
Abstract
MicroRNAs are small RNA that are tightly interrelated with the initiation, development, and metastasis of cancers. Studies have shown that miR-10b is increased in various cancers. However, the underlying mechanisms of miR-10b in the occurrence and metastasis of endometrial cancer are poorly understood. To investigate its roles and correlations with Homeobox box 3 (HOXB3) in endometrial cancer, cancer tissues and adjacent normal endometrium tissues from 20 patients with endometrial cancer were studied. miR-10b expression was significantly up-regulated (p < 0.01) in endometrial cancer tissue, whereas HOXB3 was lowly expressed. The silence of miR-10b resulted in significantly enhanced cell apoptosis, and remarkably reduced cell proliferation, migration, and invasion (p < 0.05). Moreover, the protein levels of HOXB3 were increased in KLE cells with silenced miR-10b, and dual-luciferase reporter assay suggested that miR-10b could directly target HOXB3. Furthermore, overexpression of HOXB3 promoted cell apoptosis but inhibited cell proliferation, migration, and invasion (p < 0.01). To conclude, miR-10b might control cell apoptosis, proliferation, migration, and invasion in endometrial cancer via regulation of HOXB3 expression.
Introduction
E
Homeobox (HOX) genes encode a wide range of transcription factors that modulate cell proliferation and participate in cancer pathogenesis. Reportedly, the introduction of an antisense of HOXB7 or HOXB13 in ovarian cancer cells can significantly suppress cell invasion. 4 Especially, HOXA10 and p21 are down-regulated in endometrial cancer tissue, and HOXA10 can inhibit cell cycle and proliferation of endometrial cancer cells via regulation of the expression of p21. 5 As a member of the HOX gene family, HOXB3 has been confirmed to be expressed in breast cancer, 6 lung cancer, 7 ovarian cancer, 8 as well as squamous cervical carcinomas. 9 Generally, HOXB3 is expressed in early mammalian embryogenesis, and its overexpression in hematopoietic cells results in progressive myeloproliferation. It promotes angiogenesis, 10 which is crucial for the development of cancer. Hoxb3 can induce the transformation and proliferation of rat prostate cancer cells via cooperating with Pbx1, a transcriptional activator that is associated with HOX proteins. 11 Therefore, although little is known about the role of HOXB3 in endometrial cancer, it is assumed that HOXB3 may play a role in endometrial cancer.
With a length of ∼22 nucleotides, microRNAs (miRNAs) are small RNA molecules that can negatively and post-transcriptionally control the expression of their target genes. Reportedly, miRNAs can regulate cell proliferation, apoptosis, migration, and invasion, and, thus, take part in the initiation, development, and metastasis of cancers. For instance, miR-10b has oncogenic effects in glioblastoma cells, as it is up-regulated in human glioblastoma tissues and glioblastoma cells in comparison with normal tissues and astrocytes, and the inhibition of miR-10b can strongly repress the proliferation, invasion, and migration of glioblastoma cells. 12 Moreover, miR-10b is a metastasis driver in various cancers, for example, gastric cancer, 13 minimally invasive follicular thyroid carcinoma, 14 and ovarian cancer. 15 miR-10b level is remarkably elevated in gastric tumor with lymph node metastasis (LNM) in comparison with that in gastric tumor without LNM. Also, miR-10b is highly expressed in cells with high metastatic potential, and it promotes cell invasion by targeting HOXD10 in gastric cancer. 13 However, few reports have demonstrated the roles of miR-10b in endometrial cancer.
Reportedly, miR-10b is located in the cluster of HOX genes, and HOX genes are the downstream target genes of miR-10b, for example, HOXD10. 13,15 Only one study has clearly reported that miR-10 represses HoxB3a, however, in the spinal cord of zebrafish. 16 Based on sequence alignment, it was predicted that miR-10b might target human HOXB3. Therefore, miR-10b may participate in endometrial cancer via targeting HOXB3. To confirm this prediction, miR-10b was silenced and HOXB3 was overexpressed in endometrial cancer cells, respectively, and then, cell proliferation, apoptosis, migration, and invasion were detected. Besides, dual-luciferase reporter assay was performed to validate the regulatory relationship between miR-10b and HOXB3.
Materials and Methods
Patients and cell lines
Twenty patients with endometrial cancer were collected from the First Affiliated Hospital of Guangxi Medical University between January 2014 and January 2015, and their cancer tissues and adjacent normal endometrium tissues were obtained. After a full explanation of this study, all patients provided written informed consents. This study was approved by the institutional review board of the First Affiliated Hospital of Guangxi Medical University. Based on WHO classifications, all the endometrial cancer tissues were evaluated by two pathologists. Patients who were diagnosed with other malignant diseases and pregnant or lactating individuals were excluded from our experimental group. No patients underwent preoperative chemotherapy and/or radiotherapy. Human endometrial cancer cell lines were purchased from ATCC, including RL95-2, AN3CA, ECC-1, and KLE. Cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Cat. No. SH30022.01B; Hyclone) supplemented with penicillin (100 U/mL; Gibco), streptomycin (100 μg/mL; Gibco), and fetal bovine serum (FBS, 10% [v/v], Cat. No. 10099-141; Gibco). Cells were incubated at 37°C in a 5% CO2 humidified atmosphere.
qRT-PCR
Total RNA was extracted from cells using Trizol agent (Takara Biotechnology) according to the manufacturer's instructions. cDNA was synthesized using Bestar qPCR RT Kit (DBI Bioscience) in a 20 μL reaction system. Primer sequences were synthetized by Sangon Biotech (Table 1). Then, RT-PCR was performed using the Bestar® SybrGreen qPCR Master Mix kit (DBI Bioscience) and Strata gene Mx3000P real-time PCR instrument (Agilent Technologies) in a 20 μL reaction system. PCR conditions were as follows: 94°C 2 minutes, 40 cycles (94°C 20 seconds, 58°C 20 seconds, and 72°C 20 seconds), and 72°C 10 minutes. Samples were run in triplicate. Relative mRNA levels of miR-10b and HOXB3 were normalized by using the 2−ΔΔCt method 17 based on the expression of U6 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH), respectively.
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; HOXB3, homeobox B3; miR-10b, microRNA-10b.
Western blot and quantification
Total proteins were extracted from all samples and then quantified using the Pierce™ BCA Protein Assay kit (Cat. No. 23227; Thermo Scientific). Rabbit anti-human anti-HOXB3 (1:1000 dilution), rabbit anti-human anti-GAPDH (1:1000 dilution; G9545), and horseradish peroxidase-conjugated goat anti-rabbit IgG (1:20,000 dilution; Cat. No. BA1054) were purchased from Abcam, Sigma-Aldrich, and Boster Biotech, respectively. Standard Western blot was carried out, and band intensity was quantified using BioPix iQ22. Image-Pro Plus 6.0 software.
Construction and transfection of miR-10b-inhibitor or pcDNA3.0-HOXB3
The open reading frames of HOXB3 were cloned using the primer pairs HOXB3 NotI-F and HOXB3 xhol-R. The primer pairs for HOXB3 were 5′-ATTTGCGGCCGCCTTTCCAAGTTGCCACCCAC-3′ (forward) and 5′-CCGCTCGAGTCTTCCTCCCCATCCCCTAA-3′ (reverse). The pcDNA3.0-HOXB3 was obtained by the cloning of HOXB3 genes into the mammalian expression vector pcDNA3.0 (Invitrogen) to express HOXB3, and the pcDNA3.0 empty vector was used as a control.
All transfections were performed using Lipofectamine™ 2000 (Cat. No. 52887; Invitrogen) according to the manufacturer's instructions. Cells stably silencing miR-10b were generated via transfection of miR-10b-inhibitor, whereas cells stably overexpressing HOXB3 were obtained via transfection of the plasmid carrying HOXB3 (pcDNA3.0-HOXB3). Scrambled sequences of miR-10b inhibitor and pcDNA3.0 were, respectively, used as nontargeting controls (NC).
Dual-luciferase reporter assay
The information about the luciferase experiment and the construction of 3′-UTR Luciferase Vector were introduced in the revised manuscript. A fragment of the HOXB3 mRNA-3′-UTR was amplified by PCR and purified using Qiaquick gel extraction kit (Qiagen). The purified PCR product was cloned into multiple cloning sites of psiCHECK-2 Luciferase Vector (Promega) downstream of the firefly luciferase gene to construct the 3′UTR Luciferase Vector of HOXB3.
The primers for wild-type 3′-UTR-HOXB3 were as follows: HOXB3-XhoI F, 5′-CCGCTCGAGTGTTAGGACTGAAGGTTTGC-3′, HOXB3-NotI R, and 5′-ATAAGAATGCGGCCGCAGAACACACGCTTTTTTGAG-3′. The mutant 3′-UTR-HOXB3: HOXB3-mut F, 5′-TTAACGTGAAACTGATCGATATTTGAACAAACTG TCCCAGGC-3′, HOXB3-mut R, and TGTTCAAATATCGATCAGTTTCACGTTAAG TCAAGAGAGCAAC.
According to the manufacturer's instructions, luciferase reporter assay was performed using the Dual-Luciferase® Reporter (DLR™) Assay System (Cat. No. E1910; Promega). The luciferase activity was detected 48 hours after transfection using a Turner 20/20 luminometer (Turner Biosystems).
Cell apoptosis assay based on flow cytometry
Cells were treated with 0.25% tyrisin 48 hours after transfection, and they were then stained with Annexin V and PI (Annexin V/PI apoptosis kit; Cat. No. AP101; Multi Sciences Biotech) for 15 minutes. An FACSCalibur flow cytometer (BD Biosciences) was used to detect cell apoptosis.
Cell proliferation assay based on CCK8 method
Cell proliferation was assessed 24, 48, and 72 hours after transfection using the CCK8 kit. The optical density at 450 nm (OD450) was detected using an iMark 680 microplate reader (Bio-Rad Laboratories).
Migration and invasion assays based on transwell
Cell migration and invasion were detected using Costar transwell chamber (Cat. No. 3422; Corning Life Sciences), and the membrane with 8 μm pores was coated with Matrigel (20 μL 2.5 mg/mL; BD Falcon) in invasion assay. The upper and lower chambers were filled with 1% FBS/DMEM and 10% FBS/DMEM, respectively, and 105 cells were seeded in the upper chamber. After incubation at 37°C for 24 hours, the cells that had migrated into the lower chamber were fixed in 4% paraformaldehyde/HBS-Ca2+ and then stained with 0.5% crystal violet. Images were obtained using an inverted microscope (MOTIC AE-31).
Statistical analyses
Two-tailed unpaired Student's t-test was performed using SPSS 19.0 (IBM Corporation), and p-value <0.05 was set as the criterion for statistical significance. In figures, p-value <0.05, p-value <0.01, and p-value <0.001 were represented by one, two, and three asterisks, respectively. All data were presented as mean ± SD.
Results
Levels of miR-10b and HOXB3 in endometrial cancer
The clinicopathologic characteristics of 20 patients were shown (Table 2). Levels of miR-10b and HOXB3 were measured in endometrial cancer and adjacent normal tissues. Results showed that the miR-10b level in cancer tissue was significantly up-regulated in more than 3.6-fold of normal tissue (p-value <0.01, Fig. 1A). On the contrary, the protein level of HOXB3 was down-regulated in cancer tissue in comparison with adjacent normal tissue (Fig. 1B). KLE cell line showed a high miR-10b level (Fig. 1C) and a low HOXB3 level (Fig. 1D), and it was used to conduct the following functional experiments.
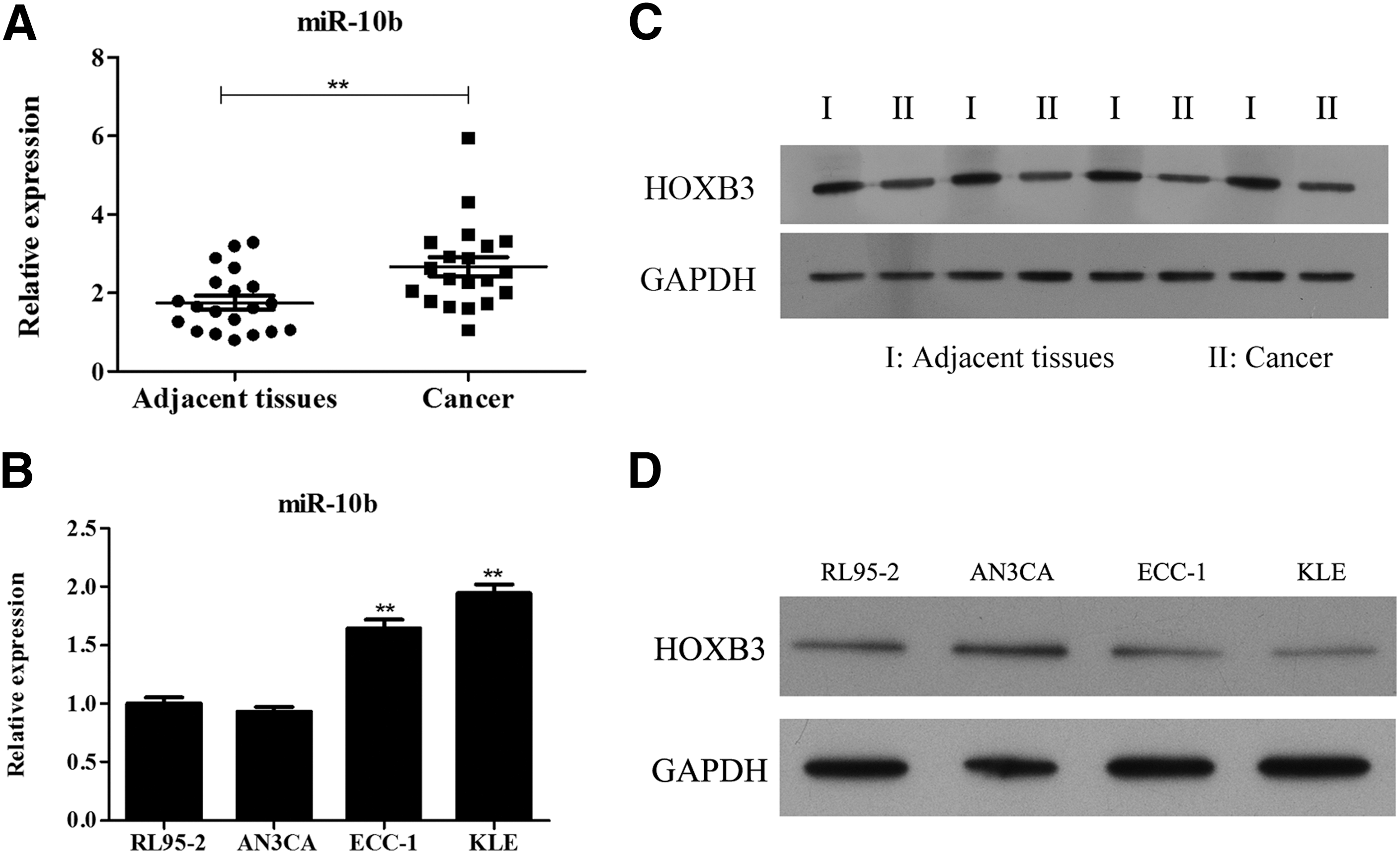
Levels of miR-10b and HOXB3 in the tissue and cell lines of endometrial cancer.
miR-10b inhibits apoptosis and promotes proliferation, migration, and invasion of KLE cells
After the transfection of miR-10b-inhibitor, miR-10b level was significantly decreased to 87.5% of NCKLE cells (p-value <0.01, Fig. 2A). Cell proliferation in the miR-10b-inhibitor group was remarkably lower than that in the NC group during the 72 hours after transfection, and especially, it was reduced to 68.8% of the NC group at 72 hours after transfection (p-value <0.01, Fig. 2B). Cell apoptosis was significantly enhanced (p-value <0.01, Fig. 2C). Besides, the number of KLE cells that migrated through matrigel-coated and noncoated transwell filter was decreased, namely, invasion (p-value <0.05, Fig. 2D) and migration (p-value <0.01, Fig. 2E) were significantly reduced with the silencing of miR-10b. Similarly, overexpression of miR-10b mimics could promote the cell proliferation, migration, and invasion (Fig. 2F–H).

Proliferation, apoptosis, invasion, and migration of KLE cells after transfection with miR-10b-inhibitor.
miR-10b targets HOXB3 in KLE cells
After miR-10b silencing, the protein level of HOXB3 was increased in KLE cells (Fig. 3A). In dual-luciferase reporter assay, luciferase activity in the miR-10b mimics group declined to 47.6% of the NC group (p-value <0.01, Fig. 3B), indicating that miR-10b targeted HOXB3.
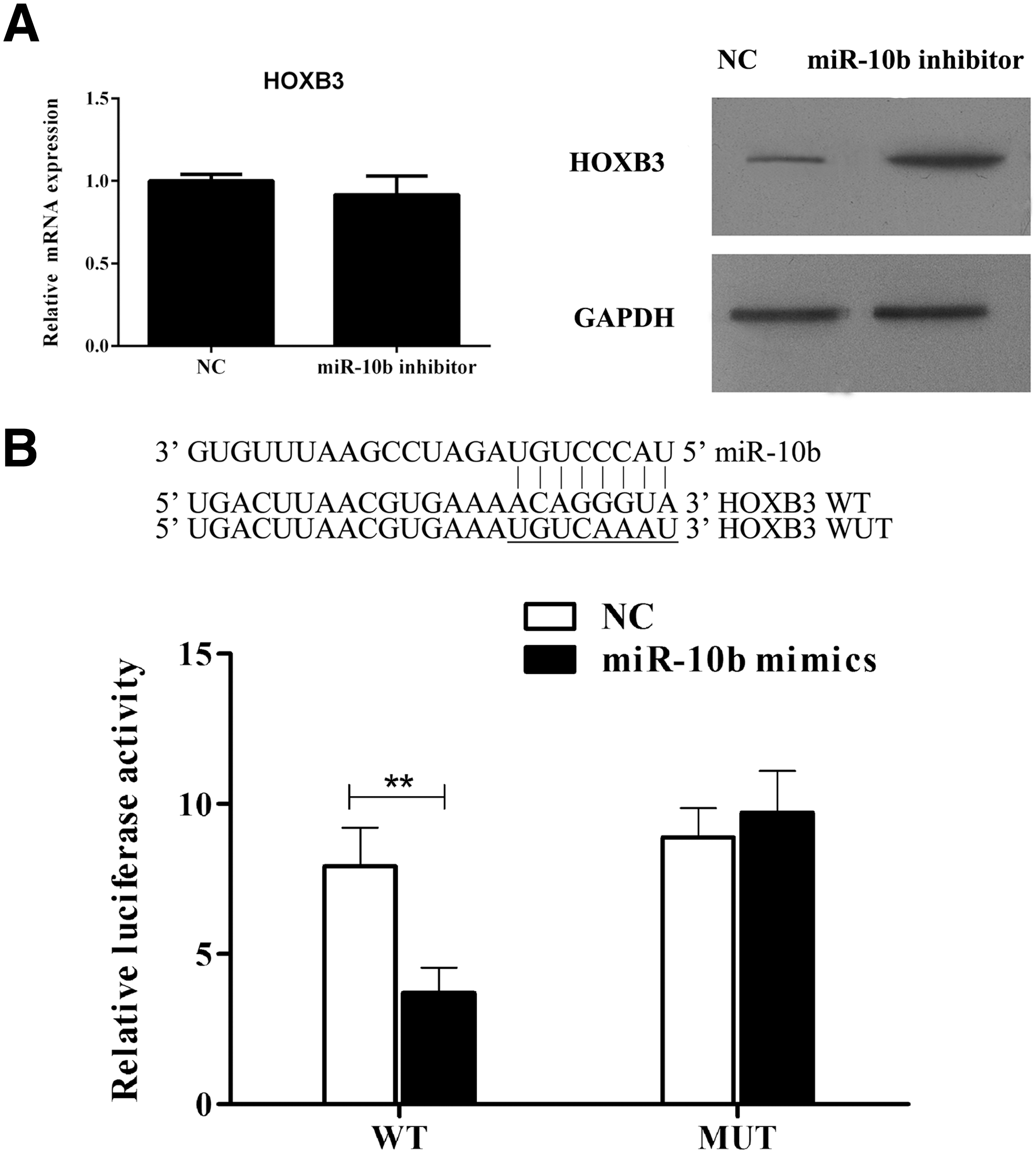
miR-10b targets HOXB3 in KLE cells.
HOXB3 promotes apoptosis and inhibits proliferation, migration, and invasion of KLE cells
After the transfection of pcDNA3.0-HOXB3, HOXB3 level was increased in KLE cells (Fig. 4A). Cell proliferation in the HOXB3 overexpression group was constantly lower than that in the NC group, and especially, it was reduced to 72.3% of the NC group at 72 hours post-transfection (p-value <0.01, Fig. 4B). Cell apoptosis was remarkably promoted (p-value <0.01, Fig. 4C), whereas both invasion (p-value <0.01, Fig. 4D) and migration (p-value <0.01, Fig. 4E) were reduced in KLE cells that overexpressed HOXB3.
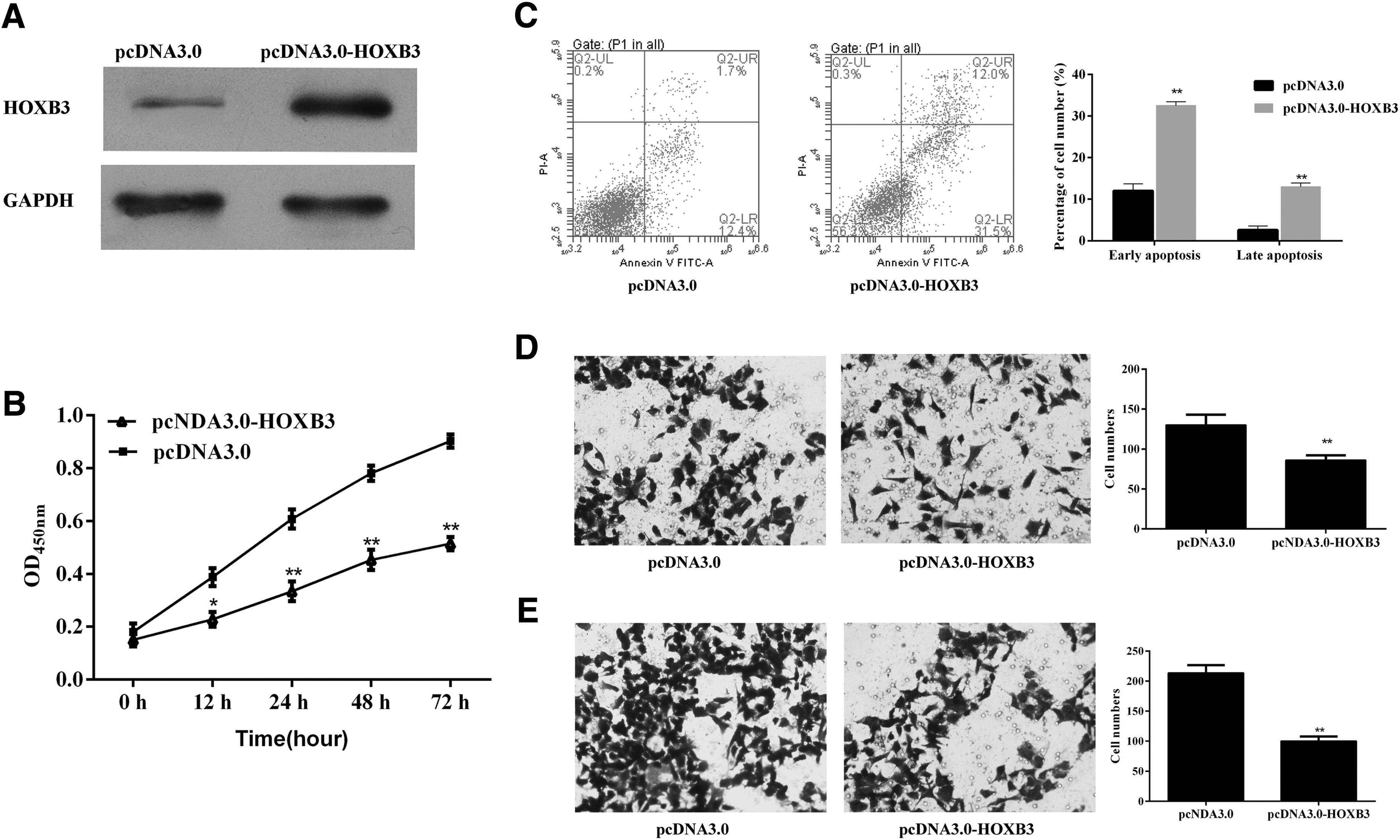
Proliferation, apoptosis, invasion, and migration of KLE cells after transfection with pcDNA3.0-HOXB3.
Discussion
With an increasing mortality, endometrial cancer is the most common gynecologic malignancy. 1 A clearer understanding of its molecular mechanism might help us treat patients with endometrial cancer. 3
Studies have identified crucial roles of HOX genes in endometrial cancer. HOXA10 can block cell cycle and suppress cell proliferation in endometrial cancer. 5 Also, HOXA10 is down-regulated in endometrial carcinomas, and HOXA10 overexpression inhibits invasion in vitro and tumor metastasis in nude mice. 18 HOXA10 induces expression of E-cadherin, an epithelial cell adhesion molecule, promoting epithelial–mesenchymal transition (EMT). 18 HOXB13 is abnormally overexpressed in endometrial cancer tissues, and it is positively related with the invasive ability of endometrial cancer cells, as its knockdown leads to a 90% reduction in the invasive ability of AN3CA cells. 19 However, the role of HOXB3 in endometrial cancer is still unclear. In the present study, HOXB3 was found to be down-regulated in endometrial cancer tissues. Besides, after overexpressing HOXB3 in KLE cells, cell apoptosis was increased, whereas cell proliferation, migration, and invasion were decreased. This was consistent with the down-regulation of HOXB3 in endometrial cancer tissue. These results indicated that HOXB3 might promote apoptosis and inhibit proliferation, migration, and invasion of endometrial cancer cells, and it might be a novel therapeutic target for treating endometrial cancer.
Accumulating evidences have shown that miR-10b regulates cell proliferation, migration, and invasion, and, thus, plays crucial roles in various cancers, for example, glioblastoma, 12 minimally invasive follicular thyroid carcinoma, 14 gastric cancer, 13 and ovarian cancer. 15
Although the down-regulation of miR-10b* has been identified in endometrial serous adenocarcinoma, 20 the roles of miR-10b in endometrial cancer have rarely been studied. In the present study, miR-10b was found to be significantly up-regulated in endometrial cancer tissues, which was consistent with previous studies about glioblastoma 12 and gastric cancer. 13 Moreover, the silencing of miR-10b accelerated apoptosis and suppressed proliferation, migration, and invasion of endometrial cancer cells. This was consistent with the up-regulation of miR-10b in endometrial cancer tissue. Therefore, miR-10b might be an oncogene that promotes the initiation, development, and metastasis of endometrial cancer via the enhancement of cell proliferation, migration, and invasion.
Studies have demonstrated that miR-10b targets HOX genes in gastric cancer 13 and ovarian cancer. 15 It was predicted that miR-10b might target human HOXB3. However, clearer evidence is required to demonstrate the regulation between miR-10b and HOXB3 in human endometrial cancer. In the present study, HOXB3 was up-regulated in endometrial cancer cells with silenced miR-10b. What is more, dual-luciferase reporter assay indicated that miR-10b targeted HOXB3. Thus, miR-10b might modulate cell apoptosis, proliferation, migration, and invasion in endometrial cancer via targeting HOXB3.
Nowadays, miRNAs are regarded as a novel class of potential therapeutic targets, since their biosynthesis, maturation, and regulatory activity can be manipulated using oligonucleotides. 21 Moreover, the less toxic oligonucleotides and an effective delivery system may minimize the side effects that are caused by low specificity for cancer cells or cross-sensitivity with endogenous miRNAs. 21,22 A number of studies on miRNA-based therapy have been conducted in animal models. 23 Therefore, miRNA-based therapy is a novel promising method in treating cancer, and our results might provide a potential therapeutic target, miR-10b, for treating primary endometrial cancer and blocking tumor metastasis.
However, limitations exist in this study. HOXB3 is a transcription factor, and its target genes participate in not only apoptosis, proliferation, migration, and invasion but also EMT. 24 The regulatory pathway of the miR-10b-HOXB3-target will be further investigated in our future study.
Footnotes
Acknowledgment
This study was supported by Nature Science Foundation of Guangxi (No. 2013GXNSFAA01956).
Disclosure Statement
There are no existing financial conflicts.