiao-Xue Ke, Fan Xuan, Juan Tan, Jianbing Hou, Mei Wang, Hongjuan Cui, and Yundong Zhang (Cancer Biother Radiopharm 2016;31(2):44–51; doi: 10.1089/cbr.2015.1952).
Readers are advised that the CBR editorial office received a request via email from the first author, Dr. Liqun Yang, to replace Figure 3, indicating, “This article contains some errors in figure 3. In figure 3A (RA-7d) and figure 3C, we mixed up images from different groups when we prepared the figure. Unfortunately, these mistakes were not corrected when we submitted the manuscript.” This request came after the appearance of a comment posted to the PubPeer platform.1
The publisher of the journal made several attempts to contact the authors' institutions to inquire about the veracity of the study and the claimed error, and though a reply was eventually received, it did not appear to be official institutional documentation and was not signed by an institutional official's name but rather only by the institutional name, nor did it provide a comprehensive defensible explanation, rendering it unacceptable (see Fig. R1). The Publisher responded to all authors and to the sender of the “declaration” by stating that the document was not admissible and that the article would be officially retracted. No further response or rebuttal was received.
Insufficient declaration letter received by Editor and publisher.
Cancer Biotherapy & Radiopharmaceuticals
is committed to upholding the rigorous standards of scientific publishing and the veracity of the literature.
1. Actinopolyspora biskrensis.
Introduction
Neuroblastoma, a common type of extracranial solid tumor that occurs within the sympathetic nervous system, 1 is considered one of the most malignant pediatric cancers, accounting for up to 15% of all childhood cancer-related mortality. 2 Histologically, neuroblastoma tumors are quite heterogeneous: their phenotypes range from tumors composed of primarily undifferentiated or poorly differentiated neuroblasts to those composed of fully differentiated sympathetic neurons. 1,3,4 It has been reported that the degree of neuroblastoma differentiation is closely related to its prognosis. 5 Thus, patients with undifferentiated or poorly differentiated neuroblastomas have significantly poorer outcomes than those with well-differentiated neuroblastomas, who in contrast have relatively good survival. 6,7
Retinoic acid (RA) can induce neuroblastoma cell differentiation 8,9 and is considered an effective therapeutic agent for high-risk neuroblastomas in clinical practice. 10,11 Additionally, paired-like homeobox 2b (PHOX2B) has been demonstrated to be a critical mediator of neuroblastoma development in a TH-MYCN mouse model, 12 whereby the mice develop tumors that closely resemble the human disease. 13,14 In that previous study, the authors reported that PHOX2B is a major cellular regulator responsible for maintaining neuroblastoma stemness and promoting neuroblastoma progression in TH-MYCN mice. 12 The present study was performed with the following two aims: (1) to identify the functional role of PHOX2B in human neuroblastoma differentiation and (2) to confirm PHOX2B as a potential prognostic marker in neuroblastoma patients.
Materials and Methods
Patient data analysis
All patients' data were obtained from the R2 microarray analysis and visualization platform (
Cell culture
SMS-SAN and SMS-KCNR human neuroblastoma cell lines were grown in RPMI 1640 medium. The BE(2)-C cell line was grown in a 1:1 mixture of Dulbecco's modified Eagle's medium (DMEM) and Ham's nutrient mixture F12 (F12/DMEM). The SK-N-DZ, SK-N-AS, and SHEP1 cell lines were grown in DMEM. All growth media were supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin (P/S). The 293FT lentiviral packaging cell line was cultured in DMEM containing 10% FBS, 0.1 mM nonessential amino acids, 1 mM sodium pyruvate, 4 mM
Western blotting
Human neuroblastoma cells or xenograft tumors were harvested and washed once with ice-cold phosphate buffered saline (PBS). Cell pellets or tumor tissues were suspended in sodium dodecyl sulfate (SDS) sample buffer, boiled for 10 minutes, and centrifuged for 10 minutes. Each protein sample (60 μg) was separated by 12% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene fluoride membrane (Millipore Corporation), which was probed with primary and secondary antibodies and visualized using enhanced chemiluminescence (Beyotime). The following primary antibodies were used: rabbit antihuman PHOX2B (1:1000, ab183741; Abcam), mouse antihuman α-tubulin (1:2000, B-5–1-2; Sigma-Aldrich), mouse antihuman Mash1 (1:100, clone 24B72D11.1; BD Pharmingen), rabbit antihuman TH (1:500, ab6211; Abcam), goat antihuman GFAP (1:500, ab53554; Abcam), rabbit antihuman GATA3 (1:100, sc-9009,; Santa Cruz), rabbit anti-human peripherin (1:1000, ab53051; Abcam), and rabbit antihuman Snap25 (1:1000, ab53723; Abcam). The following horseradish peroxidase-conjugated secondary antibodies were used: goat antimouse, goat antirabbit, and mouse antigoat IgG (1:20,000; KPL).
Real-time quantitative polymerase chain reaction assay
Human neuroblastoma cells were harvested and lysed with TRIzol (Invitrogen; Life Technologies) to purify total RNA, which was reverse transcribed into cDNA using M-MLV reverse transcriptase (Promega). PHOX2B mRNA transcripts were quantified by real-time quantitative polymerase chain reaction (qPCR) using SYBR® Green PCR Master Mix (TaKaRa). Real-time qPCR assays were performed in triplicate using a StepOne Plus 7500 real-time PCR system (Bio-Rad). All individual values were normalized to that of glyceraldehyde-3-phosphate dehydrogenase, which was used as a control. The following primer sequences were used:
PHOX2B (Forward) 5′-AGTGGCTTCCAGTATAACCCG-3′
PHOX2B (Reverse) 5′-GGTCCGTGAAGAGTTTGTAAGG-3′
Cell growth and differentiation assays
For differentiation assays, RA (Sigma-Aldrich) was dissolved in dimethyl sulfoxide (DMSO), and a 10 mM stock solution was prepared. BE(2)-C cells were treated with 1 μM RA for 10 consecutive days. DMSO (0.1%) was used for the untreated control. Cell growth was determined by cell counting using the Trypan Blue dye analysis and a TC10™ Automated Cell Counter (Bio-Rad).
Cell proliferation assays
BE(2)-C cells were plated in 96-well culture plates. Cell proliferation was determined by the MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide; Sigma-Aldrich) assay. Briefly, 10 μL MTT was added to 100 μL medium in each well. The cells were incubated at 37°C for 2 hours, and 100 μL DMSO was added to each well to dissolve the MTT. The plate was then incubated for 20 minutes with shaking on an orbital shaker. Absorbance was measured at a wavelength of 560 nm using a microplate reader (Model 550; Bio-Rad).
Soft agar clonogenic assay and sphere formation assay
BE(2)-C cells were mixed with 0.3% Noble agar in DMEM supplemented with 10% FBS and plated at 1000 cells/well into six-well plates containing a solidified bottom layer (0.6% Noble agar in the same growth medium). After 21 days, colonies were stained with 5 mg/mL MTT and photographed. For serial soft agar assays, individual colonies were removed from the agar using sterile Pasteur pipettes and then pooled. After a brief treatment with trypsin, the colonies were mechanically dissociated into single-cell suspensions and then replated at 1000 cells/well (six-well plates). For sphere formation assays, cells were plated at 2000 cells/well in serum-free DMEM supplemented with 20 ng/mL EGF and bFGF (Invitrogen; Life Technologies) in Matrigel ultra-low attachment (ULA) plates. Spheres developing within 1–2 weeks were counted.
Lentiviral constructs and infection
The lentiviral constructs, pLK0.1-puro-GFPsi and pLK0.1-puro-PHOX2Bsi, were used in knockdown studies. First, the constructs were transfected into 293FT packaging cells using Lipofectamine® 2000 reagent (Invitrogen; Life Technologies); the virus-containing supernatant was harvested, and the titer was measured. The supernatant was then used to infect target cells in conjunction with 4 μg/mL Polybrene (Santa Cruz). Target cells were cultured in the presence of 2 μg/mL puromycin (Life Technologies) for 3 days after the final round of infection, and drug-resistant cells were pooled.
Xenograft assay
Six female NOD/SCID mice (4 weeks old) were used in a xenograft assay. The animals were maintained under specific pathogen-free conditions in the animal facility at Southwest University (Chongqing, China). Both flanks of each mouse were subcutaneously injected with 1 × 106 BE(2)-C cells suspended in 200 μL serum-free DMEM. One week after tumor cell injection, tumor growth was measured using calipers, and the tumor volume was calculated using the formula 4/3πr 3 , where “r” represents the tumor radius. The xenograft tumors were removed and weighed after 3 weeks of growth. The tumors were washed in ice-cold PBS and prepared for western blot assay. All animal experiments were conducted in accordance with the ethical standards of approved guidelines, and the protocol was preapproved by the Institutional Animal Care and Use Committee of Southwest University.
Quantification and statistical analyses
Each experimental value was confirmed by at least three independent experiments. Quantitative data were expressed as the mean ± SD. A two-tailed Student's t-test was performed for paired samples. p < 0.05 was considered statistically significant.
Results
High PHOX2B expression is associated with poor prognosis in neuroblastoma patients
The authors previously described the major regulatory role of PHOX2B in maintaining neuroblastoma stemness, which consequently promotes neuroblastoma tumor growth in mouse models.
12
Therefore, they investigated the relationship between PHOX2B expression and prognosis in neuroblastoma patients using microarray-based analysis with the Tumor Neuroblastoma public—Versteeg database, which is available online from the R2 microarray analysis and visualization platform (
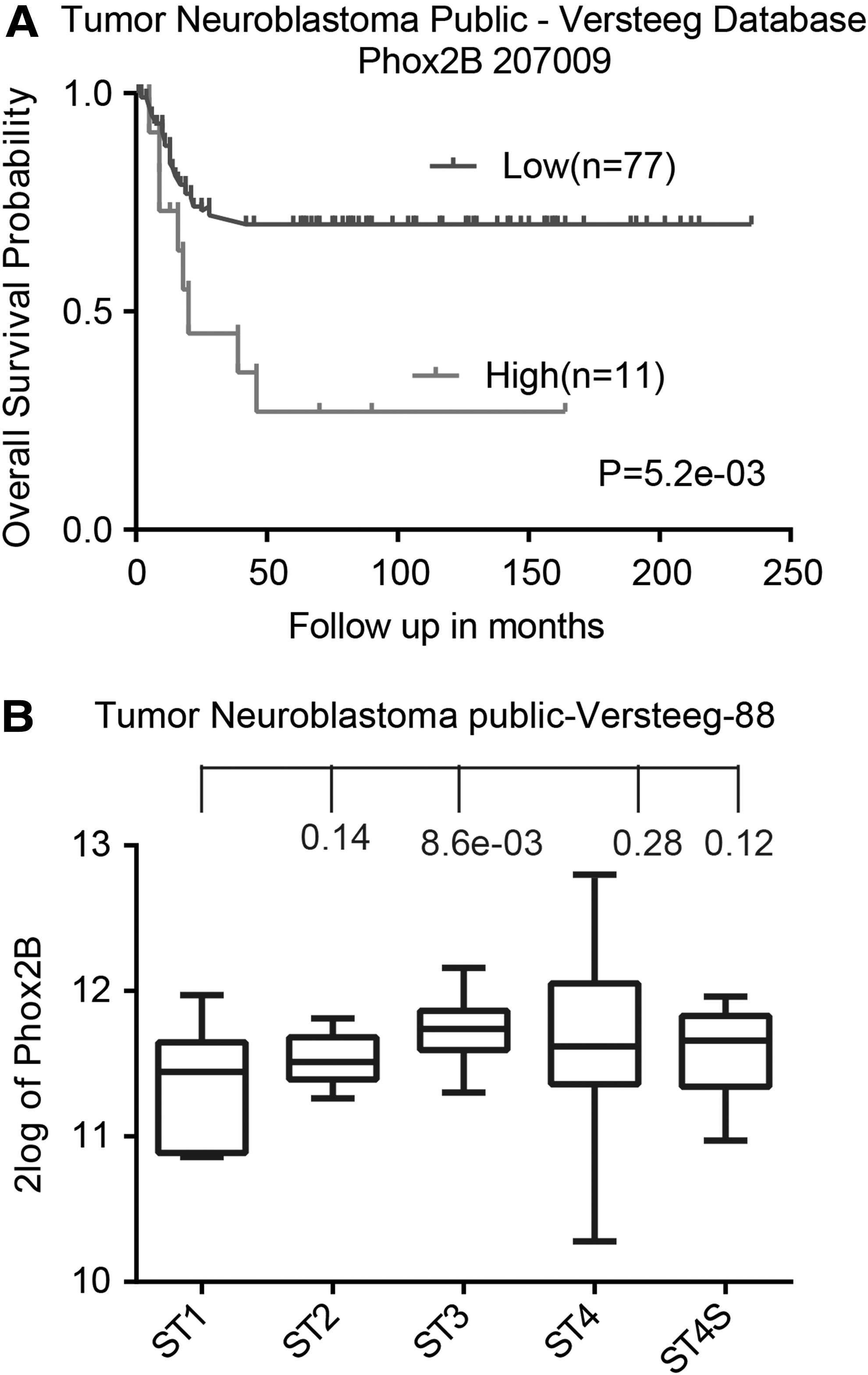
High levels of PHOX2B expression are associated with poor outcomes in neuroblastoma patients.
PHOX2B is commonly expressed in neuroblastoma cell lines
The authors next assessed PHOX2B expression in various human neuroblastoma cell lines. Western blot analysis revealed PHOX2B expression in the majority of tested neuroblastoma cells (Fig. 2A), including malignant SMS-SAN, SMS-KCNR, BE(2)-C, SK-N-DZ, and SK-N-AS cells. Conversely, PHOX2B expression was not detected in the SHEP1 cell line, which is derived from a benign and highly differentiated neuroblastoma. 16 –18 These trends were confirmed by the real-time quantitative polymerase chain reaction assay (Fig. 2B). Collectively, these results demonstrated that PHOX2B is associated with neuroblastoma differentiation and prognosis.
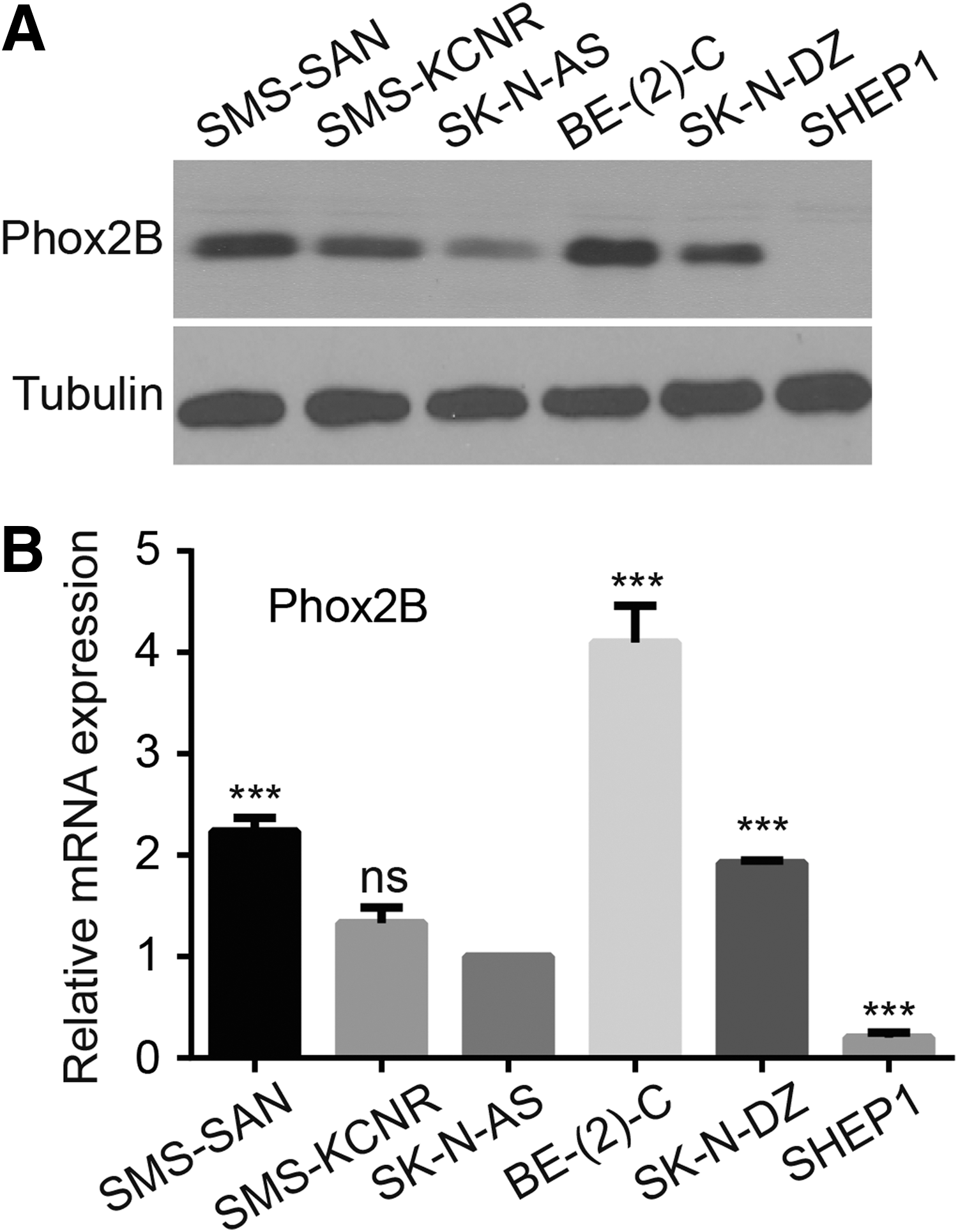
PHOX2B is commonly expressed in neuroblastoma cells.
PHOX2B is associated with neuroblastoma cell proliferation and differentiation
Since BE(2)-C cells exhibited the highest PHOX2B expression level, they treated these cells with RA, the drug most commonly used to induce neuronal differentiation in neuroblastoma. 8,10 As shown in Figure 3, after treating BE(2)-C cells with RA for a continuous period of time, the cells displayed morphologic features indicative of neuronal differentiation, including small and rounded cell bodies, scant cytoplasm, and extensive neurite-like processes (Fig. 3A). The results of cell counting and soft agar and sphere assays verified that RA treatment inhibited cell proliferation, self-renewal, and tumorigenicity in BE(2)-C cells (Fig. 3B, C). They also investigated the role of PHOX2B expression in RA-induced differentiation and found a significant time-dependent decrease in PHOX2B expression following RA treatment. These data suggested that RA-induced neuronal differentiation led to PHOX2B downregulation, which in turn inhibited neuroblastoma cell self-renewal ability and tumorigenicity.

PHOX2B expression is associated with neuronal differentiation in neuroblastoma cells.
PHOX2B knockdown inhibits neuroblastoma cell proliferation and self-renewal
To confirm an association between PHOX2B expression and self-renewal in neuroblastoma cells, they downregulated PHOX2B expression in BE(2)-C cells using small-interfering RNA (siRNA); siRNA against green fluorescent protein (GFPsi) was used as a control (Fig. 4A). MTT assay results verified that PHOX2B downregulation dramatically inhibited BE(2)-C cell proliferation (Fig. 4B). Next, they investigated the functional role of PHOX2B in maintaining BE(2)-C cell self-renewal ability. Following PHOX2B downregulation, BE(2)-C cells formed tiny, sparse colonies in soft agar and small spheres (Fig. 4C, D). These individual primary colonies were removed from the agar using sterile Pasteur pipettes, pooled and dissociated into single-cell suspensions, which were immediately replated at 1000 cells/well into six-well plates for serial soft agar assays. The results showed that the BE(2)-C cells that underwent PHOX2B knockdown could not form large colonies (Fig. 4E). These data demonstrate that downregulation of PHOX2B markedly inhibited neuroblastoma cell proliferation and self-renewal. Therefore, PHOX2B is critically associated with neuroblastoma growth.
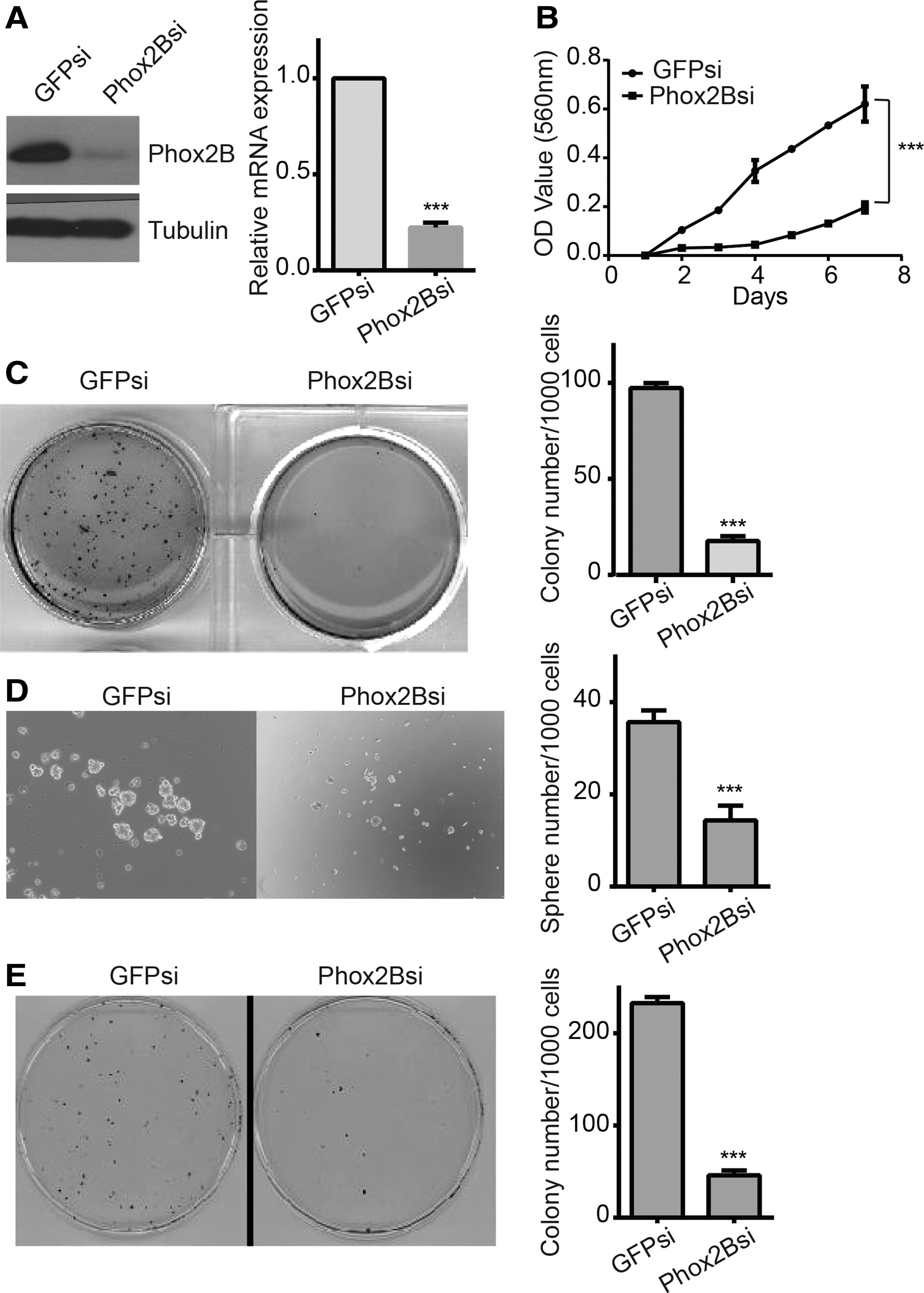
PHOX2B downregulation inhibited neuroblastoma cell proliferation and self-renewal ability.
PHOX2B knockdown inhibits neuroblastoma cell tumorigenicity and promotes differentiation
Xenograft tumor growth assays were performed to assess the role of PHOX2B in regulating neuroblastoma cell tumorigenicity. To accomplish this, PHOX2B-knockdown BE(2)-C cells were injected into NOD/SCID mice to form xenograft tumors; BE(2)-C-GFPsi cells were used as a control. As shown in Figure 5A, B, the volumes and weights of the xenograft tumors in the PHOX2Bsi group were much lower than those in the GFPsi group. These results indicate that PHOX2B downregulation inhibited neuroblastoma cell tumorigenicity. Following the removal of the xenograft tumors, immunoblot analysis was performed to investigate the differentiation status of the tumor cells. The results demonstrated that PHOX2B downregulation significantly decreased the expression of mammalian achaete-scute homolog 1 (MASH1), a stem cell/progenitor cell marker (Fig. 5C) 19 ; conversely, the expression of peripherin, a neuronal differentiation marker, was markedly upregulated. 20 PHOX2B knockdown also led to the downregulation of GATA binding protein 3 (GATA3), a prognostic marker of neuroblastoma. 21 They also detected the expression of other differentiation markers, including glial fibrillary acidic protein (GFAP) and synaptosomal-associated protein, 25 kDa (SNAP25); the former is an astrocyte marker, 22,23 and the latter is involved in the molecular regulation of neurotransmitter release. 24 The authors found that the expression of both proteins decreased following PHOX2B knockdown. Collectively, the above data indicated that PHOX2B knockdown inhibited neuroblastoma cell tumorigenicity and promoted neuronal differentiation in xenograft tumors.
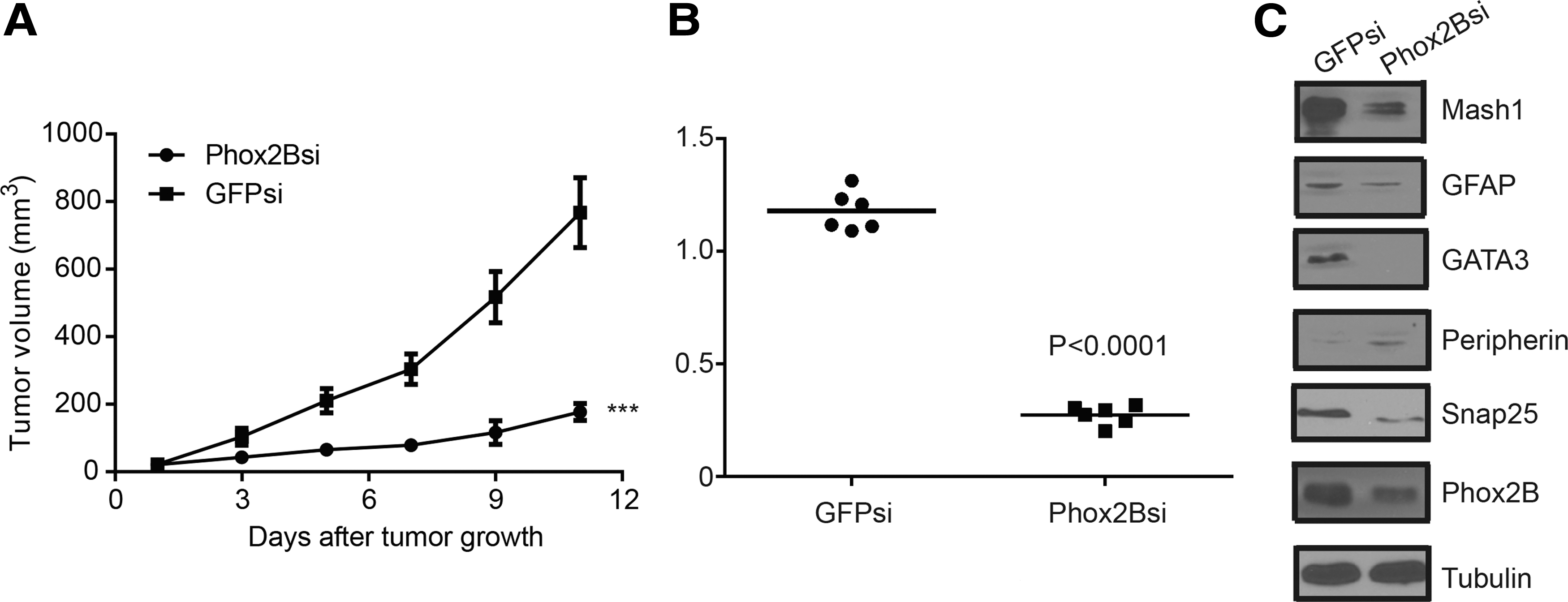
PHOX2B downregulation inhibited neuroblastoma cell tumorigenicity.
Discussion
Neuroblastoma is believed to arise from adrenal glands and sympathetic ganglia during early development of the sympathetic nervous system. 25 –27 During normal sympathetic neurogenesis, bone morphogenetic proteins (BMPs), a group of signaling molecules, are released from the dorsal aorta, which leads to the upregulation of MASH1 and PHOX2B. MASH1 and PHOX2B activation in turn upregulates the expression of heart and neural crest derivative-expressed protein 2 (HAND2), PHOX2A, GATA2, and GATA3, consequently promoting further neuronal differentiation. All of the above transcription factors collaborate through a complex regulatory network that ultimately induces the expression of tyrosine hydroxylase (TH) and dopamine β-hydroxylase, resulting in pan-neuronal and catecholaminergic differentiation into sympathetic ganglia. 28 –38 Neuroblastomas appear to form as a result of the deregulation of this network. Additionally, TH-MYCN mice, a transgenic model carrying human MYCN under control of the TH promoter, develop neuroblastomas that closely resemble human tumors. 13 In a previous study, they demonstrated that PHOX2B functions as a major cellular regulator in promoting neuroblastoma progression in TH-MYCN mice and that high PHOX2B expression is essential in maintaining neuroblastoma stemness. 12 In the present study, they investigated the role of PHOX2B in regulating neuroblastoma proliferation and differentiation in human cells. Analysis of a gene expression dataset of 88 neuroblastoma patients demonstrated that high PHOX2B expression is closely associated with poor prognosis. PHOX2B expression was also found to be related to clinical tumor stage in these neuroblastoma patients. Overall, these results indicate that PHOX2B is a prognostic marker in neuroblastoma.
Over the last few decades, a number of studies have illustrated that the degree of neuroblastoma cell differentiation is intimately related to prognosis in these patients. 1,4,5,39 As neuroblastomas undergo spontaneous, complete regression through neuronal differentiation in a proportion of patients, 40 malignant neuroblastoma is believed to be caused by the inhibition of cell differentiation. The first report that RA could induce neuroblastoma differentiation was published in 1982. 41 Since then, RA differentiation therapy has been considered an effective conventional therapeutic strategy with minimal side-effects on normal cells compared to chemo-radiotherapy, as normal nonmalignant cells are already fully differentiated. 8,42 In the present study, RA-induced neuronal differentiation resulted in PHOX2B downregulation, which led to the inhibition of neuroblastoma cell self-renewal ability. These data support the importance of PHOX2B as a mediator of neuroblastoma differentiation.
Additionally, PHOX2B knockdown was found to be associated with the suppression of neuroblastoma cell proliferation, self-renewal, and tumorigenicity. Furthermore, PHOX2B downregulation was followed by the upregulation of peripherin, a neuronal differentiation marker, as well as the decreased expression of MASH1, a stem cell/progenitor cell marker. According to the data presented in this study, PHOX2B has a functional role in regulating neuroblastoma cell differentiation and maintaining stemness. Thus, PHOX2B is a potential therapeutic target for neuroblastoma.
Footnotes
Acknowledgments
The authors' work was supported by the National Natural Science Foundation of China (Grant numbers 81201551 and 81502574) and the Fundamental Research Funds for the Central Universities (Grant numbers XDJK2013B020, XDJK2016C007, and 2362015XK09).
Disclosure Statement
No competing financial interests exist.