
Abstract
Uveal melanoma is the most common primary malignant intraocular tumor in adults and still lacks effective systemic therapies. Annexin A2 receptor (AXIIR), a receptor for Annexin II, was demonstrated to play an important role in multiple cells, but its role in uveal melanoma cells remains exclusive. Herein, the authors reported that overexpression of AXIIR was able to reduce cell viability and activate apoptosis apparently in the Mum2C uveal melanoma cell line. Meanwhile, overexpression of AXIIR could induce autophagy and increase autophagy flux. After autophagy was inhibited by chloroquine, enhanced apoptosis and cytotoxicity could be detected. In summary, these data highlighted the crucial role of AXIIR in reducing Mum2C cell viability through inducing apoptosis, while autophagy played a protective role in this process. Interference of this gene may be a promising method for uveal melanoma therapy and combination with specific inhibitor of autophagy may serve as a supplementary.
Introduction
Human uveal melanoma is the most common primary malignant intraocular tumor in adults. 1 According to the Collaborative Ocular Melanoma Study, the accuracy of clinical diagnosis of uveal melanoma is about 99%, while in most cases, cancer cells have already disseminated when diagnosed. Like other melanomas, uveal melanoma presents a high metastasis rate and poor prognosis. 2 –4 Options for the treatment of uveal melanoma include enucleation as well as newly developed conservative therapy. 5 Although modalities such as charged-particle radiotherapy, proton beam therapy, brachytherapy, photodynamic therapy, and surgical excision have been shown to be effective, 6 –9 however, to date, the selection of therapy has not been proven to affect the risk of distant disease spread or overall survival. 10 Recently, several promising molecular therapeutic targets such as protein kinase C and tyrosine kinase c-Kit have been identified and currently under clinical investigation. 11,12 Unfortunately, due to a high risk of local recurrence and drug resistance, the current management of uveal melanoma still lacks effective systemic therapies up to now. 13 –15 Therefore, it is imperative to seek more effective strategies for the treatment of uveal melanoma.
Annexin A2 receptor (AXIIR), also called chromosome 5 open reading frame 39 (C5orf39), is first identified as a receptor for Annexin A2 (ANXA2) in 2006. 16 The AXIIR gene encodes a protein with 193 amino acids, which can only be detected in primate species up to now. 4,16 As a receptor for Annexin II, AXIIR has been further implicated in regulating adhesion, migration, homing, and growth of prostate cancer cells and multiple myeloma cells. 17 –19 So far, the only known function about AXIIR is mediating ANXA2 signal. In several types of cells, AXIIR was located at the plasma membrane and functioned as a cell surface receptor for ANXA2. While recent work by Xiong et al. and Shiozawa et al. reported that AXIIR was located in the cytoplasm and nucleus. 20 However, previous report also indicates that AXIIR may not be the specific receptor of ANXA2. 21 It is shown that AXIIR is found to induce apoptosis in the chronic myeloid leukemia cell line, independent of ANXA2. 20,22 Although the exact role of AXIIR is poorly understood at present, it is noteworthy that AXIIR participates in comprehensive cell function in a more broad way. Therefore, a detailed understanding of the function of AXIIR will be helpful for further application.
Apoptosis and autophagy are two vital processes in cell to regulate cell growth, development, and homeostasis, and both of them could lead to programmed cell death (PCD). Apoptosis and autophagy are known to be type I and type II PCD separately and the relationship between them is complicated. 23 Treated with some stimulus they could occur together and when with other stimulus, they could appear independently. Meanwhile, autophagy can be protective or stimulative when apoptosis occurs.
In the present study, the authors investigated the role of AXIIR in apoptosis and autophagy in human uveal melanoma in vitro. They found that overexpression of AXIIR could be able to induce apoptosis through activation of caspase-3, capase-8, and caspase-9. Furthermore, AXIIR also acted as a novel activator of autophagy. Moreover, autophagy inhibition by chloroquine (CQ) significantly enhanced AXIIR-induced apoptosis. These data suggested that the combination of overexpression of AXIIR and autophagy inhibitors might be a new approach to treat human uveal melanoma.
Materials and Methods
Materials and reagents
Fetal bovine serum (FBS), RPMI 1640 medium, penicillin–streptomycin solution, Lipofectamine™ 2000, rabbit/mouse secondary antibody, and pcDNA3.1 vector were purchased from Invitrogen. Restriction enzyme, DNA ligase, DNA marker, and DNA polymerase were purchased from TaKaRa. Antibodies against AXIIR, caspases-9, p62 were obtained from Abcam. Antibodies against LC3, caspases-3, caspases-8, GAPDH were obtained from Cell Signaling Technologies. Cell counting kit-8 (CCK-8) was obtained from Dojindo Laboratories. Hoechst33258 dye was purchased from Sigma. The Annexin V-FITC Apoptosis Detection Kit was obtained from eBioscience. GFP-LC3 plasmid was purchased from Hanbio Company.
Cell culture
MUM2C cells were purchased from KeyGEN BioTECH, cultured in RPMI 1640 medium containing 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin, and maintained at 37°C in a 5% CO2 incubator. STR analysis was performed to authenticate the cell line used (Supplementary Table S1; Supplementary Data are available online at
Cell viability assay (CCK-8 assay)
Cell viability was evaluated by CCK-8 assay. Cells were seeded in a 96-well plate with the concentration of 3 × 103/mL and incubated overnight before the assay. After transfected with pcDNA3.1-AXIIR plasmid and incubated for 48 hours, the cells were treated with a fresh medium containing 10% volume of CCK-8 and then incubated at 37°C for 2 hours. The OD values at 450 nm were quantified by a microplate reader (BioTek synergy2). Assays were performed in three replicated wells, and each experiment was repeated three times independently.
Plasmids
The pcDNA3.1-AXIIR plasmid was constructed as follows: plasmid DNA containing the coding region of AXIIR inserted into HindIII and XbalI sites in the pcDNA3.1 vector. Mum2C cells were transfected with corresponding plasmids using Lipofectamine 2000 reagent following the manufacturer's protocol and the cellular expression was confirmed by Western blot analysis. The lentivirus of lenti-GFP-LC3 was packaged using a system consisting of VSVG, REV, and MDL plasmid.
Western blot analysis
After treatment, cells were washed with phosphate-buffered saline (PBS) twice and treated with the RIPA lysis buffer. An equal amount of protein was loaded into sodium dodecyl sulfate–polyacrylamide electrophoresis (10%) and transferred to polyvinylidene fluoride membranes. After blocking with 5% (w/v) skimmed milk for 2 hours at room temperature (25°C), the membrane was incubated with specific primary antibodies overnight at 4°C on the shaker. Then, membranes were incubated with indicated secondary antibodies, conjugated to horseradish peroxidase, and directed against mouse or rabbit IgG, for 2 hours at room temperature. Finally, membranes were visualized using the GeneGnome HR Image Capture System (Syngene).
Hoechst 33258 staining
Cells were grown on a 96-well cell culture dish overnight. After being subjected to corresponding treatments, the cells were fixed with 4% paraformaldehyde for 20 minutes, washed with PBS, and were subjected to 1 μg/mL Hoechst 33258 solution for 20 minutes in the dark. Then, the stained cells were observed under fluorescence microscopy (Olympus).
Cell apoptosis analyzed with flow cytometry
Cell apoptosis was detected with the Annexin V-FITC Apoptosis Detection Kit according to the manufacturer's instructions. In brief, harvested cells were incubated with Annexin V for 20 minutes in the dark. The cells were then washed with PBS and stained with propidium iodide. After that, the samples were detected using a MACSQuant Analyzer (Miltenyi Biotec).
GFP-LC3 punctate detection
Cells were seeded into cell culture plates overnight and then infected with lenti-GFP-LC3, which was used to make the autophagosomes visible in cells. After that, the cells were transfected with pcDNA3.1-AXIIR. After incubation for 48 hours, the cells were fixed with 4% paraformaldehyde for 20 minutes at room temperature and stained with Hoechst 33258 dye. The cells were measured by a laser scanning fluorescence confocal microscope.
Data analysis
All experiments were repeated three times independently and data were analyzed using GraphPad prism 6.01 software. Statistical analysis was performed with one-way analysis of variance and Student's t-test. A significance level of p < 0.05 was considered to be statistically significant.
Results
AXIIR reduced cell viability in uveal melanoma cells
The abnormal survival and growth of cancer cells are major characteristics of malignant tumors. Therefore, the authors investigated the effect of AXIIR on the survival of Mum2C cells after transfection of pcDNA3.1-AXIIR plasmid. After 48 hours of transfection, cell morphology was observed under an inverted light microscope, and cell viability was determined with CCK-8 assay. The results demonstrated that cells transfected with AXIIR plasmid experienced decreased cell growth ability and changed cell morphology compared with other two groups (Fig. 1A–C). Moreover, the data indicated a remarkable decrease of cell viability in the experimental group (Fig. 1D). Together, these results demonstrated that AXIIR could reduce cell viability significantly in human uveal melanoma cells.
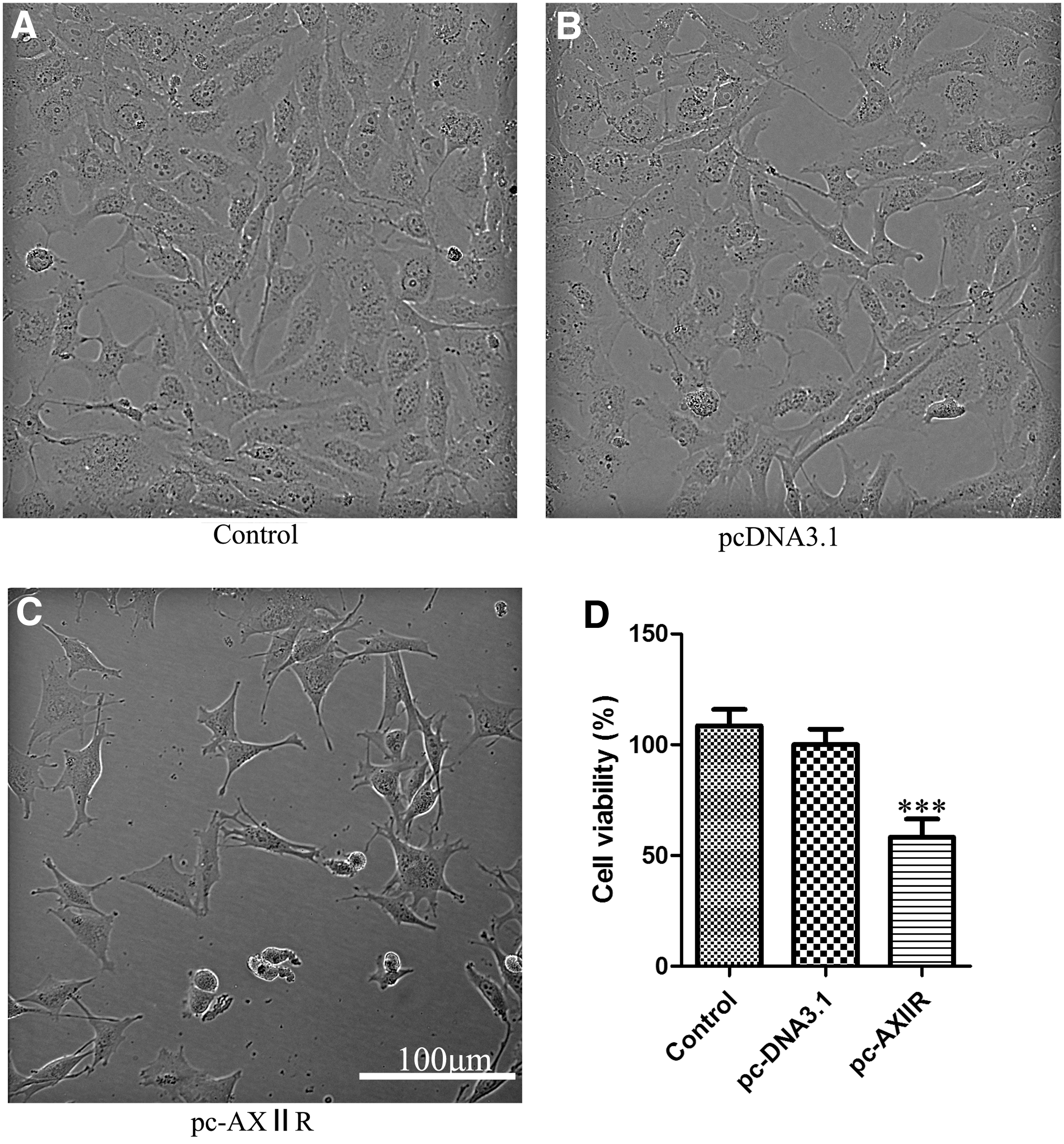
AXIIR reduced cell viability in Mum2C cells. Representative phase-contrast images of cells
AXIIR induced apoptosis and regulated components of apoptotic pathway
To better understand the possible mechanism of AXIIR-reduced cell viability, the authors examined the effect of AXIIR on cell apoptosis using Hoechst 33258 staining and flow cytometry analysis. After transfection for 48 hours, more nuclear condensation was observed in cells transfected with pcDNA3.1-AXIIR (Fig. 2A). In addition, 3.48% of the cells transfected with pcDNA3.1-AXIIR were Annexin V positive, whereas only 0.796% and 0.577% in vector group and blank control, respectively (Fig. 2B and Supplementary Figure S1; Supplementary Data are available online at
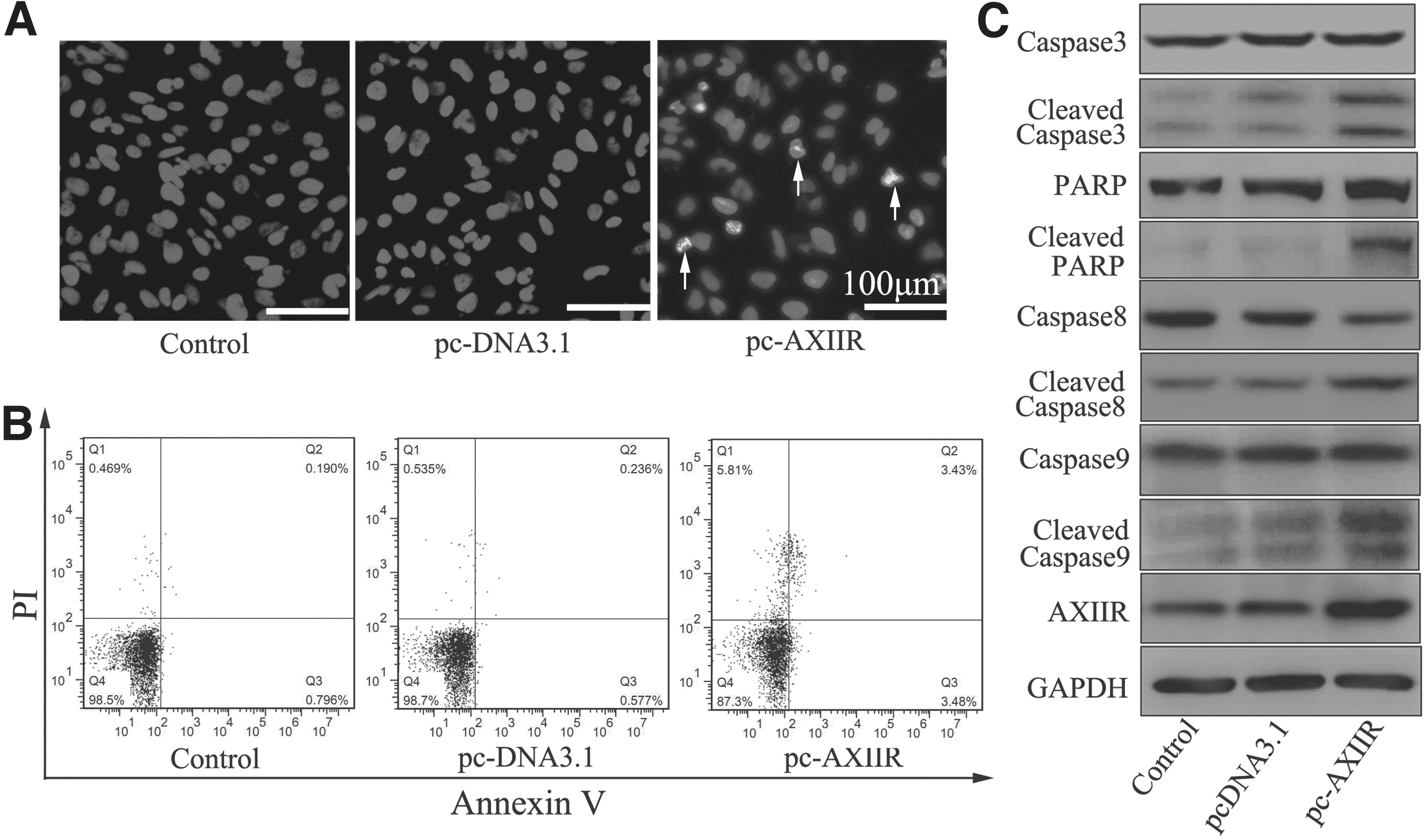
AXIIR induced apoptosis and regulated components of apoptotic pathway.
AXIIR activated autophagy
To further determine whether autophagy occurs during apoptotic process, multiple assays were evaluated. First, the authors detected the conversion of autophagy-associated protein LC3, the marker for the formation of autophagosomes. 24 As shown in Figure 3A, LC3-II was apparently increased in the experimental group. Moreover, confocal microscopy results demonstrated that cells overexpressing AXIIR exhibited a significant increase in the number of LC3 puncta, the appearance of autophagosomes (Fig. 3B). These findings suggested that autophagy might be involved in the apoptotic process of Mum2C cells overexpressing AXIIR.

AXIIR induced autophagy in Mum2C cells.
However, autophagy is a dynamic, multistep process, including the formation of phagophore, autophagosome, the amphisome, and the autolysosome. 25 –27 Thus, increases in the level of LC3-II immunoblotting are not appropriate measures of autophagic flux, the complete process of autophagy, including the delivery of cargo to lysosomes and its subsequent breakdown and recycling. 28 For this reason, the authors measured the autophagic flux with multiple assays. These results showed that treatment with CQ, a lysosomal inhibitor, led to a significant increase in endogenous LC3-II accumulation (Fig. 3C). Also, the expression of p62, whose decrease reflects the lysosomal degradation of autophagosomes, was remarkably decreased in the treatment group (Fig. 3C). Moreover, overexpression of AXIIR-induced downregulation of p62 could be reversed by CQ pretreatment, also suggesting an increase in autophagic flux. These data illustrated that AXIIR could activate autophagy in human uveal melanoma cells.
Autophagy inhibitor enhanced AXIIR-induced apoptosis
Although the data demonstrated that AXIIR could induce autophagy and apoptosis, the role of autophagy in the process of apoptosis was uncertain. Therefore, the authors further assessed whether autophagy played a role in AXIIR-induced apoptosis. In this study, they used CQ to explore the role of autophagy in AXIIR-induced apoptosis. They first examined the cytotoxic effect of CQ and DMSO, and the data showed that both CQ and DMSO had no evident effect on cell viability (Fig. 4A). While AXIIR-induced cytotoxicity was enhanced when the cells were pretreated with CQ (Fig. 4B), which suggested a protective role of autophagy. Then, they detected apoptosis-related proteins, which demonstrated that caspase-3 and PARP were activated with CQ pretreatment (Fig. 4D; Fig. S3). Collectively, these data strongly suggest that autophagy seemed to play a protective role against AXIIR-induced apoptosis in human uveal melanoma cells.
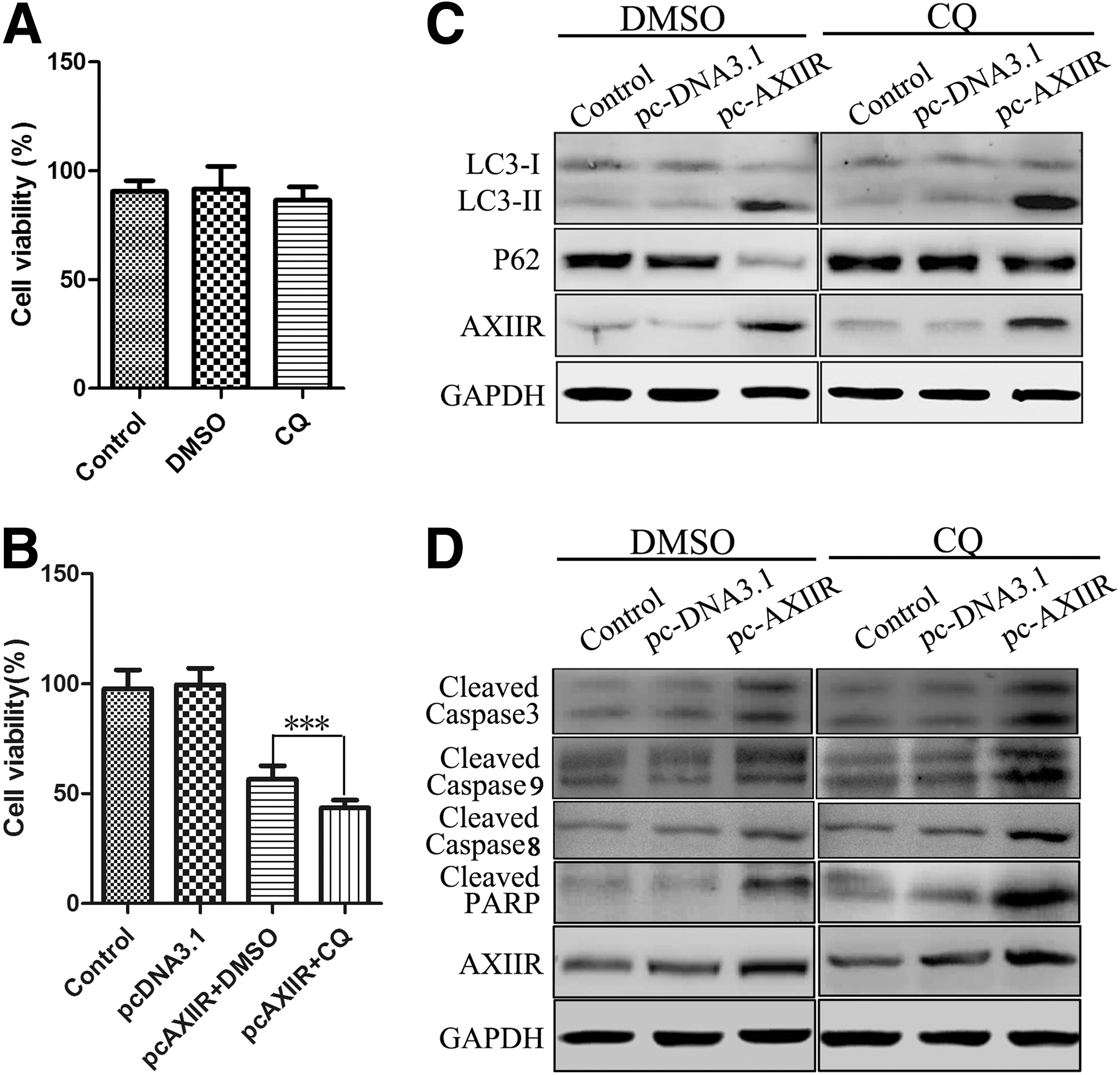
Autophagy inhibitor enhanced AXIIR-induced apoptosis.
Discussion
Being the most common intraocular tumor, human uveal melanoma has captured researchers' eyes broadly, but the treatment of uveal melanoma is far from satisfactory. Therefore, novel strategies need to be developed, and the molecular therapy may be a potential one. In this study, the authors reported that overexpression of AXIIR was able to reduce human uveal melanoma cell viability and the mechanism might be overstimulated apoptosis accompanied with autophagy.
Apoptosis is the best-known type of PCD that characterized by activation of caspases. 29,30 There are two major apoptotic pathways: the intrinsic pathway is triggered by the release of cytochrome C and AIF, 31 –36 leading to the activation of caspase-9; while the extrinsic pathway is induced by the activation of caspase-8. 37 –39 In this study, the authors found a remarkable increase of caspases-8 and caspases-9 cleavage, suggesting that both intrinsic and extrinsic apoptotic pathways participate in AXIIR-induced apoptosis. The results were partially consistent with the previous study that AXIIR activated apoptosis in the chronic myeloid leukemia cell line, and AXIIR activated caspase-8 in a manner different from conventional apoptotic pathways, independent from FADD (Fas-associated with death domain) and AXA2. 20
Furthermore, the authors reported a novel function of AXIIR for the first time that it could induce autophagy in Mum2C cells. Autophagy is an evolutionarily conserved process, in which long-lived proteins, misfolded proteins, and damaged organelles are recycled to maintain cellular homeostasis. 40 The autophagic effect of AXIIR is demonstrated by its ability to induce LC3 conversion and the formation of GFP-LC3 puncta. Moreover, protein levels of p62, a well-known autophagic substrate, were also decreased. Meanwhile, the authors used CQ, a lysosomotropic drug that raises intralysosomal pH and impairs autophagic protein degradation, to further ascertain the result. 41 As was shown, LC3 conversion was enhanced, and AXIIR-induced reduction of p62 was reversed when the cells were pretreated with CQ, which proved the elevated autophagic flux. Collectively, these data illustrate the ability of AXIIR to increase autophagic flux in Mum2C cells.
The relationship between apoptosis and autophagy is intricate and paradoxical. Sometimes, they cooperate with each other to cause cell death, during which autophagy may act as an upstream signal of apoptosis. Whereas autophagy could also antagonize apoptotic cell death in some cases. 42 To further explore the role of autophagy in AXIIR-induced apoptosis, the authors examined cell proliferation and the activation of apoptosis-relevant molecules with CQ pretreatment. Interestingly, they found CQ could effectively augment the inhibitory effect of cell proliferation, as well as increase the levels of cleaved caspase-3 and cleaved PARP. Along with these data, it suggested a protective role of autophagy in AXIIR-induced apoptosis of Mum2C cells. Meanwhile, previous report shows that AXIIR might lead to the damage of mitochondria and release of cytochrome. 20 Mitochondria play an essential role in the mechanism of apoptosis. 43 Initiators lead to increased permeabilization of the outer mitochondrial membrane, which causes the leakage of small molecules, such as cytochrome c and second mitochondrion-derived activator of caspases (Smac), from the mitochondria into the cytoplasm, followed by recruitment of apoptotic protease activating factor-1 (Apaf-1) and procaspase 9 to form the apoptosome, and execution of apoptosis. 44 –51 Integrating the updated knowledge with these findings, in this study, the authors hypothesized that overexpression of AXIIR in Mum2C cells led to the damage of mitochondria, which initiates the mitochondrion-mediated apoptosis signaling pathway, as well as autophagy activation that occurs as a protective compensation reaction.
In conclusion, this article demonstrated that AXIIR was able to induce apoptosis and autophagy in Mum2C cells, and autophagy played a protective role of in AXIIR-induced apoptosis, which provide evidence of AXIIR as a potent target in anticancer therapy. Since the poor prognosis of uveal melanoma is often associated with deficiency in apoptotic machinery, AXIIR could provide a novel approach for uveal melanoma treatment. It is noteworthy that the autophagy pathway may serve as a supplementary. Combination with specific inhibitor of autophagy may augment the therapeutic efficacy of human uveal melanoma treatment.
Footnotes
Acknowledgments
The authors are grateful to Meihong Chen (State Key Laboratory of Medical Molecular Biology, Institute of Basic Medical Sciences, Chinese Academy of Medical Sciences and Peking Union Medical College) for excellent technical assistance. This work was supported by the National Natural Science Foundation of China (No. 81271017, No. 81470652, and No. 81221061) and the National Science and Technology Major Project (2011ZXJ09104-10C).
Disclosure Statement
No conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.